利用配合力和SSR标记对玉米Tuxpeno种质遗传潜势的研究
2016-01-17郭向阳陈泽辉祝云芳王安贵陈建军刘鹏飞
郭向阳, 陈泽辉, 祝云芳, 王安贵, 胡 兴,2, 陈建军,2, 刘鹏飞,2
(1.贵州省旱粮研究所, 贵阳550006; 2.贵州大学农学院, 贵阳550025)
伴随着杂种优势利用技术以及现代育种研究的发展,我国玉米种质基础狭窄和利用效率低的问题突出[7]。Tuxpeno种族是世界玉米主要的一个种族,具有抗旱、耐瘠和耐涝能力较强,茎秆坚韧,抗斑病、锈病,Tuxpeno种质选系和改良系组配的杂交种也具备这些特点[1]。Hallauer认为,美国玉米带马齿来自北方硬粒和南方的马齿复合种,而南方的马齿复合种是来自于 墨 西 哥 的 Tuxpeno 种 质[12]。Hallauer等[12],陈泽辉等[4]认为,美国的瑞德种质是玉米带马齿的典型代表,而Tuxpeno在热带地区非常突出,因此,Tuxpeno种族也是热带、亚热带玉米非常重要的种质资源,是选育自交系和杂交种的重要原始材料。Tuxpeno种质与ETO或Suwan种质构成了热带地区玉米杂交种的主要杂种优势模式。Hallauer[13]提出了美国新的杂种优势模式BSSS-Tuxpeno×nonBSSS-nonTuxpeno,由此可见美国已经成功地运用Tuxpeno种质扩增了BSSS种质,还显示了杂种优势群和杂种优势模式是人为的、可以改变的属性。Well hausen[14]根据双列杂交分析,鉴别出4个比较好的种族群。Tuxpeno种群与其他3个种族群杂交,都表现很强的杂种优势。利用配合力和SSR标记对玉米Tuxpeno种质遗传潜势的研究国内还未见报道。本研究以Tuxpeno种质选系和测验种10份玉米自交系为材料,基于配合力和微卫星(SSR)标记的遗传潜势数据的评估,分析玉米Tuxpeno种质的配合力,SSR标记对10份玉米自交系的遗传变异及产量结果和遗传变异的关系;从而,最大限度地利用于育种研究;配合力高的材料可作为有利等位基因资源引入温带育种方案,拓宽种质基础,进一步提高产量和抗性;因此,开展外来种质与适应种质间的杂种优势和配合力分析,是玉米种质研究中必不可少的重要内容。
1 材料与方法
1.1 实验材料
选用贵州省旱粮研究所提供的Tuxpeno种质(5份Tuxpeno选系)为母本,我国五大类群中具有代表性骨干系为父本(测验种),采用NCⅡ设计组配25个杂交组合,以黔单16号为对照种(见表1)。
1.2 田间试验设计
2008年夏季,按NC-Ⅱ遗传交配设计,在贵州省农业科学院实验田,以5个Tuxpeno种质选系为母本,以我国玉米5个类群代表系(测验种)为父本,组配25个组合。当年冬季,在海南三亚崖城;2009年夏季,贵州省农业科学院(海拔在1 200m左右)2种生态环境下试验采用完全随机区组设计。分别对25个杂交组合和1个对照种进行田间鉴定,2行区,3次重复,行长3m,行距75cm,株距25cm,每行13株,小区面积为4.5m2。田间管理同大田生产。田间调查和室内考种以小区为单位。
用小区平均值为单位对10个农艺和产量性状作方差分析。用SASS 3.0软件对每个性状的数据处理结果进行方差分析[3,5-6]。
1.3 SSR实验方法
10份自交系均用于室内SSR实验部分,对玉米种质材料进行分类,分析玉米种质间的遗传相似系数,利用统计软件绘制聚类分析图。
1.3.1 SSR实验设计
玉米SSR引物序列信息来自MaizeGDB(http://www.maizegdb.org),引物由上海捷瑞公司合成,从55对SSR引物中共筛选出25对扩增条带清晰稳定的SSR。
1.3.2 SSR实验数据处理与分析
SSR扩增带型0、1、9统计,建立数据库。在相同迁移率位置上,有带记为1,无带记为0,缺失记为9。以Jaccard’s系数计算样本间的遗传相似系数(GS),即GS(jk)=a/(a+b+c),其中a指j和k共有的位点数;b指j有而k无的位点数;c指k有而j无的位点数。
统计分析是以SSR扩增产物结果为依据,有带记为1、无带记为0、缺失或带弱记为9,统计泳带建立数据库。按 UPGMA 方法,利用软件 DPS v 7.05版进行数据聚类,获得系统聚类图;以简单配对参数估计基因频率,SSR位点的多态性信息量(PIC)按公式PIC=1-∑fi2(其中fi为i位点的基因频率)计算,材料之间的遗传相似系数变化范围(GS)按公式GS=m/(m+n)(其中m为基因型间共有带数目,n为差异带数目)计算,遗传距离(GD)按公式GD=1-GS计算。
2 结果与分析
2.1 田间试验结果与分析
2.1.1 方差分析
本研究分别对两点所调查的10个农艺性状进行了不同处理的方差分析,包括区组、组合、地点、地点与组合互作、一般配合力(GCA)、特殊配合力(SCA)(见表2)。组合间方差分析结果(表2)表明:在调查的10个农艺性状均达到显著水平。进一步表明所有性状在25个组合之间差异极显著。因此,可以进行配合力方差分析。地点间方差分析表明:在调查的10个农艺性状中,除了穗行数、百粒重和株高未达到显著差异外,其他性状均达到极显著差异;表明大部分性状明显受生态条件和地域条件的影响。地点×组合互作间方差分析表明:在调查的10个农艺性状中;行粒数、百粒重、秃尖长、出籽率、穗位高和小区产量均达到了极显著水平,说明它们的地点与组合互作效应差异显著;而其他性状均未达到显著水平,进一步说明它们的地点×组合互作效应不显著。
Tuxpeno种质的一般配合力效应方差分析表明:调查性状中除小区产量外,其他性状均达到极显著水平;同时,调查的10个农艺性状的一般配合力(GCA)方差均大于特殊配合力(SCA)方差。进一步说明,5个Tuxpeno种质选育的自交系之间差异相对较大。我国五大种质骨干自交系的一般配合力(GCA)方差均达到极显著水平,且一般配合力(GCA)方差远大于特殊配合力(GCA)方差;说明调查的10个农艺性状方面,代表我国五大玉米类群的骨干自交系之间差异比较明显。5个Tuxpeno种质选系和5个骨干系间的特殊配合力(GCA)方差分析表明,穗行数、百粒重、秃尖长和穗位高均达到极显著水平,行粒数、穗长、出籽率和株高均达到显著水平,进一步说明非加性基因效应的作用比较重要;而其他性状均未达到显著水平,说明加性基因效应比非加性基因效应重要。综合以上表明,供试材料调查的性状受加性基因效应比非加性基因效应相对重要,这对选育高产稳产杂交种和高配合力的自交系有重要意义。
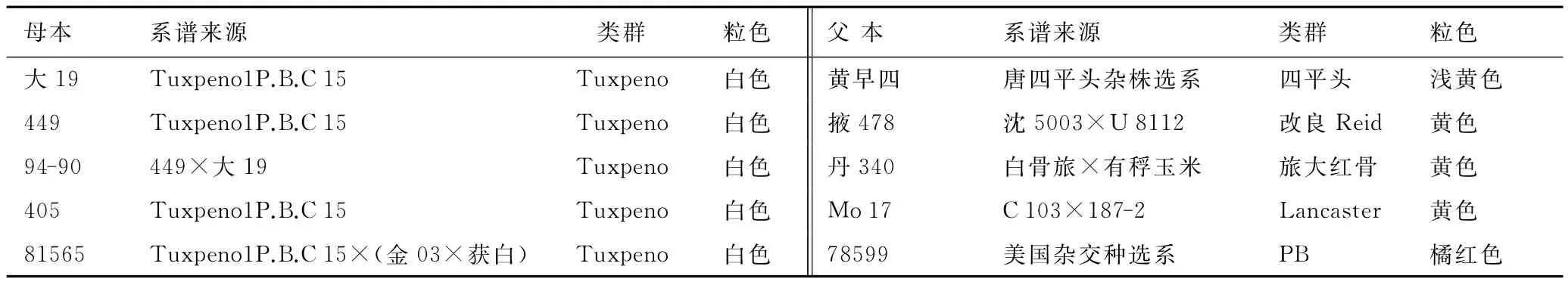
表1 供试材料名称、来源、类群和粒色

表2 25个杂交组合主要农艺性状方差及配合力方差分析
2.1.2 两点联合小区产量GCA和SCA效应分析
将两点试验结果汇总,10份供试材料在两种生态环境的产量一般配合力(GCA)相对效应值和产量特殊配合力(SCA)相对效应值(表3)。从表3可以看出,Tuxpeno种质中,大19和81565一般配合力(GCA)相对效应值呈正向效应,其值分别是1.55和2.47。这2个自交系较其他Tuxpeno种质选系增产潜力大,有效和定向地应用到育种实践中,可以有效地提高贵州玉米的产量。449、405和94-90为负效应,分别为-0.50、-2.82和-0.69。另外任意2个 Tuxpeno种质选系的GCA相对效应值之差都大于差数标准误(0.074),说明Tuxpeno种质的选系在小区产量性状上差异较明显。在五大种质骨干系中,一般配合力(GCA)相对效应值表现正效应的是黄早四(g=0.66)和掖478(g=2.52),一般配合力表现负效应的是丹340(g=-1.32),Mo 17(g=-1.77)和78599(g=-1.09)。黄改系(黄早四)×改良 Reid(掖478)是我国东、华北和黄淮海温带亚温带地区主要的杂优模式,黄早四小区产量的一般配合力(GCA)相对效应值较高,掖478的小区产量一般配合力(GCA)相对效应值也高,它们组配的杂交种丰产性一般比较好,郑单958,浚单18,浚单20,济单7等一大批优良杂交种大面积的推广正好吻合此观点。说明它们的产量性状主要受加性基因效应,其次是加性×环境的综合控制的作用。另外,在测验种中,任意2个测验系间的一般配合力(GCA)相对效应值之差均大于差数标准误(0.074);进一步说明,测验系之间在小区产量性状上的差异明显。
从两点联合小区产量一般配合力和特殊配合力相对效应值(表3)可以看出,小区产量的特殊配合力(SCA)相对效应值为正的有13个组合,有12个组合为负值;在25个组合中,5个Tuxpeno种质选系的特殊配合力(SCA)相对效应值较高的组合,其中94-90和 Mo 17的特殊配合力(SCA)相对效应值为6.86,在25个组合中最高,这与贵州主要的杂优模式(Tuxpeno种质×Lancaster)相吻合。其次是81565×78599的SCA相对效应值为6.82,依次是大19×黄早四、405×黄早四和449×掖478等,其SCA相对效应值分别是3.06、2.70和2.48;同时,特殊配合力(SCA)相对效应值较低的组合是81565×Mo 17(s=-5.29),449×78599(s=-5.35),94-90×黄早四(s=-4.36)和81565×黄早四(s=-2.86)。表明Tuxpeno种质与测验种之间具有较强的非加性基因效应,容易受到环境影响,产量容易产生较强的杂优组合。

表3 两点联合小区产量GCA和SCA相对效应值
2.2 利用SSR分子标记分析10个供试材料的遗传关系
2.2.1 SSR检测
55对SSR引物中筛选出多态性较高且带型稳定、清晰的25对SSR引物,在10个供试材料中共检测出127个等位基因变异(表4),每对引物检测出2~10个等位基因,平均每个位点检测到的等位基因数为5.08个,高于袁力行[11]利用66对SSR引物分析29个骨干自交系;Senior等[12]利用70对SSR引物分析94个骨干自交系(5.0);以上分析可以发现,待测样本的大小以及样本个体间的变异是造成这种差异的主要原因之一。
每个位点的多态性信息量(PIC)变化为0.20~0.90,其中引物umc 1065位点的PIC最大为0.90,引物umc 2163最小为0.20,平均为0.58;平均 PIC与Senior等[15](0.62)的结果相似,而高于李新海等[8](0.52),袁力行[11](0.54)的结果,这可能与供试材料样本大小以及选育引物种类和数量的不同有关。
2.2.2 SSR聚类分析
从表4和图1可以看出,10份自交系之间遗传距离范围是0.415 7~0.972 2;遗传距离最小者为 Mo 17和丹340,遗传距离为0.417 5;遗传距离最大者为94-90和丹340,遗传距离为0.972 2;根据遗传距离的数值矩阵,按UPGMA进行聚类分析,得到10份供试材料的聚类图(图1)。聚类结果表明,以遗传距离以0.70为标准,将10个供试材料划分为2个大类群:第一群包括:黄早四,78599,丹340,掖478和 Mo 17(五大测验种);第二群包括:大19,449,81565,94-90和405(Tuxpeno种质)遗传距离以0.53为标准划分,第一个大类群可以再划分成3个亚群,第一亚群包括黄早四和78599;第二亚群以掖478独立为一群;第三亚群包括丹340和Mo 17。第二大类群也划分成3个亚群,第一亚群包括大19和449;第二亚群包括81565和94-90;第三亚群以405独为一群。
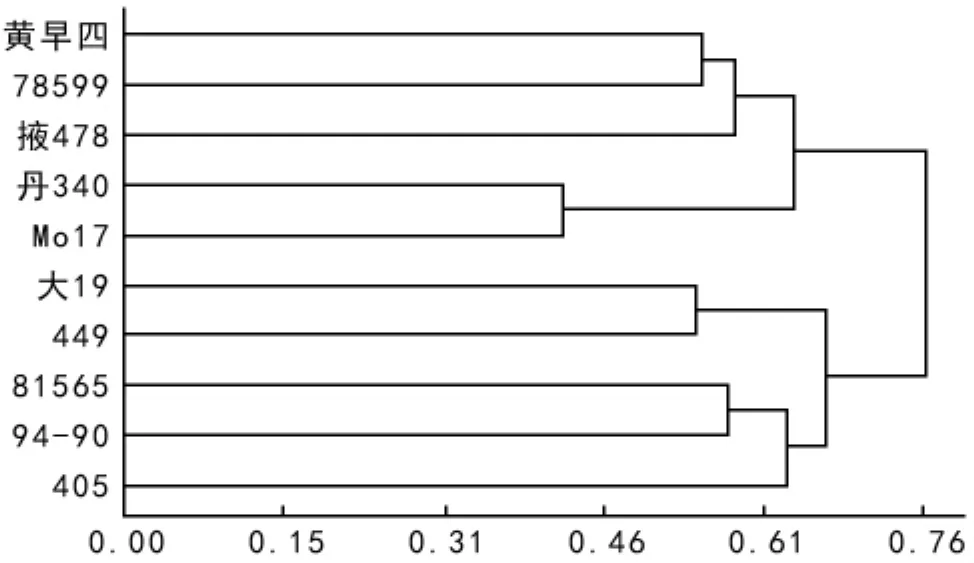
图1 SSR遗传距离对10个供试材料的聚类图
2.2.3 SSR遗传距离分析
从10个供试材料之间的遗传距离变化范围在0.415 7(掖478和丹340)至0.972 2(94-90和丹340)(见表5)。本研究结果得出:Tuxpeno种质之间的平均遗传距离为0.637 1,小于25个组合之间的平均遗传距离0.690 7;说明了供试的Tuxpeno种质之间差异较小;五大测验系之间的平均遗传距离为0.593;说明Tuxpeno种质内之间差异不明显。Tuxpeno种质和测验种之间的平均遗传距离为0.76。说明Tuxpeno种质与测验系之间的遗传距离差异明显。黄早四与Tuxpeno种质之间的平均遗传距离为0.766 3,与所有材料间的平均遗传距离为0.689 0;说明黄早四与Tuxpeno种质之间的遗传关系较远;与小区产量聚类结果一致,按地理来源划分也符合,唐四平头群是我国宝贵的地方种质资源,在贵州山地生态区利用,要注意保持其黄早四的高配合力,提高出籽率和抗性为选育目标;然后,构建Tuxpeno种质×黄改系杂种优势模式,这需要今后长期的育种实践来验证和探索。Mo 17与 Tuxpeno种质之间的平均遗传距离为0.758 0,与所有材料间的平均遗传距离为0.683 4;Mo 17与Tuxpeno种质的遗传距离较远,SSR分子标记结果与贵州省玉米杂种优势模式(Lancaster×Tuxpeno种质)相一致。丹340与Tuxpeno种质之间的平均遗传距离为0.832 4,与所有材料间的平均遗传距离为0.740 4;掖478与Tuxpeno种质之间的平均遗传距离为0.731 1,与所有材料间的平均遗传距离为0.682 1;从五个测验种与Tuxpeno种质之间的平均遗传距离作比较来看,丹340与Tuxpeno种质之间的遗传关系较远,表明Tuxpeno种质与丹340之间可能存在较强的杂种优势的遗传基础;按地理来源看,丹340是我国玉米温带区域重要的地方种质资源,Tuxpeno种质与丹340生态地理差异较远,经过长期的生态驯化,它们之间的遗传差异较大,所以,杂种优势比较明显。
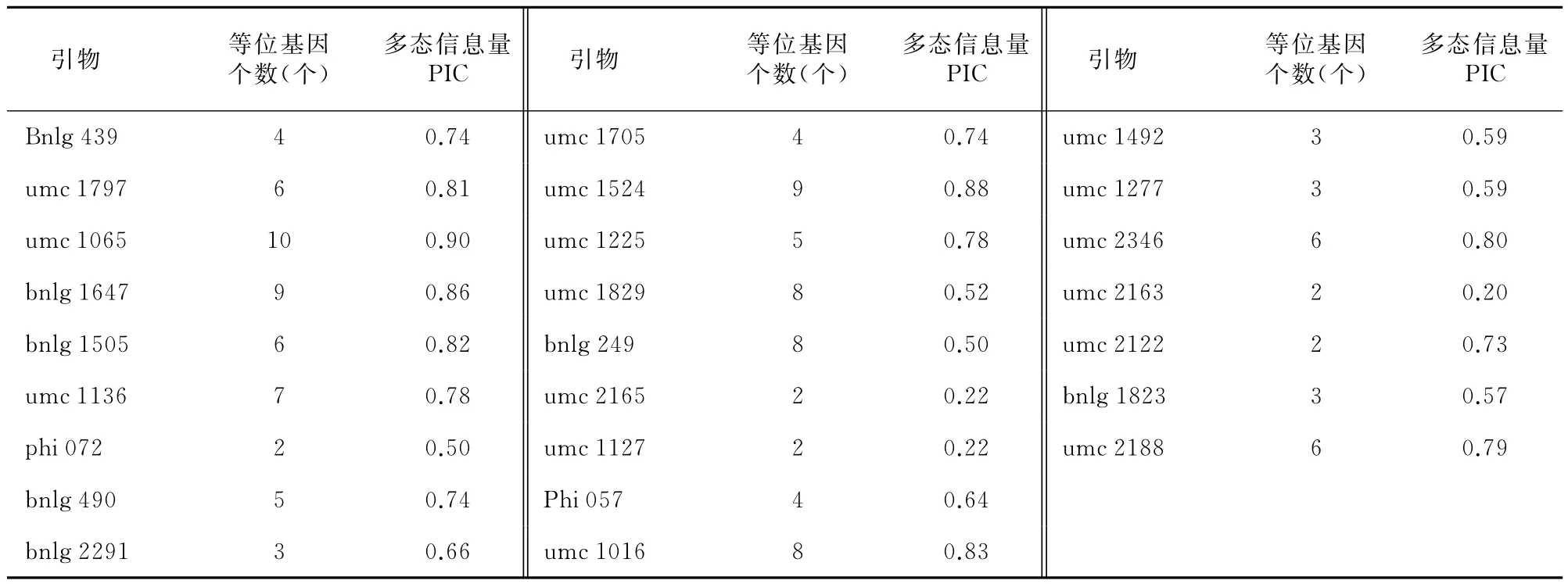
表4 25对SSR引物、等位基因数及多态信息量

表5 10个供试材料之间的遗传距离
2.3 玉米Tuxpeno种质自交系的产量及比对照优势
以玉米杂交种黔单16号为对照种,以各组合的小区产量均值为基础;分别计算各组合对照优势H(%);从表6可以看出,25个组合的平均对照优势为0.00%。其中有3个组合产量对照优势为正值;在25个杂交组合中,组合(大19×掖478)超对照种优势最强,其值为12.33%;组合(81565×掖478)超对照种优势较强,其值为7.87%;组合(81565×78599)超对照种优势较弱,其值为2.76%。

表6 玉米Tuxpeno种质杂交组合的产量及比对照优势
3 结论与讨论
3.1 田间NCⅡ杂交分析与SSR遗传距离分析结果比较
通过本研究对小区产量聚类和SSR遗传距离的比较,两者存在明显的不一致性,为了进一步探讨分子标记遗传距离和产量两者的关系,进行简单相关分析;分子标记遗传距离与两点联合产量的相关系数为0.07,说明不能单纯用SSR分子标记所计算的遗传距离对产量进行预测。
李明顺等[7]利用SSR聚类与产量的两种聚类结果差异比较大,进一步说明,不能简单地用分子标记遗传距离来预测产量;要对产量进行预测,忽视双亲GCA的作用,这是造成SCA与杂交种产量相关程度不高的一个重要原因,但对遗传距离与杂交种产量的相关程度没多大关系。这说明小区产量划分遗传距离受到环境影响比较大。SSR分析以作物的DNA为研究对象,排除环境对性状的影响,能从本质上解释作物的遗传特性,由于SSR分析是以DNA中的简单重复序列为扩增对象,仅能反映DNA中的部分信息,并不能反映作物基因型的综合表现。因此,在实际育种工作中,应该采用小区产量和分子标记分析相结合的方法。
综合以上得出,通过数量遗传学和分子标记聚类,即小区产量和分子遗传距离,综合这些方法分析结果信息,不能简单的用一种模式束缚育种工作者创新思维的开发,要有创新思维和清晰的思路,同时与育种实践相结合,田间试验与生物技术相结合,以致于更好的改良材料和种质创新,探索出新的杂种优势模式。在改良和利用Tuxpeno种质时,我们必须建立正确的育种技术核心。这样做还要考虑一个因素,那就是Tuxpeno本身的遗传基础很复杂,我们得到的Tuxpeno可能与别人的Tuxpeno有很大区别。这就好像有许多不同的Reid一样。SS就在A列,而Reid-Iodent就被美国人推到NSS那一边去了[10]。首先,我们要全面灵活掌握Tuxpeno种质材料的特征特性,采用田间试验和分子标记相结合,准确的把Tuxpeno种质选系或改良性归于不同的类群,以致于更好的为玉米育种和生产所利用。
3.2 热带玉米Tuxpeno种质的利用探讨
玉米种质匮乏和生产上所用杂交种遗传基础狭窄已经成为制约我国玉米育种和生产发展的重要因素[10]。为了拓展我国玉米种质基础,近年来许多育种者在引进热带种质上做了大量的工作,并取得了一定的进展。热带亚热带与温带玉米种质遗传差异较大,形成的杂种优势潜力较大。Tuxpeno种族是墨西哥纯马齿型玉米,种质组成复杂,包括白粒和黄粒,配合力好,坚秆,对小斑病有抗性,缺点是晚熟,株高和穗位高都偏高,根系不发达,感甘蔗花叶病。谢传晓等[2]利用SSR分子标记技术把Reid和Tuxpeno划分到一个杂种优势列,从理论上是正确的,但实践中还少见报道过把Tuxpeno种质导入Reid群,然后选系成功的例子;本研究综合分析发现,Tuxpeno种质群体内遗传比较丰富,通过热导温的方法,可以有效地改良我国温带材料,提高温带系的抗逆性,选育出的二环系按原来杂优模式组配新的组合,最终实现热带种质的合理有效利用;此外,在本研究中,根据产量鉴定结果和SSR综合分析,Tuxpeno种质与78599和Reid种质可能组配成较大潜力的杂种优势模式。Tuxpeno种质与Reid类群的配合力较强,与Lan.类群的配合力总效应比较弱;原因一是Tuxpeno种质遗传基础复杂,遗传多样性丰富;二是所选的Tuxpeno种质选系材料较少,三是鉴定地点少;由此应该进一步对Tuxpeno种质进行深入的研究,合理评价和利用Tuxpeno种质资源,以至于拓宽我国狭窄的玉米种质基础。
[1]张志国.Tuxpeno种质在玉米育种中的利用[J].作物杂志,1994(3):20-21.
[2]谢传晓,张世煌,李明顺.187份中国玉米自交系的Q-matrix数据―数值化系谱[J].遗传学报,2007,34(8):738-747.
[3]陈泽辉,祝云芳,王安贵,等.玉米Tuxpeno和Suwan种质的改良研究[J].贵州农业科学,2010,38(2):1-4.
[4]陈泽辉.群体和数量遗传学[M].贵州:贵州科技出版社,2009.
[5]郭平仲编著.数量遗传分析[M].北京:北京师范学院出版社,1987.
[6]孔繁玲.植物数量遗传学[M].北京:中国农业大学出版社,2006.
[7]李明顺,张世煌,李新海,等.根据产量特殊配合力分析玉米自交系杂种优势群[J].中国农业科学,2002,35(6):600-605.
[8]李新海,袁力行,李晓辉,等.利用SSR标记划分70份我国玉米自交系的杂种优势群[J].中国农业科学,2003,36(6):622-627.
[9]袁力行,傅骏骅,张世煌,等.利用RFLP和SSR标记划分玉米自交系杂种优势群的研究[J].作物学报,2001,27(2):149-156.
[10]张世煌.玉米种质创新和商业育种策略[J].玉米科学,2006,14(4):1-3,6.
[11]袁力行,傅骏骅,张世煌,等.利用RFLP和SSR标记划分玉米系杂种优势群的研究[J].作物学报,2001,27(2):149-156.
[12]Hallauer A R.,J.B.Miranda,Quantitative Genetics in Maize Breeding,Iowa State University Press,1981,231-233.
[13]Hallauer A R.Introgression of Elite Subtropical and Tropical Germplasm with U.S.Corn Belt Germplasm[C].2003:743-746.
[14]Well hausen E.J.,Improving American Corn with exotic germplasm,Annu.Hybrid Corn Ind.Res.Conf.proc.,1956,11:85-86.
[15]Senior M L,Murphy J P,Good man M M,et al.Utility of SSRs for determining genetic similarities and relationships in maize using an agarosege system [J].Crop Sci,1998(38):1 088-1 098.
