结球甘蓝与油菜杂交胚挽救及其后代倍性鉴定
2016-02-14赵艳艳蒋武生原玉香王志勇姚秋菊魏小春张晓伟河南省农业科学院园艺研究所河南郑州450002
赵艳艳,蒋武生,原玉香,王志勇,姚秋菊,魏小春,张 强,张晓伟(河南省农业科学院 园艺研究所,河南 郑州 450002)
结球甘蓝与油菜杂交胚挽救及其后代倍性鉴定
赵艳艳,蒋武生,原玉香,王志勇,姚秋菊,魏小春,张 强,张晓伟*
(河南省农业科学院 园艺研究所,河南 郑州 450002)
为将油菜中根肿病抗性基因CRR导入到甘蓝从而获得抗根肿病的结球甘蓝材料,以二倍体结球甘蓝与四倍体人工合成油菜为亲本进行正反杂交,采用MS和B5培养基对杂交子房和胚珠进行胚挽救试验,研究不同取样时期、培养基、基因型以及暗处理对胚珠萌发率的影响,并采用流式细胞仪法对杂交后代幼苗倍性进行初步鉴定。结果表明:胚珠培养优于子房培养。以人工合成油菜为母本的杂交组合获得的杂种苗数(57株)远多于反交组合(仅获得7株杂种苗)。子房培养取材时间以蕾期授粉后第10天为宜,以B1(B5+6-BA 1.5 mg/L+NAA 0.2 mg/L+水解酪蛋白0.5%+活性炭0.5%)培养基最好(萌发数6株);胚珠培养适宜取材时间为蕾期授粉后第11天,以M1(MS+6-BA 1.0 mg/L+NAA 0.1 mg/L+水解酪蛋白0.5%+活性炭0.5%)培养基最佳(萌发率13.55%)。胚珠经过暗处理(24 h)后萌发率明显提高。倍性鉴定结果显示,胚挽救获得的64株幼苗中有57株为三倍体植株(89.06%)。
结球甘蓝; 人工合成油菜; 远缘杂交; 胚挽救; 倍性鉴定
结球甘蓝(BrassscaoleraceaL.var.capitata,2n=2x=18,CC)又名包菜、洋白菜、卷心菜、包心菜等,是十字花科芸薹属的植物,具有适应性广、耐寒和耐热性较强、病害少、产量高、营养丰富、抗癌、抗诱变、耐贮运等特点[1-2],是我国的一种重要蔬菜。其种植面积以每年约90万hm2的速度递增[3]。随着其种植面积的不断扩大,十字花科根肿病也不断蔓延,培育抗根肿病的甘蓝新品种是解决根肿病危害最有效的方法。而种间杂交是创造作物新种质的有效途径,研究人员通常利用胚挽救技术克服远缘杂交中幼胚死亡或败育的问题[4]。在十字花科蔬菜育种过程中胚挽救技术得到很好的应用并取得了明显的成绩,如利用胚珠培养在改良式H培养基上获得萝卜×大白菜属间杂种9株,但子房培养未得到杂种[5];利用子房培养在添加6-BA 和NAA的B5培养基上得到甘蓝型油菜×羽衣甘蓝杂种3株[6];用萝卜雄性不育系与甘蓝杂交,通过幼胚培养得到胞质雄性不育杂种[7];刘忠松等[8]先用芥菜型油菜与甘蓝型油菜杂交,再用甘蓝型油菜回交,在回交后代中获得了高油酸含量的甘蓝型油菜材料;扈新民等[9]以甘蓝显性雄性不育系 C08-11为母本,以萝卜自交系Q07-12为父本进行属间杂交,通过胚珠培养获得 29株杂交苗,子房培养仅获得 3株杂交苗;祝朋芳等[10]通过胚挽救,获得了大白菜与羽衣甘蓝杂交后代39株等。利用远缘杂交手段,可以实现抗病性从一个物种转移到另一个物种。如Snowdon等[11]将抗黑胫病基因从白芥转移到了甘蓝型油菜;2004年国外研究人员应用胚挽救培养甘蓝品种与芥菜型油菜 A19182抗黑腐病的种间杂种[12],通过胚挽救技术将芥菜的白粉病抗性转移到甘蓝中获得成功[13]。十字花科根肿病是一种重要的世界性植物土传真菌性病害,严重影响结球甘蓝的可持续生产,而甘蓝型油菜人工合成种(BrassicanapusL.)遗传多样性十分丰富[14]。鉴于此,本研究采用含根肿病抗性基因CRR的人工合成油菜(异源四倍体)与不含抗性基因的结球甘蓝材料(二倍体)人工杂交,由于杂交后代为三倍体,不结实,需要对杂交后代进行胚挽救,以期将根肿病抗性基因CRR导入到结球甘蓝材料中培育出抗根肿病的甘蓝新材料。
1 材料和方法
1.1 试验材料
甘蓝材料C308、C318、C315、C365由河南省农业科学院园艺研究所蔬菜研究室提供;人工合成油菜 (BrassicanapusL.,2n=36,RR)W01、W02、W03、W04由加拿大曼尼托巴大学栗根义教授惠赠,抗根肿病。
1.2 试验方法
试验于2013年4月在河南省农业科学院现代化农业基地内进行,将甘蓝与人工合成油菜分别与其他作物隔离种植,常规栽培管理。分别以结球甘蓝、人工合成油菜作母本,在初花期于杂交前2~3 d摘除母本花序上已开放的花朵,套袋隔离。于套袋父本开花当天取花药,同时对母本植株上3 mm左右的花蕾进行剥蕾授粉。授粉后套袋隔离,严格控制父本以外的花粉污染。分别于杂交后9、10、11、12、13、14 d取子房和胚珠进行离体培养。试验设计MS和B5基本成分添加不同浓度的激素、水解酪蛋白(CH)、活性炭(AC)等共10种培养基。
1.2.1 培养基成分对子房培养的影响 分别剪取W01×C308、W01×C318蕾期授粉后10 d的子房,用70%乙醇浸泡30 s,再用10%HgCl2消毒6 min,然后用无菌水冲洗4次,每次5 min,最后在无菌条件下分别接种至B0、B1、B2、B3、B4培养基上(表1)培养,每个处理15个子房,共计150个子房。培养基在高温灭菌前pH值调至5.8,121 ℃高压灭菌20 min后备用。接种子房的培养皿置于温度(24±2)℃、光照强度1 500 lx、光照 12 h/d的环境中培养。接种36 d后剥出幼胚继续培养,最后统计出苗情况。
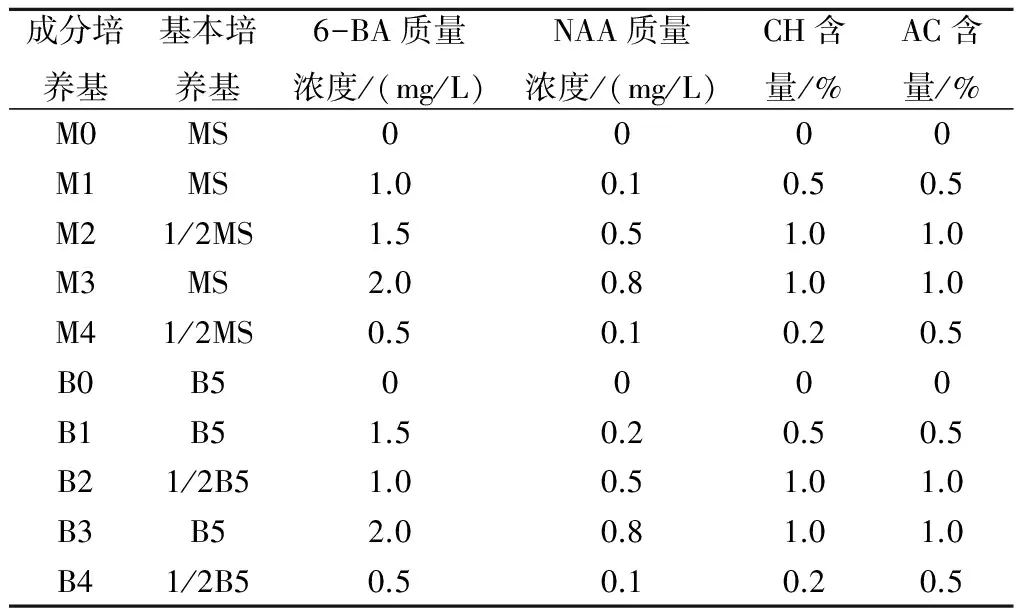
表1 培养基成分
1.2.2 培养基成分对胚珠培养的影响 取W01×C308蕾期授粉后11 d的胚珠,采用与1.2.1中子房培养相同的消毒方法,在无菌条件下将角果放于无菌滤纸上剥出幼胚接种至固体培养基B0、B1、B2、B3、B4、M0、M1、M2、M3、M4上(表1)培养,培养基在高温灭菌前pH值调至5.8,121 ℃高压灭菌20 min后备用。培养条件同子房培养。接种10 d后调查胚珠萌发情况。
1.2.3 取材时期对子房培养的影响 取W01×C308蕾期授粉后9、10、11、12、13、14 d的子房接种至B1培养基上培养,接种36 d后剥出幼胚继续培养,最后统计出苗情况。
1.2.4 取材时期对胚珠培养的影响 取W01×C308蕾期授粉后9、10、11、12、13、14 d的胚珠接种至M1培养基上培养,接种10 d后调查胚珠萌发情况。
1.2.5 不同基因型及正反交之间的胚珠萌发情况比较 取W01与C308、W02与C318、W03与C315、W03与C315、W04与C365正反交蕾期授粉后11 d的胚珠,接种至M1培养基上培养,接种10 d后统计胚珠萌发情况。
1.2.6 暗处理对胚珠培养萌发率的影响 取W01×C308蕾期授粉后11 d的胚珠接种至M1培养基上,分别在光照强度1 500 lx(正常处理)和24 h暗培养后于光照强度1 500 lx(暗处理24 h)条件下继续培养,接种10 d后统计胚珠萌发情况。
1.2.7 利用流式细胞仪对胚挽救杂种后代组培苗进行倍性鉴定 为了确认胚挽救成株材料是否为杂合体,取胚挽救杂种后代组培苗的幼嫩叶片,用德国PARTEC公司生产的流式细胞仪(CyFlowRCube 8)检测植株倍性,本试验选用四倍体人工合成油菜做为模板,调整电压为523 V,根据峰值出现的位置判断植株的倍性[15-16]。
2 结果与分析
2.1 不同培养基成分对子房培养和胚珠培养萌发的影响
2.1.1 子房培养 从表2可以看出,W01×C308、W01×C318子房培养在所有供试处理中仅有7个萌发,培养基以B5培养基添加适当的生长调节剂、水解酪蛋白和活性炭更有利于子房生长发育,其中以B1(6-BA 1.5 mg/L +NAA 0.2 mg/L+CH 0.5%+CA 0.5%)培养基中的子房出胚最好,共计萌发6个,最终发育成株。未添加水解酪蛋白、活性炭以及生长调节剂的培养基上,子房逐渐萎缩,甚至停止生长,有愈伤组织形成;而在添加水解酪蛋白、活性炭以及生长调节剂的培养基上,子房都能伸长、膨大(图1),但并不是都能萌发。在B1培养基上培养的W01×C308比W01×C318多2株萌发苗,说明基因型不同也影响子房萌发情况。
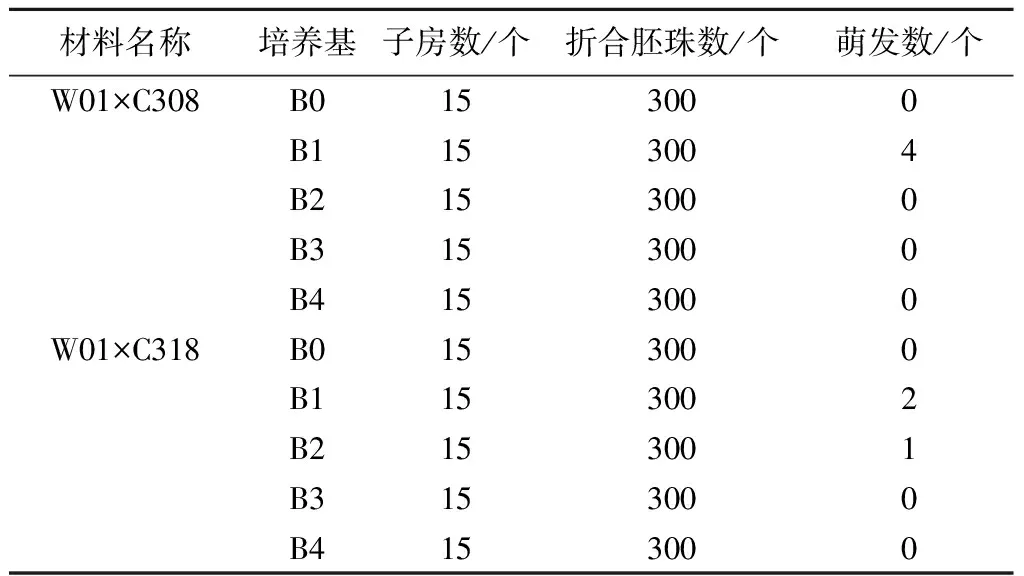
表2 不同基因型子房在不同培养基上的萌发情况
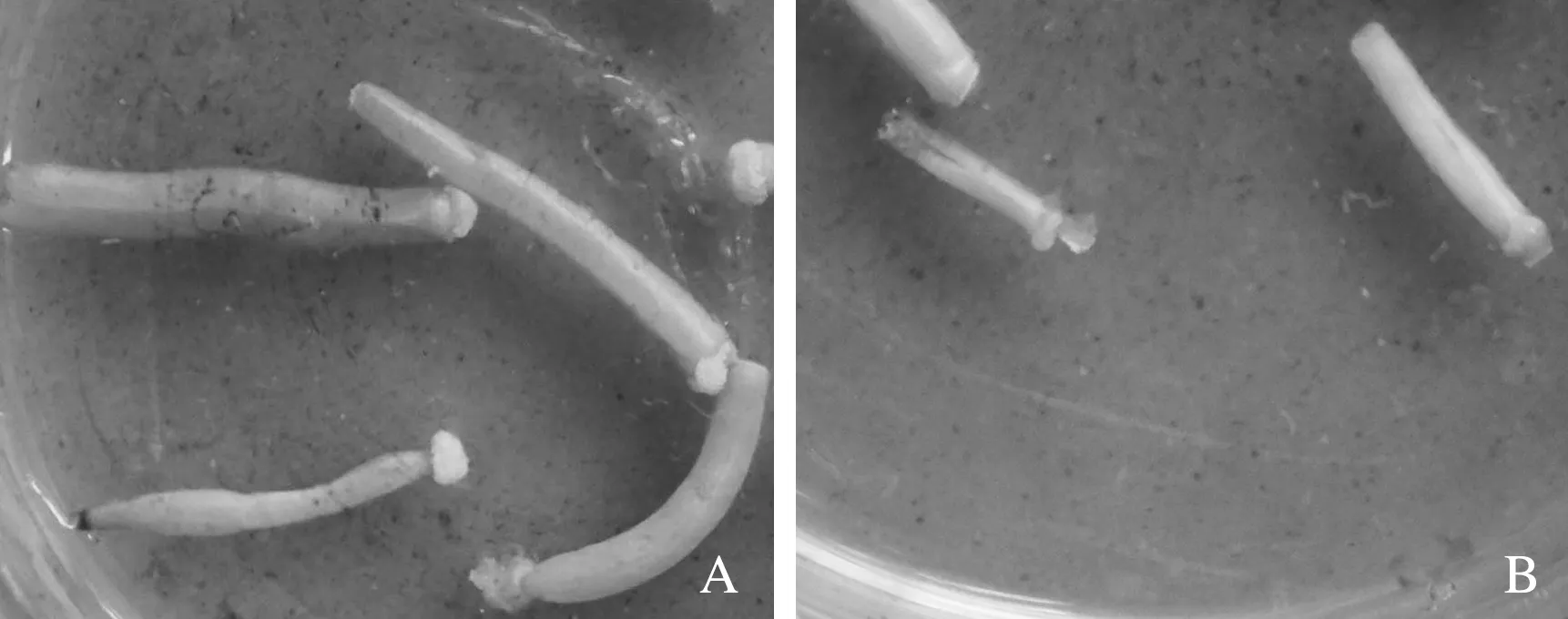
A.添加生长调节剂的培养基; B.未添加生长调节剂的培养基图1 生长调节剂对结球甘蓝×人工合成油菜离体子房培养的影响
2.1.2 胚珠培养 从表3可以看出,胚珠培养以MS基本培养基添加适当的生长调节剂、水解酪蛋白和活性炭更有利于胚珠萌发,高浓度生长调节剂影响胚的正常发育。以M1培养基(6-BA 1.0 mg/L+NAA 0.1 mg/L+CH 0.5%+CA 0.5%)中的胚珠萌发最好,萌发率为7.74%,其次为M2 和B1,萌发率分别为3.33%和3.23%,M4萌发率为2.25%。添加高浓度生长调节剂的MS培养基和B5培养基萌发率均较低,分别为0.89%(M3)和0.61%(B3)。总的来看,杂交胚珠离体培养在萌发率上明显优于子房培养,且杂交胚的生长情况较好(图2)。所有处理共计获得38株苗,按一个子房里面有20个胚珠计算,同等条件下子房培养才获得7株苗(表2)。
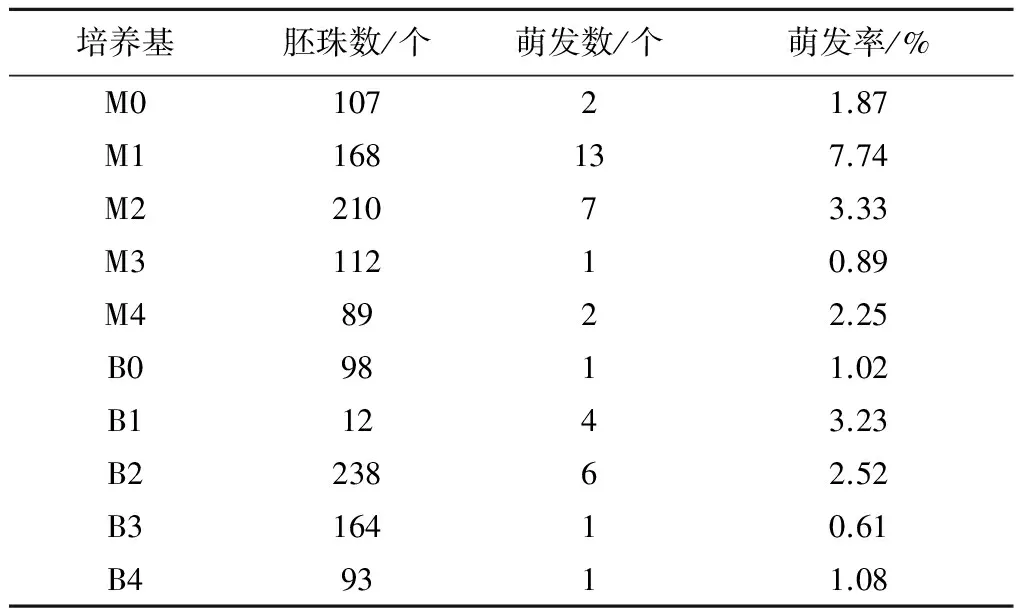
表3 W01×C308胚珠在不同培养基上的萌发情况

图2 结球甘蓝×人工合成油菜杂交胚珠生长情况
2.2 不同取材时期对子房培养和胚珠培养萌发情况的影响
2.2.1 子房培养 取W01×C308蕾期授粉后9、10、11、12、13、14 d的子房接种至B1培养基上培养。从表4可以看出,子房培养取材时间以授粉后第10天为宜,萌发率达到10.00%;取材时间为授粉后第9、11、12天的萌发率分别为3.33%、6.67%、3.33%,第13、14天未出胚,可能是由于杂种胚已逐渐败育所致,从子房中无法获得营养。
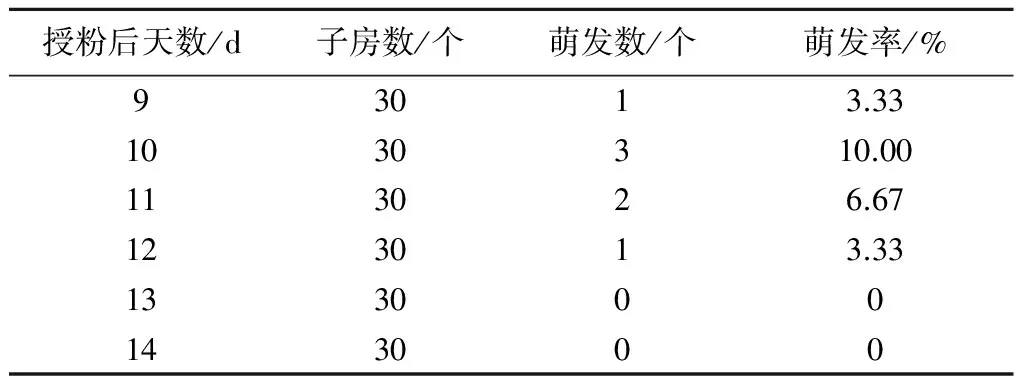
表4 不同取材时期W01×C308的子房在B1培养基上的萌发情况
2.2.2 胚珠培养 取材时间对胚珠培养非常重要,必须在胚珠发育到一定时期但未开始退化前取样,才能得到较好的萌发率。从表5可以看出,不同发育时期的胚珠在 M1培养基上培养10 d后,成活胚珠数有明显差别。胚珠培养取材时间以授粉后第11天为宜,萌发率高达13.55%;取材时间为授粉后第9、10、12、14天的萌发率分别为4.17%、8.25%、10.83%、7.00%、5.36%。授粉后第9天萌发率为4.17%,这可能是由于胚珠过小,过于依赖母体提供营养而难以离体培养;授粉后第14天仅有1个胚珠成活,萌发率为5.36%,这是由于随着受精后时间的延长,杂种胚已逐渐败育所致。因此,授粉后11 d是胚珠培养的适宜取材时间。胚珠培养出苗49株,子房培养出苗7株,按照每个子房25个胚珠计算,胚珠培养明显优于子房培养。
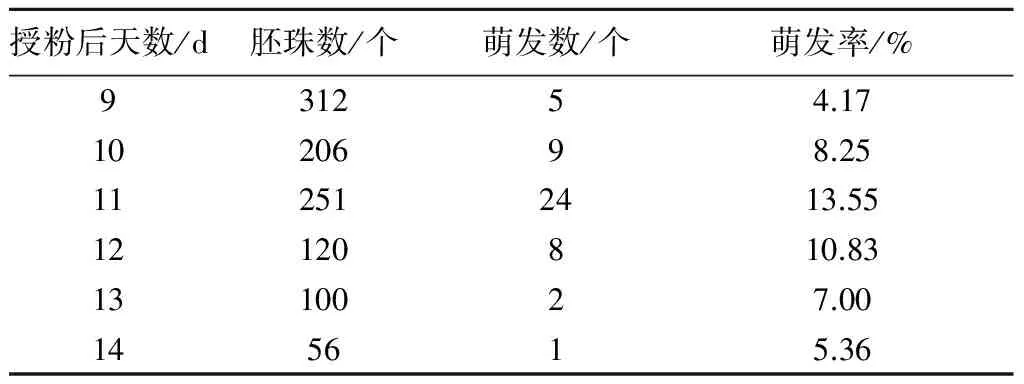
表5 不同取材时期W01×C308的胚珠在M1培养基上的萌发情况
2.3 不同基因型及正反交胚珠培养的萌发情况
从表6可以看出,在W01与C308、W02与C318、W03与C315、W04与C365等8个正反交组合中,除C365×W04外7个组合最终出胚成苗,共计出胚成苗64株,其中以人工合成油菜为母本的杂交组合获得的杂种苗数(57株)远多于反交组合(仅获得7株杂种苗)。对于本研究材料来说,不同基因型杂交组合的萌发率有显著差异。W01×C308萌发率最高,为16.0%,W02×C318、W03×C315、W04×C365的萌发率依次为 4.0%、5.5%、3.0%。C308×W01、C318×W02、C315×W03的萌发率依次为1.0%、1.0%、1.5%,C365×W04没有出胚。C308作父本萌发率仅为1%,而其作母本时达16.0%,因此为获得高萌发率,可考虑将C308作父本、人工合成油菜作母本。
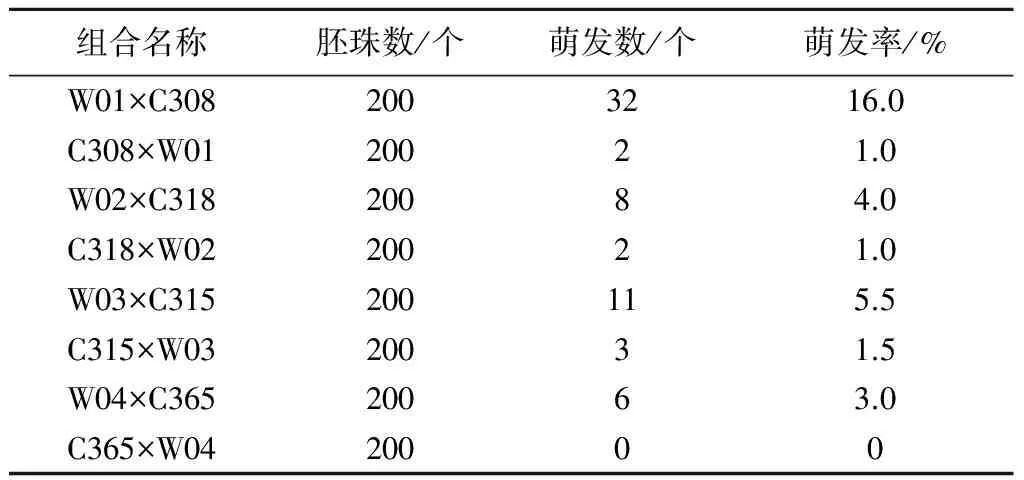
表6 各杂交组合后代胚珠培养的萌发情况
2.4 暗处理对胚珠培养萌发率的影响
从表7可见,W01×C308胚珠培养条件以暗处理24 h后再转入光照下培养效果较好,经过暗处理的萌发率达到17.5%,较未经暗处理的材料萌发率(12.0%)有明显提高。

表7 W01×C308的胚珠在正常处理和暗处理后的萌发情况
2.5 胚挽救杂种后代组培苗的倍性鉴定结果
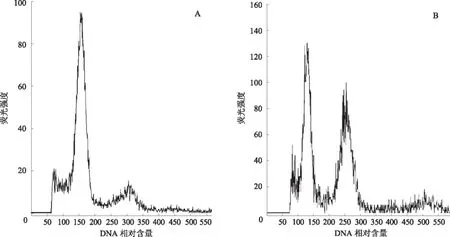
由图 3、4可见,二倍体结球甘蓝的荧光强度高峰出现在 100 位置,四倍体人工合成油菜的荧光强度高峰出现在200位置,而三倍体杂合体的对应值应该是2个亲本平均值,位置在150。通过流式细胞仪对培养获得的杂交植株进行倍性分析,64株胚挽救幼苗中有57株是三倍体真杂株,杂合率为89.06%,经过鉴定的植株需经过生根后移栽到大田进行进一步的生物性状鉴定和分子鉴定。
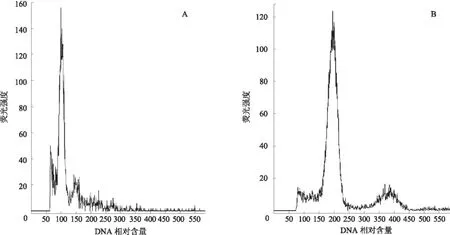
A为结球甘蓝;B为人工合成油菜图3 结球甘蓝、人工合成油菜DNA 相对含量的流式细胞分析图
A为杂合体;B为嵌合体图4 结球甘蓝×人工合成油菜部分杂交后代DNA相对含量的流式细胞分析图
3 结论与讨论
胚挽救包括胚珠培养、子房培养和幼胚培养,是克服受精之后远缘杂交不亲和性的有效方法[15]。程雨贵等[17]利用幼胚培养的方法获得了萝卜与甘蓝之间的杂种;胡大有等[18]通过子房培养获得了黑芥与诸葛菜的属间杂种;吴红美等[16]经胚挽救获得甘蓝型油菜与甘蓝种间杂种19株,经流式细胞仪鉴定11株真杂株;乔海云[15]利用胚珠培养获得了大白菜与紫甘蓝杂交后代,利用胚珠培养菜薹与芥蓝杂交获得杂交后代。本研究中将结球甘蓝与含根肿病抗性基因CRR的人工合成油菜杂交,采用子房培养和胚珠培养获得杂交后代,其中胚珠培养获得杂种的概率明显高于子房培养,这与前人研究结果[5,7,9,15]相符。
不同材料进行种间杂交,正反交的萌发结果有差异。乔海云[15]以菜薹为母本能获得少量种子,以芥蓝为母本未得到杂交株,以菜薹为母本的正交组合获得了 69 株幼苗;胡大有等[18]以黑芥为母本、诸葛菜为父本的正交组合获得了1株真杂种,而以诸葛菜为母本、黑芥为父本的反交组合未获得杂种;李俊等[19]以白菜和芥蓝进行正反杂交,研究结果显示,白菜×芥蓝的平均杂种获得率(2.32%)明显高于芥蓝×白菜的杂种获得率(1.16%);富贵等[20]将青海大黄油菜与甘蓝杂交,大黄油菜为母本的多个杂交组合获得 69株杂种苗,反交组合均未得到正常发育的胚[15-20]。文雁成等[21]对甘蓝型油菜与白菜型油菜进行种间杂交,结果显示,以甘蓝型油菜为母本的杂交结角率和亲和指数明显高于其反交组合。本研究表明,人工合成油菜作母本的萌发率高于作父本。W01作父本萌发率仅为1.0%,而其作母本时杂种胚萌发率高达16.0%,与前人[15-21]的研究结果一致。
本研究结果表明,结球甘蓝×人工合成油菜胚珠培养适宜取材时间为花期授粉后第11天,培养基以MS基本培养基添加适当的生长调节剂、水解酪蛋白和活性炭更有利于胚珠萌发,其中以M1培养基(MS+6-BA 1.0 mg/L+NAA 0.1 mg/L+水解酪蛋白0.5%+活性炭0.5%)中的胚珠萌发最好;子房培养取材时间以授粉后第10天为宜,培养基以B5培养基添加适当的生长调节剂、水解酪蛋白和活性炭更有利于子房生长发育,以B1(B5+6-BA 1.5 mg/L+NAA 0.2 mg/L+水解酪蛋白0.5%+活性炭0.5%)培养基中的子房出苗最好,但是不同材料之间杂交胚珠的萌发率有明显差异。胚珠经过暗处理(24 h)的萌发率较未经过暗处理的材料明显提高。
乔海云[15]、吴红美等[16]、王玉书等[22]利用流式细胞仪分别对十字花科杂种胚和羽衣甘蓝小孢子再生植株的DNA相对含量进行测定,同时采用细胞学方法对染色体数目进行鉴定,所得的结果一致,证明用流式细胞仪测杂种胚的结果是可信的。本研究中通过流式细胞仪对胚挽救获得的64株幼苗进行初步鉴定,确定57株为真杂株。
Harberd[23]采用离体胚培养获得甘蓝×白菜杂种,发现影响杂交胚珠培养的因素是多方面的,其中胚龄的大小对胚珠培养影响较大,一般来说胚龄越小,胚珠培养条件越复杂。胚挽救材料及其处理方式、培养基种类及培养条件、培养基渗透压和添加剂及胚龄等因素的变化均会直接影响到植物幼胚离体培养挽救的成功与否[4,24]。本研究结果也表明,胚龄是萌发率高低的关键,同时基因型、培养基、培养条件等对萌发率也有一定的影响[25],这几个方面缺一不可。用胚挽救技术提高杂种后代成活率的研究具有重要的现实意义,获得的胚挽救植株经过生根后移栽到大田需进一步进行生物性状鉴定和分子鉴定,期望能够获得抗根肿的结球甘蓝材料。
[1] 山东农业大学.蔬菜栽培学各论(北方本)[M].北京:中国农业出版社,2000:27-37.
[2] 叶恩赐,张欣文.卷心菜汁抗突变作用[J].上海铁道大学学报(医学版),1997,11(3):252.
[3] 杨丽梅,方智远,刘玉梅,等.“十一五”我国甘蓝遗传育种研究进展[J].中国蔬菜,2011(1):1-10.
[4] 贺佳玉,李云,姜金仲,等.植物胚败育机理及其离体培养挽救技术之研究进展[J].中国农学通报,2008,2(1):141-146.
[5] 赵德培.通过组织培养获得萝卜与大白菜的属间杂种[J].实验生物学报,1983,16(1):21-29.
[6] 陈树忠,殷家明,唐章林,等.甘蓝型油菜与羽衣甘蓝远缘杂交初步研究[J].西南农业大学学报,2000,22(3):208-210.
[7] 方智远,孙培田,刘玉梅.萝卜与甘蓝远缘杂交研究初报[J].园艺学报,1983,10(3):187-191.
[8] 刘忠松,官春云,李木旬,等.甘蓝型油菜与芥菜型油菜种间杂交研究[J].中国油料作物学报,2001,23(2):82-86.
[9] 扈新民,李锡香,梁燕,等.甘蓝与萝卜属间杂交及其胚挽救技术优化[J].中国蔬菜,2009(10):7-12.
[10] 祝朋芳,魏毓棠,张月.大白菜与羽衣甘蓝杂交胚的发育及其离体培养[J].园艺学报,2007,34(2):501-503.
[11] Snowdon R J,Winter H,Diestel A,etal.Development and characterisation ofBrassicanapus-Sinapisarvensisaddition lines exhibiting resistance toLeptosphaeriamaculans[J].Theor Apll Genet,2000,101(7):1008-1014.
[12] 向平.应用胚挽救培养甘蓝品种与芥菜型油菜A19182间抗黑腐病的种间杂种[J].作物育种信息,2005(6):8.
[13] 谢国禄.通过胚挽救技术将芥菜的白粉病抗性转移到甘蓝中[J].作物育种信息,2006(2):10.
[14] 刘平武,杨光圣.甘蓝型油菜人工合成种遗传多样性分析[J].作物学报,2004,30(12):1266-1273.
[15] 乔海云.芸薹种与甘蓝种杂交获得新种质的研究[D].北京:中国农业科学院,2012.
[16] 吴红美,徐跃进,万正杰.甘蓝型油菜与甘蓝种间杂种的鉴定及特性研究[J].华中农业大学学报,2011,30(3):290-294.
[17] 程雨贵,吴江生,陈洪高,等.萝卜和甘蓝远缘杂交研究[J].中国油料作物学报,2006,28(1):1-6.
[18] 胡大有,王爱云,吴金花.黑芥与诸葛菜远缘杂交研究[J].种子,2008,27(5):10-13.
[19] 李俊,罗莉霞,王转,等.人工合成具有白菜或甘蓝细胞质的甘蓝型油菜[J].作物学报,2010,36(8):1280-1285.
[20] 富贵,赵志刚,杜德志.利用青海大黄油菜和芥蓝合成大粒甘蓝型油菜[J].中国油料作物学报,2012,34(2):136-141.
[21] 文雁成,鲁丽萍,张书芬,等.利用十字花科种间杂交创造甘蓝型油菜种质资源的研究[J].河南农业科学,2014,43(6):30-34.
[22] 王玉书,王欢,高美玲,等.羽衣甘蓝小孢子再生植株的倍性鉴定及加倍[J].河南农业科学,2015,44(7):107-110,127.
[23] Harberd D J.A simple effective embryo culture technique forBrassica[J].Euphytica,1969,18(3):425-429.
[24] 陈纪鹏,陈婷,罗魏林.甘蓝型油菜与黑芥种间杂种的合成[J].河南农业科学,2014,43(7):43-47.
[25] 朱迎春,刘君璞,邓云,等.不同因素对西瓜花药愈伤组织诱导的影响[J].河南农业科学,2015,44(12):104-111.
Embryo Rescue ofBrassicaoleraceaL.var.capitata×BrassicanapusL.and Their Ploidy Identification
ZHAO Yanyan,JIANG Wusheng,YUAN Yuxiang,WANG Zhiyong,YAO Qiuju,WEI Xiaochun,ZHANG Qiang,ZHANG Xiaowei*
(Horticultural Institute,Henan Academy of Agricultural Sciences,Zhengzhou 450002,China)
For getting the cabbage materials which carried theCRRgene and was resistant to clubroot,the hybird ovary and ovules from the reciprocal crosses between diploidBrassscaoleraceaL.var.capitataand tetraploidBrassicanapusL.were cultured on the MS and B5 medium.Meanwhile,the effects of different sampling time,medium type,genotype and dark treatment on the germination rate of embryo were investigated.Furthermore,the ploidy of these hybirds was preliminarily determined by flow cytometer.The results showed that ovule culture was obviously better than ovary culture,57 embryos were produced inBrassicanapusL.×BrassscaoleraceaL.var.capitatcrosses,but 7 embryos was produced from the reciprocal crosses through embyro rescue.The best seedling from ovary culture on B1 (B5+6-BA 1.5 mg/L+NAA 0.2 mg/L+CH 0.5%+CA 0.5%) medium was obtained when inoculation was done 10 d after bud pollination.The highest embyro surviving rate (13.55%) by ovule culture on M1 (MS+6-BA 1.0 mg/L+NAA 0.1 mg/L+CH 0.5%+CA 0.5%) medium was obtained when inoculation was done 11 d after bud pollination.The surviving rate of the embryo was significantly increased after dark treatment for 24 h than that under normal culture.The identification results of cell ploidy showed that 57 seedlings from 64 seedlings rescued from this cross were triploid,accounting for 89.06%.
cabbage; synthetic rape; distant hybridization; embryo rescue; identification of ploidy
2016-03-18
国家大宗蔬菜产业技术体系项目(CARS-25-G-27);农业部重点实验室(站)项目(10205020);河南省科技攻关项目(142102110065);河南省专项资金项目(20157805);河南省2016年财政预算项目(20167907);河南省农业科学院自主创新专项基金项目(2016ZC25)
赵艳艳(1981-),女,河南临颍人,助理研究员,硕士,主要从事蔬菜育种研究。E-mail:zhaoyanyan9621@126.com
*通讯作者:张晓伟(1965-),男,河南平舆人,研究员,主要从事蔬菜育种研究。E-mail:xiaoweizhang5737@163.com
S565.2
A
1004-3268(2016)09-0088-06
