携带抗白粉病基因Pm21的小麦–簇毛麦小片段易位染色体在不同小麦背景中的传递率及遗传稳定性
2016-03-03王海燕袁春霞于春艳孙昊杰陈佩度王秀娥
王海燕 肖 进 袁春霞 徐 涛 于春艳 孙昊杰 陈佩度 王秀娥
南京农业大学作物遗传与种质创新国家重点实验室 / 细胞遗传研究所 / 江苏省现代作物生产协同创新中心, 江苏南京 210095
携带抗白粉病基因Pm21的小麦–簇毛麦小片段易位染色体在不同小麦背景中的传递率及遗传稳定性
王海燕肖进袁春霞徐涛于春艳孙昊杰陈佩度王秀娥*
南京农业大学作物遗传与种质创新国家重点实验室 / 细胞遗传研究所 / 江苏省现代作物生产协同创新中心, 江苏南京 210095
摘要:抗白粉病基因Pm21来自小麦近缘种簇毛麦。小麦–簇毛麦小片段顶端易位系NAU418 (T1AS.1AL-6VS)和小片段中间插入易位系NAU419 (T4BS.4BL-6VS-4BL)携带Pm21, 高抗白粉病, 是小麦抗病育种新种质。为了对其育种利用提供依据, 以NAU418和NAU419为亲本分别与来源于不同生态区的郑麦9023等12个小麦品种杂交, 杂种F1再分别与来源于不同生态区的农艺亲本进行正、反回交, 研究两种易位染色体在不同小麦背景中的遗传稳定性及其通过雌雄配子的传递规律。DNA分子原位杂交结果表明, 在杂种F1花粉母细胞减数分裂中期I (Pollen Mother Cell, PMC MI), 两种易位染色体分别可以与对应的小麦染色体配对形成棒状二价体。正、反交结果分析表明, NAU418中的小片段顶端易位染色体T1AS.1AL-6VS通过雌配子和雄配子的传递率分别为8.00%~50.98%和7.89%~45.07%, NAU419中的小片段中间插入易位染色体T4BS.4BL-6VS-4BL通过雌配子和雄配子的传递率分别为29.17%~52.38% 和7.69%~47.06%。表明2个易位系中的易位染色体都可以通过雌、雄配子传递, 但是其通过雄配子的传递率均显著低于通过雌配子的传递率。
关键词:小麦; 簇毛麦; 白粉病; 小片段易位; 配子传递率
本研究由国家高技术研究发展计划(863计划)项目(2011AA100101-03, 2011AA10010201), 国家自然科学基金项目(31201204), 中央高校基本科研业务费专项资金(KYZ201403)和江苏省优势学科建设工程项目(PAPD)资助。
This study was supported by the National High Technology Research Program (863 Program) of China (2011AA100101, 2011AA10010201), the National Natural Science Foundation of China (31201204), the Fundamental Research Funds for the Central Universities (KYZ201403) and the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
第一作者联系方式: E-mail: hywang@njau.edu.cn, Tel: 025-84395344
小麦白粉病是由小麦白粉菌(Blumeria graminearum f. sp. tritici)引起的全世界范围的主要病害之一,可导致小麦产量和品质大幅下降, 严重时产量损失达50%以上。由于小麦白粉病菌具有高度的变异性,新毒性生理小种的产生导致原有抗病品种丧失抗性,引起病害再度流行, 如来自黑麦的抗白粉病基因Pm8曾在世界范围广泛利用, 但目前在大部分地区均丧失了抗性[1]。因此, 发掘新的更为丰富的抗病基因资源, 培育和利用新的抗病品种, 对于保障小麦安全生产有重要意义。
二倍体簇毛麦(Haynaldia villosa, VV)是原产于地中海沿岸的小麦近缘物种, 具有抗病、抗逆、高产、营养高效和优质等特性, 尤其是对白粉病表现免疫[2]。为了将簇毛麦所携带的抗白粉病基因导入普通小麦, 南京农业大学细胞遗传研究所通过染色体工程的手段获得高抗白粉病的小麦–簇毛麦T6VS/6AL整臂易位系, 该易位系携带来源于簇毛麦的抗白粉病基因被命名为Pm21, 它是目前国际上公认的抗性最强、抗谱最广的抗白粉病基因[3-6]。在我国, 小麦–簇毛麦6VS/6AL易位系已经得到广泛应用, 育种单位以其为亲本, 已经选育和审定了20多个品种, 还有多个正在参加区试, 推广面积超过330万公顷[7]。但是, T6VS/6AL易位系在获得抗性的同时也伴随株高略有增加的不利性状[7]。
转移外源基因较理想的方法是选育携有外源有用基因的染色体易位, 尤其是小片段染色体易位。导入的染色体片段愈短, 它携带的不利基因越少,遗传越稳定, 在育种中的应用价值也就越大。因此,小片段易位系, 尤其是小片段中间插入易位系的创制一直受到小麦遗传育种学家的高度重视。南京农业大学细胞遗传研究所为了更好地利用抗白粉病基因Pm21, 利用电离辐射技术选育获得2个抗白粉病的小片段易位系, 即携带T1AS.1AL-6VS染色体的小片段顶端易位系NAU418和携带T4BS.4BL-6VS-4BL染色体的小片段中间插入易位系NAU419[6]。
导入小麦背景中的外源染色体或染色体片段在雌、雄配子中的传递, 受外源染色体来源、与之发生易位的小麦染色体、外源染色体片段大小和插入方式, 小麦品种遗传背景等的影响。研究外源染色体或携带外源染色体片段的易位染色体在小麦背景中的遗传和传递规律, 可以为这些外源种质的育种利用提供依据[7-12]。为了更好地利用6VS染色体短臂上所携带的抗白粉病基因Pm21, 本研究选用易位系NAU418和NAU419分别与不同生态区的主栽小麦品种组配杂交组合, 通过对杂种F1及BC1F1进行细胞遗传学分析和分子标记鉴定, 了解易位染色体在不同小麦遗传背景中的遗传稳定性以及通过雌、雄配子的传递率, 为易位系在小麦育种和生产中的利用提供参考依据。
1 材料与方法
1.1植物材料
小麦–簇毛麦小片段易位系NAU418和NAU419由南京农业大学细胞遗传所提供。其中, NAU418是簇毛麦6V染色体短臂的抗白粉病小片段纯合顶端易位系, 6VS染色体的1/8片段易位到1A染色体的长臂上, 易位染色体是T1AS.1AL-6VS; NAU419是簇毛麦6V染色体短臂的抗白粉病中间插入易位系, 6VS染色体的1/7片段插入到4B染色体的长臂上,易位染色体是T4BS.4BL-6VS-4BL。选取长江中、下游冬麦区的扬麦15、扬麦11、宁麦13、南农0686;黄淮麦区的郑麦9023、周麦9823、济麦22、远丰175、烟农19; 北方冬麦区的石4185和西南冬麦区的绵阳26和川麦54等12个小麦品种, 分别与NAU418 和NAU419杂交和回交。以上材料均由南京农业大学细胞遗传研究所征集保存。其中远丰175和川麦54的亲本中虽然有小麦–簇毛麦T6VS.6AL易位系,但本研究中所用的通过白粉病抗性鉴定对白粉病表现感病, 不含T6VS.6AL整臂易位染色体。
1.2易位染色体的雌、雄配子传递分析
易位系NAU418和NAU419在开花前人工去雄, 2~3 d后分别采集上述12个小麦主推品种的花粉进行授粉, 获得杂种F1。为研究易位染色体通过雌、雄配子的传递率, 将杂种F1分别作父、母本与相应
小麦品种回交, BC1F1种子播种于塑料大棚, 分单株取苗期幼叶, 用SDS法提取基因组DNA。正、反BC1F1回交后代中, 含有外源易位染色体的植株占总植株数的百分数即为易位染色体的传递率。
1.3细胞遗传学分析
选取处于减数分裂中期I的花药, 用固定液(无水乙醇∶冰醋酸体积比为3∶1)固定24 h以上, 45%醋酸压片, 相差显微镜(Olympus BX60)观察并照相。
参照Zhang等[13]描述的方法, 以Fluorescein-12-UTP标记的簇毛麦基因组DNA为探针进行荧光原位杂交, 用PI (propidium idodide)套染, 通过SPOT CCD (charge coupled device)获取FISH图像, 在450~490 nm波长下, 簇毛麦染色体呈绿色, 小麦染色体呈红色。
1.4分子标记鉴定
采用SDS法[14]提取基因组DNA。共显性标记引物CINAU15 (南京农业大学细胞遗传所开发[15]和提供)用于追踪抗白粉病基因Pm21, 具902 bp扩增产物的单株被认为含有Pm21基因, 引物由上海捷倍思生物公司合成。PCR体系10 µL, 含1× buffer (10 mmol L–1Tris-HCl, pH 8.3, 50 mmol L–1KCl)、1 mmol L–1MgCl2、200 µmol L–1dNTPs、4 ng EST-PCR引物和10 ng模板DNA。扩增程序为 94℃初始变性3 min; PCR扩增35个循环, 每循环在94℃变性50 s, 55℃退火1 min, 72℃延伸1 min; 在72℃延伸10 min。采用8%聚丙烯酰胺凝胶电泳及银染技术检测PCR扩增产物。
1.5自交结实率计算
成熟期统计各发育小穗基部两朵小花的总数和结实数, 计算所有的F1代和12个小麦品种的自交结实率。自交结实率 = (结实小花数/小花总数)×100%。
1.6白粉病抗性鉴定
将所有杂种F1以及BC1F1播种于南京农业大学江浦试验农场塑料大棚, 在参试材料两侧种植高感白粉病小麦品种苏麦3号, 作为白粉菌诱发行, 以含有Pm21基因的小麦品种南农9918和感病的小麦品种苏麦3号分别作为抗、感病对照。分别于拔节期和抽穗期调查每个单株的白粉病抗性。抗病性分为高抗(茎、叶和穗上均无可见病斑或白粉菌孢子)和感病(茎、叶和穗上有病斑或白粉菌孢子)两类。
2 结果与分析
2.1NAU418和NAU419杂种F1代PMC MI的染色体构型及易位染色体的传递率
在NAU418和NAU419与各小麦品种的杂种F1中, 易位染色体常分别与1A和4B染色体配对形成棒状二价体(图1), 其染色体构型平均为0.12 I+20.94 II和0.17 I+20.92 II, F1自交结实率平均为95.71%和92.48% (表1)。说明2个易位系与不同小麦品种杂交结实均表现正常, 这与F1代在细胞学上的稳定性相吻合。
2.2NAU418和NAU419中易位染色体通过雌、雄配子的传递率
利用可追踪Pm21的特异引物CINAU15F和CINAU15R对各正、反回交群体单株的DNA进行扩增, 清晰可见902 bp的特异产物(图2), 并根据扩增结果确定易体染色体是否传递到这些单株中。NAU418中T1AS.1AL-6VS在11个小麦品种背景条件下通过雌配子的传递率为8.00%~50.98%, 在周麦9823 (38.10%)、石4185 (38.10%)、扬麦15 (43.30%)、远丰175 (47.95%)和烟农19 (50.98%)中的传递率与50%的理论值无显著差异, 而在其余品种中通过雌配子的传递率偏离理论值(表2)。T1AS.1AL-6VS在这11个小麦背景中通过雄配子的传递率为8.00%~ 45.07%, 在郑麦9023 (41.18%)、绵阳26 (39.74%)、石4185 (33.33%)、扬麦15 (38.46%)和远丰175 (45.07%)中的传递率与理论值无显著差异, 而在其他品种中的传递率显著低于理论值(表2)。表明NAU418中易位染色体T1AS.1AL-6VS通过雌、雄配子的传递率受遗传背景的影响。
NAU419中T4BS.4BL-6VS-4BL在9个不同小麦背景下通过雌配子的传递率为29.17%~52.38%, 经χ2检验, 均与理论值(50%)无显著差异; 在7个不同小麦背景下通过雄配子的传递率为7.69%~47.06%,仅在石4185中的传递率与理论值无显著差异(表2)。说明不同遗传背景下NAU419中易位染色体T4BS. 4BL-6VS-4BL通过雌配子的传递率显著高于通过雄配子的传递率。

图1 (NAU418 ×周麦9823) F1 (A)和(NAU419 ×周麦9823)F1 (B) 的PMC MI FISH结果Fig. 1 (NAU418 × Zhoumai 9823) F1(A) and (NAU419 × Zhoumai 9823) F1(B) PMC MI FISH箭头指易位染色体与对应的小麦染色体配成棒状二价体。Arrows show that the translocation chromosomes paired and formed rod bivalents with their corresponding wheat chromosomes.
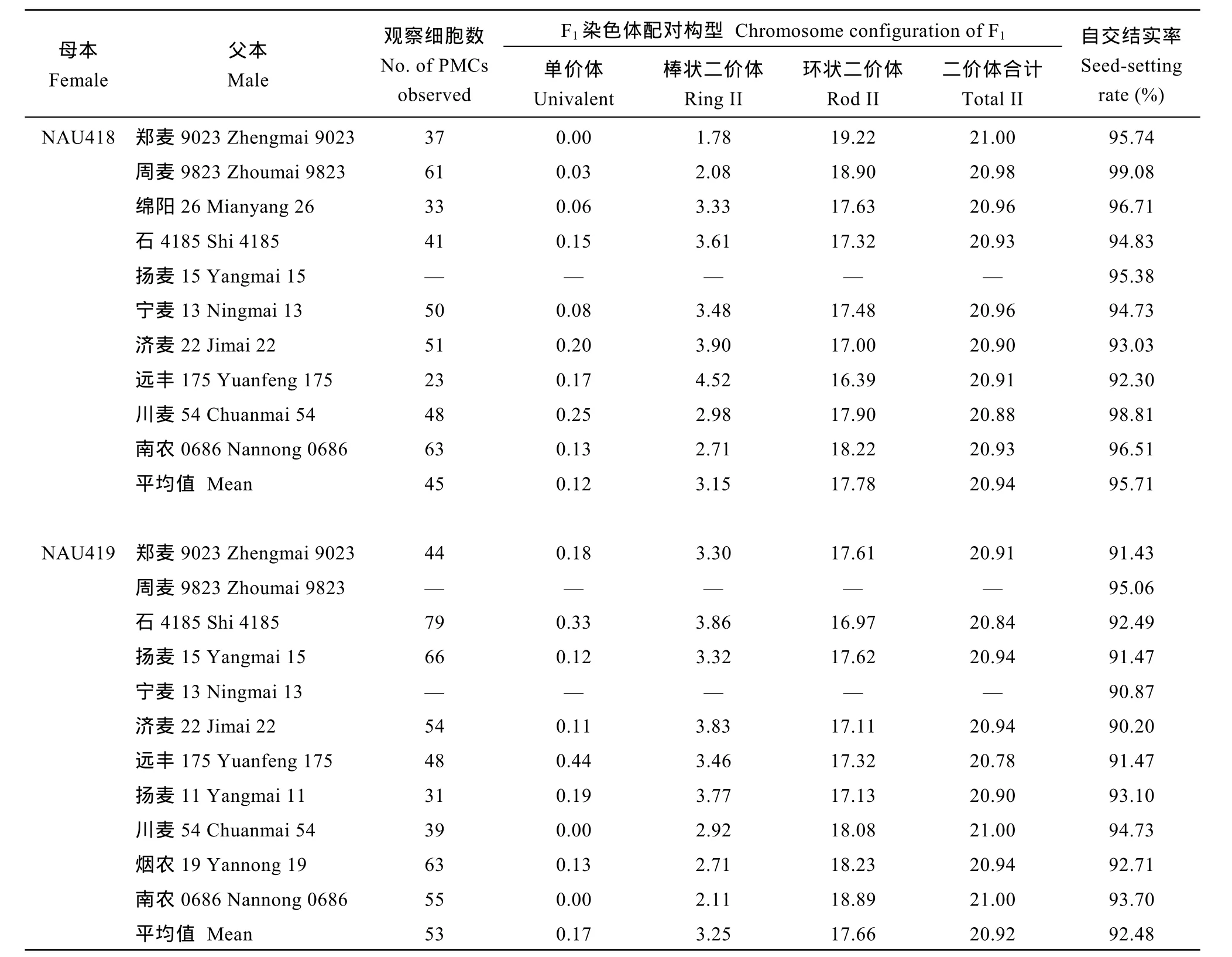
表1 NAU418和NAU419与不同品种的杂种F1 PMC MI染色体构型及自交结实率Table 1 Chromosome configuration and seed-setting rates of F1hybrids from crosses of NAU418 and NAU419 with different wheat varieties
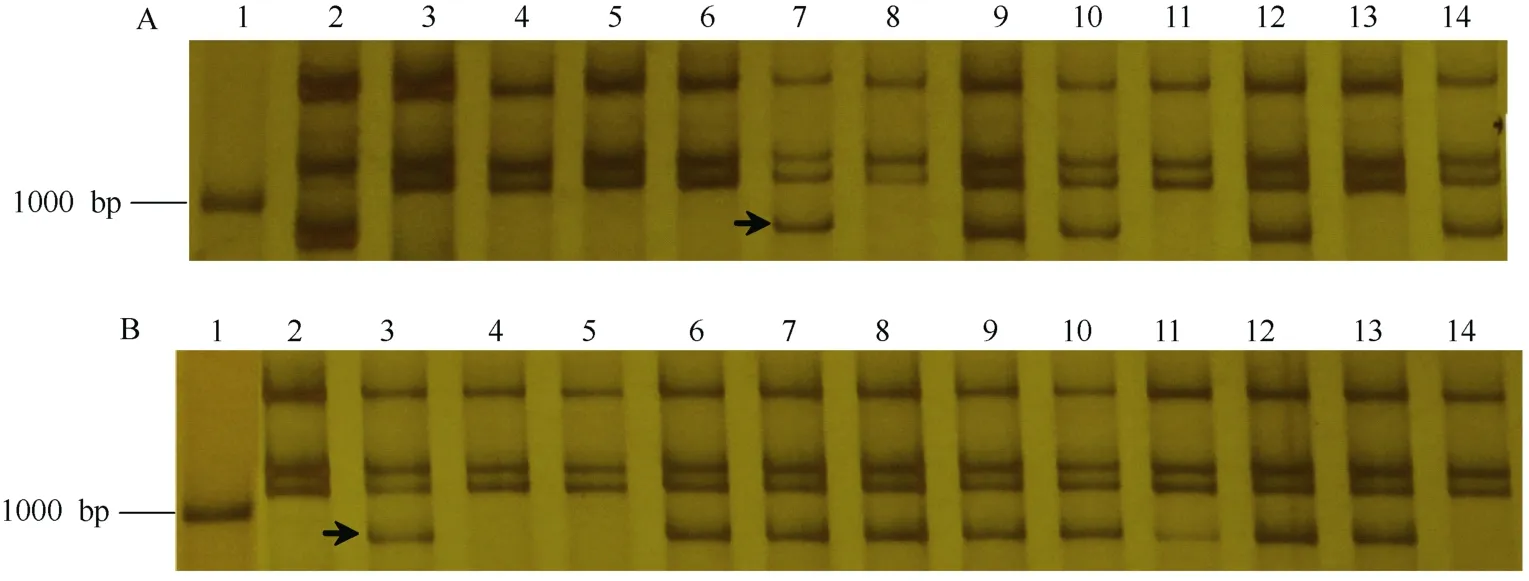
图2 与Pm21基因紧密连锁的EST标记CINAU15在NAU418和NAU419回交群体部分单株的扩增结果Fig. 2 Electrophoresis of Pm21-specific EST marker CINAU15 in partial plants from the NAU418 and NAU419 backcross populationsA: (NAU418 × 郑麦9023) × 郑麦9023群体。1: Marker; 2: 南农9918 (T6VS.6AL); 3: 中国春; 4~14: 回交群体部分单株。B: (NAU419 × 郑麦9023) × 郑麦9023群体。1: Marker; 2: 中国春; 3~14: 回交群体部分单株。箭头指示902 bp的特异条带。A: (NAU418 × Zhengmai 9023) × Zhengmai 9023 population. 1: Marker; 2: Nannong 9918 (T6VS.6AL); 3: Chinese Spring; 4–14: partial individuals from the backcross population. B: (NAU419 × Zhengmai 9023) × Zhengmai 9023 population. 1: Marker; 2: Chinese Spring; 3–14: partial individuals from the backcross population. Arrows show the 902-bp specific band.
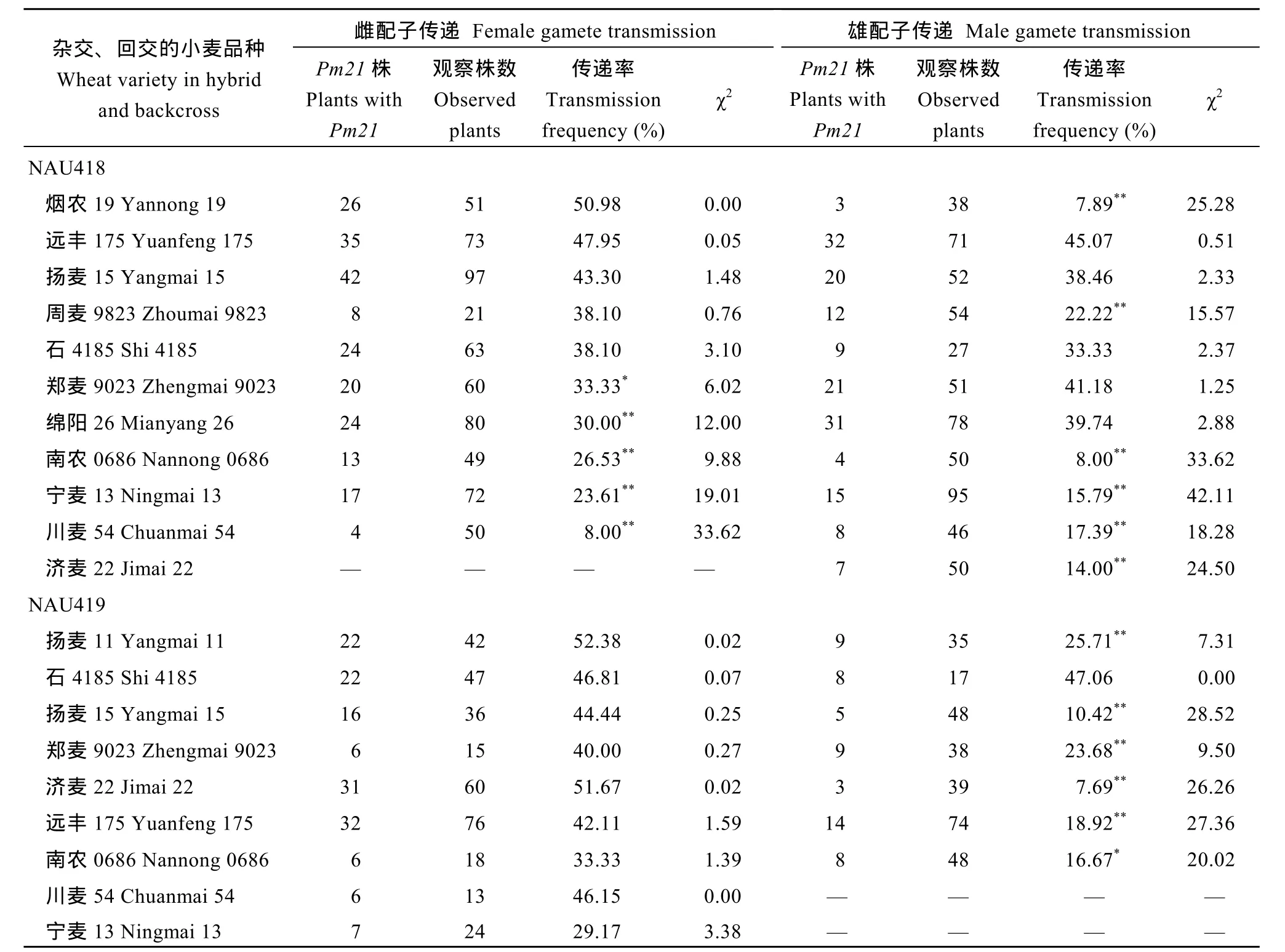
表2 T1AS.1AL-6VS (NAU418)和T4BS.4BL-6VS-4BL (NAU419)易位染色体在不同小麦背景中通过雌雄配子的传递率Table 2 Gamete transmission frequencies of T1AS.1AL-6VS (NAU418) and T4BS.4BL-6VS-4BL (NAU419) in different wheatbackgrounds
3 讨论
研究外源染色体在小麦背景中的传递对于利用外源染色体所携优异基因具有重要意义。NAU418 和NAU419均包含有抗白粉病基因Pm21, 利用与抗白粉病基因Pm21紧密连锁的分子标记CINAU15对后代植株进行鉴定可以推断外源目标片段的存在与否, 对小片段易位系NAU418和小片段中间插入易位系NAU419与不同小麦品种的杂种F1代以及相应BC1F1群体检测发现, 易位染色体能够基本正常参与配对和分离, 能够通过雌、雄配子传递, 但通过雄配子的传递率低于通过雌配子的传递率, 其原因可能是在受精过程中含有外源小片段易位染色体的配子与不含易位染色体的正常雌、雄配子之间存在受精竞争, 含有外源染色体的雄配子的竞争能力稍差于正常雄配子; 在减数分裂后期含有外源片段的易位染色体有可能出现分离落后现象, 致使外源染色体丢失。此外, 导入小麦背景中的外源染色体或染色体片段在雌雄配子中的传递, 常常因外源染色体的身份、其易位的小麦染色体身份、外源染色体片段大小和插入方式以及小麦品种的背景而不同。如李桂萍等[7]研究小麦–簇毛麦6VS/6AL易位染色体在百农64、百农9310、邯5310、小偃54、淮麦20、徐麦856背景中的遗传稳定性及其在配子中的传递,发现6VS/6AL易位染色体通过雄配子的传递率均低于通过雌配子的传递率。王海燕等[12]分析小麦–簇毛麦T4DL/4VS易位染色体在郑麦9023、周麦9823、绵阳26、石4185、扬麦15中的遗传稳定性及其在配子中的传递, 结果表明, 在不同组合的F2分离群体中, T4DL.4VS易位染色体在不同小麦遗传背景中的遗
传方式不相同, T4DL.4VS易位染色体的传递受到小麦遗传背景的影响。张文俊等[11]在研究携带黑麦6R染色体的配子在小麦背景中传递率时, 发现6R通过雄配子的传递率(10.3%)略高于通过雌配子的传递率(8.8%), 二者差异不显著, 且传递率均较低。张悦等[16]在研究中间偃麦草2Ai-2染色体在小麦部分同源染色体代换背景中的遗传时, 发现2Ai-2染色体通过雌配子的传递率达21.4%~52.9%, 通过雄配子的传递率达41.2%~69.3%, 依不同遗传背景而异。刘琳等[17]对偏凸山羊草6Mv染色体在不同四川小麦品种中的传递分析, 发现通过雌、雄配子的传递率平均分别为35.18%和31.72%, 传递频率差异不显著。这些研究说明不同外源染色体的传递率差异较大, 而且因不同遗传背景而异。本研究也有同样发现, 除了易位染色体本身之外, 回交亲本的遗传背景对后代偏分离都可能有一定影响。
小麦–簇毛麦纯合易位系NAU418和NAU419是小麦抗白粉病育种的理想抗源。本研究发现其中易位染色体都可以基本稳定遗传, 可以作为抗病育种的优良亲本, 通过直接与其他品种杂交, 将其所携的优异基因转移到待改良的小麦品种中。考虑到其中易位染色体通过雌、雄配子传递率的差异, 在利用易位染色体所携的优异基因时, 最好以该易位系为母本以提高该染色体在杂种后代中的传递率。同时, 本研究还发现, 易位系NAU419中的小片段中间插入易位染色体通过雌配子的传递率, 比小片段顶端易位系NAU418中的小片段顶端易位染色体通过雌配子的传递率要高, 进一步说明在育种上有用的易位系最好是携带有用基因的小片段中间插入易位, 它们在细胞学和遗传学上比较稳定且容易通过基因重组转入栽培品种并遗传给后代。
4 结论
小麦–簇毛麦纯合易位系NAU418和NAU419中的易位染色体都可以稳定遗传, 可以作为抗病育种亲本, 通过直接与其他品种杂交, 将其所携的优异基因转移到待改良的小麦品种中。在利用易位染色体所携的优异基因时, 最好以该易位系为母本以提高易位染色体在杂种后代中的传递率。
References
[1] 谢超杰, 杨作民. 孙其信. 小麦抗白粉病基因. 西北植物学报, 2003, 23: 822−829
Xie C J, Yang Z M, Sun Q X. Resistance genes to powdery mildew in wheat. Acta Bot Boreali-Occident, 2003, 23: 822−829 (in Chinese with English abstract)
[2] Gradzielewska A. The genus Dasypyrum: 2. Dasypyrum villosum, a wild species used in wheat improvement. Euphytica, 2006, 152: 441−454
[3] Chen P D, Qi L L, Zhou B, Zhang S Z, Liu D J. Development and molecular cytogenetic analysis of wheat–Haynaldia villosa 6VS/6AL translocation lines specifying resistance to powdery mildew. Theor Appl Genet, 1995, 91: 1125−1128
[4] Qi L L, Cao M S, Chen P D, Li W L, Liu D J. Identification, mapping, and application of polymorphic DNA associated with resistance gene Pm21 of wheat. Genome, 1996, 39: 191−197
[5] Cao A Z, Xing L P, Wang X Y, Yang X M, Wang W, Sun Y L, Qian C, Ni J L, Chen Y P, Liu D J, Wang X E, Chen P D. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc Natl Acad Sci USA, 108: 7727−7732
[6] Chen P D, You C F, Hu Y, Chen S W, Zhou B, Cao A Z, Wang X E. Radiation-induced translocations with reduced Haynaldia villosa chromatin at the Pm21 locus for powdery mildew resistance in wheat. Mol Breed, 2013, 31: 477−484
[7] 李桂萍, 陈佩度, 张守忠, 赵和. 小麦–簇毛麦6VS/6AL易位染色体对小麦农艺性状的影响. 植物遗传资源学报, 2011, 12: 744−749
Li G P, Chen P D, Zhang S Z, Zhao H. Effects of the 6VS/6AL translocation chromosome on agronomic characteristics of wheat. J Plant Genet Resour, 2011, 12: 744−749 (in Chinese with English abstract)
[8] Smith J D. The effect of chromosome number on competitive ability of hexaploid wheat gametophytes. Can J Genet Cytol, 1963, 5: 200−226
[9] Efremova T T, Maystrenko O I, Laykova L I. Development of alien substitution lines of wheat with rye chromosome 5R. Cereal Res Commun, 1996, 24: 33−39
[10] Whelan E D, Conner P R L, Thomas J B. Transmission of a wheat alien chromosome translocation with resistance to the wheat curl mite in common wheat, Triticum aestivum L. Can J Genet Cytol, 1986, 28: 294−297
[11] 张文俊, 景健康, 胡含. 黑麦6R染色体在小麦背景中的传递.遗传学报, 1995, 22: 211−216
Zhang W J, Jing J K, Hu H. The transmission of rye chromosome 6R in wheat background. Acta Genet Sin, 1995, 22: 211−216 (in Chinese with English abstract)
[12] 王海燕, 赵仁慧, 袁春霞, 张守忠, 肖进, 王秀娥. 小麦–簇毛麦T4DL.4VS易位染色体在不同背景中的遗传稳定性及其在配子中的传递. 麦类作物学报, 2013, 33: 13−17
Wang H Y, Zhao R H, Yuan C X, Zhang S Z, Xiao J, Wang X E. Transmission of T4DL.4VS chromosome through gametes and its genetic stability in different genetic background. J Triticeae Crops, 2013, 33: 13−17 (in Chinese with English abstract)
[13] Zhang P, Li W, Friebe B, Gill B S. Simultaneous painting of three genomes in hexploid wheat by BAC-FISH. Genome, 2004, 47: 979−987
[14] Sharp P J, Cao S, Desai S, Gale M D. The isolation characterization and application in the Triticeae of a set of wheat RFLP probes identifying each homoeologous chromosome arm. Theor
Appl Genet, 1989, 78: 342−348
[15] Cao A Z, Wang X E, Chen Y P, Zou X W, Chen P D. A sequence-specific PCR marker linked with Pm21 distinguishes chromosomes 6AS, 6BS, 6DS of Triticum aestivum and 6VS of Haynaldia villosa. Plant Breed, 2006, 125: 201−205
[16] 张悦, 林志珊, 曹保久, 郭义强, 王美蛟, 叶兴国, 辛志勇, 徐琼芳, 郭世华. 2Ai-2染色体在小麦部分同源染色体代换背景中的遗传. 作物学报, 2009, 35: 424−431
Zhang Y, Lin Z S, Cao B J, Guo Y Q, Wang M J, Ye X G, Xin Z Y, Xu Q F, Guo S H. Genetic behaviour of Thinopyrum intermedium chromosome 2Ai-2 in different wheat chromosome substitution backgrounds of group 2. Acta Agron Sin, 2009, 35: 424−431 (in Chinese with English abstract)
[17] 刘琳, 邓光兵, 易玲, 李林, 赵柳笛, 龙海, 潘志芬, 余懋群. 偏凸山羊草6Mv染色体在不同四川小麦品种中的传递. 应用与环境生物学报, 2010, 16: 50−53
Liu L, Deng G B, Yi L, Li L, Zhao L D, Long H, Pan Z F, Yu M Q. Transmission of chromosome 6Mvfrom Aegilops ventricosa in Sichuan wheat varieties. Chin J Appl Environ Biol, 2010, 16: 50−53 (in Chinese with English abstract)
URL: http://www.cnki.net/kcms/detail/11.1809.S.20151218.0915.002.html
Transmission and Genetic Stability of No-homoeologous Small Fragment Wheat–Haynaldia villosa Translocation Chromosomes with Pm21 in Various Cultivar Backgrounds of Common Wheat
WANG Hai-Yan, XIAO Jin, YUAN Chun-Xia, XU Tao, YU Chun-Yan, SUN Hao-Jie, CHEN Pei-Du, and WANG Xiu-E*
State Key Laboratory of Crop Genetics and Germplasm Enhancement / Cytogenetics Institute, Nanjing Agricultural University / Jiangsu Collaborative Innovation Center for Modern Crop Production, Nanjing 210095, China
Abstract:The powdery mildew resistance gene Pm21 comes from a diploid wheat related species, Haynaldia villosa. Two Pm21-carrying small fragment translocation lines, the terminal translocation line NAU418 and the small interstitial translocation line NAU419, have been developed. Both lines are highly resistant to powdery mildew and serve as new genetic resources for improvement of disease resistance. For understanding the transmission rate of the translocation chromosomes through male and female gametes and the genetic stabilities in different wheat genetic backgrounds, the two translocations were crossed to 12 common wheat varieties from different wheat growing areas of China. The F1hybrids were then backcrossed as reciprocally. Chromosome configurations of the obtained F1s were analyzed by fluorescence in situ hybridization (FISH) of the PMC at MI. It was found that the translocation chromosomes formed rod bivalents with their corresponding wheat chromosomes. Test crosses showed that the translocation chromosomes T1AS.1AL-6VS and T4BS.4BL-6VS-4BL could be transmitted to their offspring. The
transmission frequency of T1AS.1AL-6VS was higher through female gametes an average of 33.20%, ranging from 0.08% to 50.98% than through male gametes an average of 23.75%, ranging from 0.14% to 45.07%. Similarly, the transmission frequency of T4BS.4BL-6VS-4BL was higher through female gametes an average of 42.90%, ranging from 29.17% to 52.38% than through male gametes an average of 21.45%, ranging from 7.69% to 47.06%. These results show that the translocated chromosomes could be transmitted through male and female gametes, while genetic background has influences on the transmission rate, especially through male gametes.
Keywords:Wheat; Haynaldia villosa; Powdery mildew; Small fragment translocation; Gamete transmission frequency
收稿日期Received(): 2015-06-11; Accepted(接受日期): 2015-11-20; Published online(网络出版日期): 2015-12-18.
通讯作者*(Corresponding author): 王秀娥, E-mail: xiuew@njau.edu.cn, Tel: 025-84395308
DOI:10.3724/SP.J.1006.2016.00361
