喀斯特山区2个鲫鱼群体基于转铁蛋白的遗传多样性研究
2016-02-09周其椿王金娜郭健康李建光
周其椿,安 苗,王金娜,郭健康,杨 兴,李建光
(1.贵州省水产研究所,贵州 贵阳 550025; 2.贵州大学 动物科学学院,贵州 贵阳 550025;3.正安县水产技术推广站,贵州 正安 563400)
喀斯特山区2个鲫鱼群体基于转铁蛋白的遗传多样性研究
周其椿1,安 苗2*,王金娜3,郭健康2,杨 兴1,李建光1
(1.贵州省水产研究所,贵州 贵阳 550025; 2.贵州大学 动物科学学院,贵州 贵阳 550025;3.正安县水产技术推广站,贵州 正安 563400)
为了解喀斯特山区2个不同鲫鱼群体在蛋白质水平的遗传多样性及其遗传差异,运用聚丙烯酰胺凝胶电泳方法对喀斯特山区2个群体鲫鱼的血清转铁蛋白电泳表型及多样性进行研究。结果表明:群体克隆多样性指数表现为草海鲫鱼(0.32)﹥红鲫(0.25,对照)﹥普安鲫鱼(0.13);3个鲫鱼群体的13个克隆由9个不同的等位基因产生,等位基因的频率分别为Tfa0.144 9、Tfb0.202 9、Tfc0.036 2、Tfd0.058 0、Tfe0.007 2、Tff0.239 1、Tfg0.014 5、Tfh0.275 4、Tfi0.029 0;普安鲫鱼与草海鲫鱼和红鲫无共享的基因型;普安鲫鱼与草海鲫鱼之间的遗传相似性指数较高(0.714 3),遗传距离较小(0.285 7)。总体来看,草海鲫鱼的克隆多样性水平较高,其转铁蛋白多样性程度较普安鲫鱼丰富;2个群体之间存在较近的亲缘关系,未达到遗传距离种的分化标准。
喀斯特山区; 鲫鱼; 转铁蛋白; 遗传多样性
鲫(Carassiusauratus) 隶属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲤亚科(Cyprininae)、鲫属(Carassius),是我国重要的经济鱼类,具有广泛的地理分布、众多的地方群体及丰富的遗传多样性。我国有2个种和1个亚种,即普通鲫鱼(C.auratus)、黑鲫(C.caraussius)和银鲫亚种(C.auratusgibelio)。贵州省地处西部高寒喀斯特山区,因其独特的地质地貌和气候条件,在漫长进化过程中,孕育了一批独特的地方性鲫鱼群体,如分布于西南部普安县青山镇一带山塘水体中(属南盘江水系,海拔1 650 m)的普安鲫鱼(C.auratus)和西北部威宁县草海国家级自然保护区(属金沙江水系,湖底海拔2 171.7 m)的草海鲫鱼(C.auratus)。
转铁蛋白(transferrin,简称Tf)是一种能够在机体内紧密且可逆地结合2个Fe3+、参与铁的运输和清除、直接关系到氧的利用和代谢的一种蛋白质。自Smithies等[1]首次在人血清中发现Tf的多态性以来,Tf的多态性在多种鱼类的血清中也得到证实。鱼类Tf仅存在于鱼类血清中,不受水温影响,与栖息水层有关[2]。同一品种鱼血清Tf的多态性、含量、活性及其基因频率和基因型频率等具有一定的稳定性,而不同种属、品种(品系)鱼的血清Tf有很大差异[2-3]。鱼类血清Tf已被众多学者广泛运用于养殖、野生鱼类的种质鉴定和种群遗传结构分析,并取得许多重要成果[2-6]。为更好地了解喀斯特山区鲫鱼群体的种质特性和遗传差异,对喀斯特山区2个鲫鱼群体的血清Tf多态性进行研究,以期揭示2个群体鲫鱼在蛋白质水平的遗传多样性及其遗传差异,为其选育和保护利用提供理论依据。
1 材料和方法
1.1 供试材料
普安鲫鱼采自贵州省普安县普安银鲫原种场饲养的后备亲鱼群体,草海鲫鱼采自贵州省威宁县草海湖泊的羊关山、邓家院子、潘家院子3个站点,对照红鲫采自贵州大学实验农场。供试鱼均为从渔获物混合群体中随机挑选的活泼健康个体,采用塑料袋密封充氧运回实验室水族箱暂养。3个群体基本情况见表1。从鱼体尾静脉取血0.8~1.0 mL,4 ℃静置5~10 min,3 000 r/min离心10 min,吸取上层血清于-20 ℃保存备用。

表1 试验鱼的样本数、体长及体质量
1.2 Tf样品制备及电泳
参照杨林[4]的方法纯化血清Tf,采用北京六一仪器公司DYCZ-21 型垂直电泳槽进行聚丙烯酰胺凝胶电泳(PAGE)。电极缓冲液为Tris(0.025 mol/L)-甘氨酸(0.192 mol/L)缓冲系统(pH值 8.3),浓缩胶含量为5%,电压为160 V,分离胶含量为8%,电压为200 V,上样量为10 ~15 μL,0.2%的溴酚蓝作指示剂,电泳时间约4 h。电泳结束后,将胶转入染缸中,清水冲洗,用0.2%的考马斯亮蓝R-250溶液(100 mL溶液中含0.2 g R-250、45 mL甲醇、10 mL冰乙酸、45 mL水)进行染色,用脱色剂(水、甲醇、冰乙酸溶液,体积比为5∶5∶1)对其进行脱色至背景清晰。
1.3 Tf电泳图谱的鉴定和多样性计算
参照李风波等[5]的方法鉴定每个群体鲫鱼的克隆组成,即每一种表型代表一种克隆。通过对每尾鲫鱼的血清Tf表型进行重复电泳检测,比对每尾鲫鱼的Tf电泳表型并进行表型克隆的判定,其克隆类型表示为C1、C2、C3…,计算其克隆多样性指数(克隆数/样品数)。为进一步验证3个群体鲫鱼不同克隆之间的相互遗传关系,将鉴定出来的15个克隆样品在同一胶板进行反复电泳对照;参照周秋白等[6]的方法进行Tf对应的基因和座位的命名,即Tf对应的基因和座位用斜体字母表示(如Tfa),多个等位基因依其电泳迁移速度的减小依次用小写字母表示(如a、b、c…)。并计算基因型频率、基因频率、遗传相似性指数和遗传距离。
基因型频率=特定基因型个体数/测定个体总数;
基因频率=特定等位基因出现数/测定个体所有等位基因出现总数;
遗传相似性指数(I)和遗传距离(D)计算公式为:
I=2Nij/(Ni+Nj)
D=1-I
式中,Nij是鲫鱼群体i和j之间共同的等位基因数,Ni、Nj分别是鲫鱼群体i和j全部的等位基因数。
2 结果与分析
2.1 3个群体鲫鱼血清Tf的PAGE表型及克隆组成
图1是32尾普安鲫鱼血清Tf的PAGE表型(图1a)和克隆组成(图1b)。32尾普安鲫鱼共鉴定出4个克隆,其中有1尾为克隆C1,有25尾共享克隆C2,4尾为克隆C3,另有2尾为克隆C4。通过计算可知,其群体克隆多样性指数为0.13。
图2是25尾草海鲫鱼血清Tf的PAGE表型(图2a)和克隆组成(图2b)。25尾草海鲫鱼共鉴定出8个克隆,其中有1尾为克隆C1,有1尾为克隆C2,2尾为克隆C3,12尾为克隆C4,有6尾共享克隆C5,1尾为克隆C6,1尾为克隆C7,1尾为克隆C8。通过计算可知,群体克隆多样性指数为0.32。
图3是12尾红鲫血清Tf的PAGE表型(图3a)和克隆组成(图3b)。12尾红鲫共鉴定出3个克隆,其中有1尾为克隆C1,有3尾共享克隆C2,剩余的8尾为克隆C3。通过计算可知,克隆多样性指数为0.25。

1—32分别为32尾普安鲫鱼; a为PAGE结果; b为克隆组成。下同

图2 草海鲫鱼血清Tf的PAGE结果及克隆组成
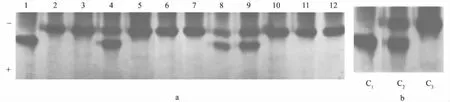
图3 红鲫血清Tf的PAGE结果及克隆组成
2.2 3个群体鲫鱼的基因型频率和基因频率
从图4可以看出,草海鲫鱼的克隆C7与红鲫的H2属相同的图谱,草海鲫鱼的克隆C5与红鲫的H3属相同的图谱,故共计有13个克隆。

图中C1—C8代表草海鲫鱼的8个克隆;P1—P4代表普安鲫鱼的4个克隆;H1—H3代表红鲫的3个克隆
本研究鉴定出的13个克隆共有Tfa、Tfb、Tfc、Tfd、Tfe、Tff、Tfg、Tfh和Tfi9个不同的等位基因(图4),其中,Tfe、Tfg为草海鲫鱼特有的等位基因,Tfi为普安鲫鱼特有的等位基因,草海鲫鱼与普安鲫鱼共享Tfa、Tfb、Tfd、Tff和Tfh5个等位基因。草海鲫鱼与红鲫共享等位基因Tfc,Tff为3个群体所共享等位基因。等位基因的频率分别为Tfa0.144 9、Tfb0.202 9、Tfc0.036 2、Tfd0.058 0、Tfe0.007 2、Tff0.239 1、Tfg0.014 5、Tfh0.275 4、Tfi0.029 0(表2)。
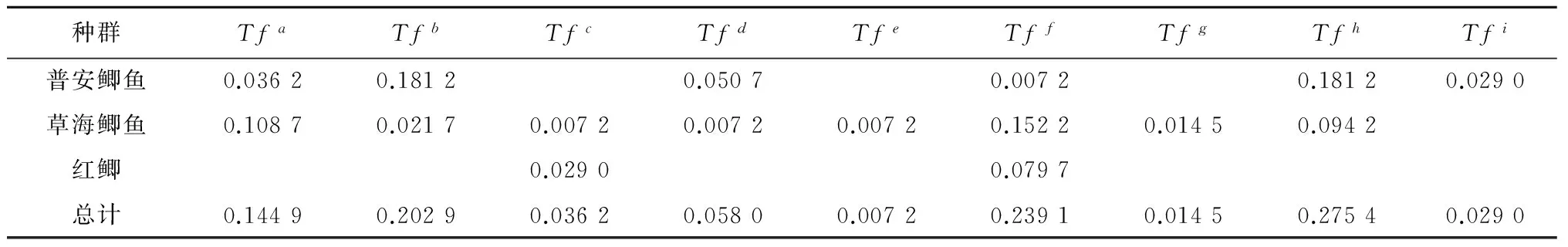
表2 鲫鱼群体等位基因频率
3个群体69尾个体共有13个基因型,Tfadf、Tfbh、Tfadi、Tfdd、Tfbb、Tfde、Tfabg、Tfafh、Tfff、Tfaf、Tfcf、Tffh、Tfcc。基因型频率分别为Tfadf0.014 5、Tfbh0.362 3、Tfadi0.057 9、Tfdd0.029 0、Tfbb0.014 5、Tfde0.014 5、Tfabg0.029 0、Tfafh0.174 0、Tfff0.202 9、Tfaf0.014 5、Tfcf0.057 9、Tffh0.014 5、Tfcc0.014 5(表3)。
32尾普安鲫鱼鉴定出来的4个不同克隆的基因型主要是由6个等位基因Tfa、Tfb、Tfd、Tff、Tfh、Tfi分布频率不同产生的。克隆C1的基因型为Tfadf、克隆C2的基因型为Tfbh、克隆C3的基因型为Tfadi、克隆C4的基因型为Tfdd;25尾草海鲫鱼鉴定出来的8个不同克隆的基因型主要是由Tfa、Tfb、Tfc、Tfd、Tfe、Tff、Tfg、Tfh8个不同的等位基因分布频率不同产生的,克隆C1的基因型为Tfbb、克隆C2的基因型为Tfde、克隆C3的基因型为Tfabg、克隆C4的基因型为Tfafh、克隆C5的基因型为Tfff、克隆C6的基因型为Tfaf、克隆C7的基因型为Tfcf、克隆C8的基因型为Tffh;12尾红鲫鉴定出来的3个克隆的基因型仅由Tfc、Tff2个等位基因分布频率不同产生,克隆C1的基因型为Tfcc、克隆C2的基因型为Tfcf、克隆C3的基因型为Tfff。草海鲫鱼与红鲫之间共享2个基因型Tfff和Tfcf。普安鲫鱼与草海鲫鱼和红鲫之间无共享的基因型。

表3 鲫鱼群体基因型分布及频率
注:括号内的数字为基因型个数。
2.3 3个群体鲫鱼的遗传相似性指数和遗传距离
遗传相似性指数和遗传距离是衡量物种间亲缘关系的重要指标。从表4可以看出,普安鲫鱼与草海鲫鱼之间的遗传相似性指数较高(0.714 3),遗传距离较小(0.285 7),表明两者之间存在较近的亲缘关系,未达到遗传距离种(0.3)的分化标准,而普安鲫鱼与红鲫间的遗传距离(0.750 0)高于草海鲫鱼与红鲫间的遗传距离(0.555 6)。

表4 鲫鱼群体遗传相似性指数和遗传距离
注:右上角为遗传相似性指数,左下角为遗传距离。
3 结论与讨论
3.1 2个地方鲫鱼群体Tf电泳表型
有关鲫鱼血清Tf的多态性研究发现,由于多个等位基因造成Tf表型在品系间存在一定的差异[5-7]。本研究中,2个鲫鱼群体血清Tf也存在多个等位基因,普安鲫鱼有6个等位基因,草海鲫鱼有8个等位基因,2个群体鲫鱼存在3个特有的等位基因Tfe、Tfi和Tfg,Tfe和Tfg为草海鲫鱼所特有,Tfi为普安鲫鱼所特有。正是由于Tfe、Tfi和Tfg等位基因在群体间的差异性分布造成2个群体鲫鱼的Tf表型在群体间均存在着一定的差异。
同时,本研究中普安鲫鱼鉴定的克隆C2和克隆C4与安苗等[8]研究中的克隆C4和克隆C6表型完全一致,再次说明Tf在鲫鱼同一群体克隆鉴别中具有很好的稳定性和重复性。
3.2 2个地方鲫鱼群体克隆多样性
由于Tf多态性在鱼类中广泛存在,克隆多样性作为衡量群体遗传多样性的指标已被广泛应用。本研究中普安鲫鱼的克隆多样性为0.13,略低于安苗等[8]对普安鲫鱼的研究报道中的克隆多样性指数(0.16),与其研究中侧线鳞为31的普安鲫鱼的多样性指数(0.14)较为相近,究其原因可能与采集的样本有关,本研究采集的样品为普安鲫鱼后备亲鱼,而普安鲫鱼原种场近年来后备亲鱼选育以侧线鳞30或30以上为标准。
25尾草海鲫鱼克隆数高达8种,群体克隆多样性指数为0.32,与国内外其他鲫鱼群体比较,其多样性远高于方正银鲫(0.2)[4]、关东鲫(0.13)[9]、淇河鲫(0.14)[10]和额尔齐斯河银鲫(0.15)[11]等,略低于伊犁河鲫鱼(0.39)[6]和武汉长江江段鲫鱼(0.42)[11]。
总体来看,草海鲫鱼的克隆多样性水平较高,其Tf多样性程度较普安鲫鱼的后备亲本丰富。
3.3 2个地方鲫鱼群体遗传及变异
本研究中2个群体鲫鱼也存在着一定的差异,32尾普安鲫鱼共有4种基因型,而25尾草海鲫鱼则多达8种基因型,并且群体间的基因型均不相同。
等位基因方面,两群体鲫鱼共享5个不同的等位基因,但其基因频率在两鲫群体鱼中存在一定的差异,如Tfa在草海鲫鱼中的频率为0.108 7,在普安鲫鱼中仅为0.036 2;而Tfb在草海鲫鱼中的频率只有0.021 7,在普安鲫鱼中则高达0.181 2;Tfd在草海鲫鱼中的频率为0.007 2,在普安鲫鱼则为0.050 7;Tff在草海鲫鱼中的频率为0.152 2,而在普安鲫鱼中仅有0.007 2;同样Tfh的频率也在两群体鲫鱼中存在差异。同时在2个群体中还存在着各自群体特有的等位基因,普安鲫鱼特有Tfi,草海鲫鱼特有Tfe、Tfg。
鱼类Tf的多态性与其生活环境和生活习性存在一定的相关性,不同属种、品种(品系)间由于其生活习性和生存环境的不同,血清Tf有很大差异,主要表现为存在各自Tf区的特定位置,其基因型和基因频率分布也各不相同[12-13]。因此本研究的结果表明,在复杂多变的生存环境中草海鲫鱼较普安鲫鱼更具良好的调控能力。
从本研究3个群体鲫鱼间遗传距离和相似性指数以及共享的等位基因结果来看,草海鲫鱼与普安鲫鱼相似性最高,与红鲫相似性次之,而红鲫与普安鲫鱼相似性最低。在鲫属鱼类的系统进化关系上,红鲫是一种标准的二倍体鲫鱼,在鲫鱼中进化地位较低,普安鲫鱼是一种进化地位较高的鲫鱼类,属雌核发育的三倍体鱼类[14]。而有关草海鲫鱼的初步研究表明[15-16],它是一种由二倍体和三倍体组成的混合群体。本研究结果表明,草海鲫鱼遗传组成较为复杂,既与普安鲫鱼有相似的地方,也存在不同之处;且草海鲫鱼与红鲫也存在一定的相似性。本研究结果也从蛋白质的水平证明草海鲫鱼是一种由二倍体和三倍体组成的混合群体。
[1] Smithies O,Hiller O.The genetic control of transferrin in humans[J].Biochem,1959,72:121-126.
[2] 龙华.一种新的鱼类种质鉴定法[J].水产科学,2004,23(3):16-17.
[3] 龙华,曾勇,郑英.转铁蛋白的研究与发展[J].生物工程进展,2001,21(2):32-39.
[4] 杨林.银鲫转铁蛋白及其相关分子标记的遗传多样性研究[D].武汉:中国科学院水生生物研究所,2000.
[5] 李风波,周莉,桂建芳,等.新疆额尔齐斯河水系银鲫克隆多样性研究[J].水生生物学报,2009,33(3):363-368.
[6] 周秋白,郑宇,周莉,等.新疆伊犁河鲫鱼遗传多样性初步研究[J].水生生物学报,2009,21(4):58-64.
[7] 甘云飞.萍乡红鲫转铁蛋白研究[D].南昌:南昌大学,2008.
[8] 安苗,黄仕洪,李勇.普安鲫群体克隆组成的初步研究[J].贵州农业科学,2012,40(9):176-179.
[9] Ohara K,Ariyoshi T,Sumida E,etal.Clonal diversity in the Japanese silver crucian carp,Carassiuslangsdorfiiinferred from genetic markers[J].Zoological Science,2003,20:797-804.
[10] Li F B,Gui J F.Clonal diversity and genealogical relationships of gibel carp in four hatcheries[J].Anim Genet,2008,39(1):28-33.[11] 李风波.银鲫不同群体的分布格局、遗传多样性和系统关系研究[D].武汉:中国科学院水生生物研究所,2007.
[12] Verspoor E.Spatial correlations of transferrin allele frequenciesin atlantic salmon(Salmosalar)populations from North America[J].Can J Fish Aquat Sci,1986,43: 1074-1078.[13] Yang L,Yang S T,Wei X H,etal.Genetic diversity among different clones of the gynogenetic silver Crucian carp,Carassiusauratusgibeliorevealed by transferrin and isozyme markers[J].Biochem Genetics,2001,39:213-225.
[14] 俞豪祥,宗琴仙,关宏伟,等.天然雌核发育普安鲫(A型)细胞遗传学和血清电泳的初步研究[J].水产科技情报,1991,18(5):130-133.
[15] 王金娜,李星浩,安苗,等.草海鲫鱼染色体核型分析[J].贵州农业科学,2013,41(2):134-137.
[16] 王金娜,周其椿,安苗,等.草海鲫鱼繁殖生物学特性的研究[J].水产科学,2013,32(12):701-705.
Genetic Diversity Study of Two Groups ofCarassiusauratusinKarst Mountainous Area Based on Transferrin
ZHOU Qichun1,AN Miao2*,WANG Jinna3,GUO Jiankang2,YANG Xing1,LI Jianguang1
(1.Guizhou Fisheries Research Institute,Guiyang 550025,China; 2.College of Animal Science,Guizhou University,Guiyang 550025,China; 3.Aquaculture Technology Centre of Zheng’an County,Zheng’an 563400,China)
In order to study the genetic diversity and genetic difference at protein levels of two kinds of crucian carp (Carassiusauratus) in Karst mountainous area,the electrophoresis phenotype of serum transferrin(Tf) and diversity were studied by the method of polyacrylamide gel electrophoresis.The results showed that the groups clonal diversity index showed Caohai crucian carp(0.32)﹥Red crucian carp(0.25)﹥Puan crucian carp(0.13).Thirteen phenotypes of three kinds of crucian carp were determined by nine kinds of different alleles, frequencies of allelic genes wereTfa0.144 9,Tfb0.202 9,Tfc0.036 2,Tfd0.058 0,Tfe0.007 2,Tff0.239 1,Tfg0.014 5,Tfh0.275 4,Tfi0.029 0,respectively.There were none shared genotype among three kinds of crucian carp.They had higher genetic similarity and smaller genetic distances between Puan crucian carp and Caohai crucian carp,which were 0.714 3 and 0.285 7,respectively.Researches demonstrated that clonal diversity was higher and the transferrin diversity level was richer in Caohai crucian carp than in Puan crucian carp.The genetic diversity was more complicated of Caohai crucian carp, and there were lots of similarities with Puan crucian carp, however,there were also differences between them.
Karst mountainous area;Carassiusauratus; transferrin; genetic diversity
2016-06-10
现代农业产业技术体系建设专项资金项目(CARS-46);贵州省农业科技攻关项目[黔科合NY字(2009)3070];贵州省科学技术基金项目[黔科合J字(2011)2223]
周其椿(1987-),男,贵州石阡人,研究实习员,硕士,主要从事水产动物遗传育种及种质资源研究与应用工作。 E-mail:zhou.qichun@163.com
*通讯作者:安 苗(1970-),男,贵州德江人,副教授,主要从事水产动物增养殖的教学和鱼类种质资源研究工作。 E-mail:gzuam@163.com
时间:2016-11-25 14∶24∶33
Q953
A
1004-3268(2016)12-0143-06
网络出版地址:http://www.cnki.net/kcms/detail/41.1092.S.20161125.1424.029.html
