舟山近海日本囊对虾(Marsupenaeus japonicus)野生群体与露天养殖越冬群体间的形质差异*
2016-01-15李鸿鹏任夙艺王志铮林祖好
富 裕 李鸿鹏 任夙艺 王志铮① 林祖好
(1. 浙江海洋大学 舟山 316022; 2. 岱山沙城养殖场 岱山 316200)
日本囊对虾(Marsupenaeus japonicus)既是我国传统海水虾类养殖对象, 也是我国活虾出口创汇的重要养殖品种。舟山系我国重要的海水产品生产、加工、销售基地之一, 承担着沿海江苏、浙江、上海、福建, 和山东四省一市在东海区生产的水产品集散功能。受地理纬度限制, 分布于舟山近海的日本囊对虾具向深水区越冬洄游的习性(毛锡林等, 1994), 而养殖于舟山沿海露天养殖池塘内的日本囊对虾秋苗,因在越冬养成过程中需经受持续低温和高pH值的双重胁迫(富裕等, 2014), 致使其养殖成活率显著低于春季和夏季养殖的人工苗种。因此, 针对目前舟山市场冬、春季日本囊对虾活体供应严重短缺, 且销售价格异常高昂之实际, 深入开展适合于舟山沿海养殖的日本囊对虾秋苗的优质亲体选种工作, 无疑对促进舟山乃至东海区日本囊对虾养殖产业的健康发展具重要现实意义。
近年来, 有关日本囊对虾种质地理学的研究已日益引起国内外学者的关注和重视。蔡晓鹏等(2010)定量比较了捕捞自我国福建厦门、海南陵水、广西北海、广东惠来和福建诏安5个地理群体间的形态差异;李义军等(2010)对取自福建厦门、广东湛江和海南临高的野生群体以及养成于海南而种苗源自福建厦门的人工养成群体进行了形态差异与判别分析的研究;Tsio等(2005, 2007)根据头胸甲侧面斜纹的延伸特征将日本囊对虾分型为主布于东亚海域的形态变异Ⅰ型(varietyⅠ, 斜纹延伸至头部底缘)和主布于南海海域的形态变异Ⅱ型(varietyⅡ, 斜纹仅延伸至头胸甲侧面中央), 就上述分型, 何永琴等(2012)采用微卫星标记法比较了两形态变异类型群体间的遗传多样性,宋晓红等(2014)开展了两形态变异类型稚虾间的耐高温性能差异研究, 董宏标等(2014a, b)比较研究了在室内同池饲养条件下两形态变异类型人工养成群体间生长特征及主要外形参数的差异。但迄今国内外尚未见有关日本囊对虾夏季海捕野生群体与经冬季持续低温和高pH值双重胁迫存活下来的秋苗越冬养成群体间形质差异的研究报道。鉴于此, 本文作者拟采取聚类分析、主成分分析和判别分析方法, 定量比较舟山近海日本囊对虾海捕野生群体与露天越冬养成群体间的形质差异, 旨为舟山近海日本囊对虾秋苗优质野生亲体的筛选研究提供基础资料。
1 材料与方法
1.1 实验样品
本研究所用野生日本囊对虾捕自舟山市岱山县长涂岛邻近海域, 系2014年8月11日虾拖网作业渔获的活虾; 所用露天池塘越冬养成日本囊对虾取自长涂镇沙城养殖场, 为 2013年 9月 27日放苗养殖,2014年4月29日起捕的同池养成的活虾。于上述两类活虾中各随机选取无病无伤、附肢完整、反应灵敏的健壮个体 150尾组成本研究所涉日本囊对虾野生测定群体(以下简称“野生群”)和日本囊对虾露天池塘越冬养成测定群体(以下简称“越冬群”)。
1.2 生物学指标的测定及形质评价指标的统计
用数显游标卡尺(精度0.02 mm)和RS-232型精密电子天平(精度0.01g)逐尾进行生物学指标测定后,以野生群和越冬群实验样本间形态表型及以体长为基准的体型特征差异为基础, 分别统计归属于形态分类特征性状的X1(额剑上缘锯齿数), 归属于形态尺寸比例指数的 X2(眼窝距/体长×100%)、X3(眼窝距/头胸甲长×100%)、X4(眼窝距/额剑长×100%)、X5(眼窝距/眼径×100%)、X6(尾节宽/体长×100%)、X7(尾节宽/胸宽×100%)、X8(尾节宽/第二腹节宽×100%)、X9(尾节高/尾节宽×100%)、X10(胸宽/体长×100%), 归属于体横截面比例指数的X11(胸部横截面指数)、X12(第二腹节横截面指数)、X13(尾节横截面指数), 以及归属于营养生理状况指数的 X14(鲜出肉率)、X15(肥满度Ⅰ=体重/体长3×100)(Fulton, 1911)、X16(肥满度指数Ⅱ=肉重/体长3×100)、X17(重长指数Ⅰ=体重/体长)(房继明等, 1995)、X18(重长指数Ⅱ=肉重/体长)等18项形质评价指标。
1.3 数据处理
采用SPSS 19.0软件统计本研究所涉日本囊对虾野生群和越冬群各形质评价指标的均值、标准差和变异系数; 根据上述各项形质评价指标的均值, 计算野生群和越冬群间经标准化处理的欧氏距离(全距为1.00); 根据所有日本囊对虾实验样本的形质评价指标值进行主成分分析和判别分析, 其中主成分分析以特征值大于 1且累计贡献率大于 80%为原则确定主成分(PC)的提取个数, 采用逐步导入剔除法进行判别分析, 并计算判别准确率(P1为某判别正确的个数占该群体实际个数的百分比, P2为某判别正确的个数占判入该群体个数的百分比)和综合判别率(P为两实验对象判别正确的个数之和占实验对象总个数的百分比)(Li et al, 2005)来验证所建判别方程组的可靠性。
2 结果
2.1 日本囊对虾野生群与越冬群间形质评价指标值的比较
由表1可见, 日本囊对虾越冬群与野生群个体间的形质特征差异主要表露为: (1) 从分类特征性状上看, 越冬群个体 X1显著小于野生群(P<0.05); (2) 从尺寸比例指数上看, 越冬群个体 X2、X3、X5和 X9均显著小于野生群(P<0.05), X4、X6、X8、X10均显著大于野生群(P<0.05), 两者仅 X7无显著差异(P>0.05),表明越冬群较野生群个体凸显出胸部较宽而眼窝距和额剑均较短的体型比例特征; (3) 从体横截面指数上看, 越冬群个体 X11和 X13均显著大于野生群(P<0.05), X12与野生群无显著差异(P>0.05), 表明越冬群个体的胸部和尾部较野生群更显壮硕; (4)从营养生理状况指数上看, 越冬群所涉各项形质评价指标均显著大于野生群(P<0.05), 表明越冬群较野生群个体具更强的增重和产肉性能。综上可知, 日本囊对虾野生群与越冬群在本研究所涉18项形质评价指标中的相似性指数仅为 11.11%, 即两者为在形质特征上具较好区分度的两个群体。
2.2 日本囊对虾野生群与越冬群间形质特征的主成分分析
经Bartlett球形检验和KMO适合度检验, 发现形质评价指数相关系数矩阵与单位阵有显著差异(P<0.05)且适合度尚可(KMO=0.708>0.700), 表明本研究所涉形质评价指标适合做主成分分析。由表2可
见, 所列5个主成分的特征值均大于1且方差累计贡献率达85.270%, 故可认定它们为能概括本研究所涉日本囊对虾越冬群和野生群实验样本间形质特征差异的公共因子。其中, PC1的方差贡献率最大(35.137%)且远高于其它主成分。将载荷绝对值P>0.5的变量确定为主要影响变量, 则 PC1、PC2、PC3、PC4主要变量影响力绝对值的排序依次为X17>X1>X16>X18>X2>X14>X15>X12>X6>X7、X3>X4>X5>X8>X11>X10>X12>X6、X8>X5>X7>X9、X9>X10>X13, PC5主要变量仅为X13。由此可见, 公共因子方差贡献率的大小与其所包含的主要影响变量个数的多寡密切相关, 其中PC1不仅主要影响变量个数最多, 且所有载荷绝对值达到 0.80以上的主要影响变量也均仅存在于PC1中, 无疑其在对本研究所涉日本囊对虾越冬群和野生群实验样本间形质特征的区分上较其它主成分更具重要性。绘制PC1与其它各主成分间的得分散布图, 得图1。由图1可见, 虽然越冬群和野生群个体沿FAC1轴自左向右分别占据各自独立的区域, 但两者在该得分轴[–0.5,0]区间却存在较大程度的重叠, 表明仅通过 PC1尚未能精准地反映本研究所涉日本囊对虾全部实验个体的类群归属问题。

表1 日本囊对虾野生群和越冬群实验样本形质评价指标的参数统计Tab.1 The parameter statistics of morphological evaluation indices for all tested groups

表2 日本囊对虾野生群和越冬群实验样本形质评价指标的主成分分析Tab.2 The principal component analysis of morphological evaluation indices for all tested groups

图1 日本囊对虾野生群和越冬群实验样本形质评价指标的主成分散布图Fig.1 The scatter diagram of principal component of morphological evaluation indices for all tested groups
2.3 日本囊对虾野生群与越冬群间形质特征的判别分析
采用逐步导入剔除法, 从表1所列形质特征变量中筛选出对判别贡献较大的X1、X2、X4、X5、X10、X11、X12、X16和X17等9个变量进行判别分析,F检验表明这些变量均达到极显著水平(P<0.01)。根据上述9个变量建立本研究所涉日本囊对虾野生群和越冬群实验样本的Fisher分类函数方程组于表3。经验证, 越冬群实验样本的判别准确率P1、P2分别为 96%和99.31%, 野生群实验样本的判别准确率P1、P2分别为99.33%和96.13%, 综合判别准确率为96.67%。另, 所绘制的典型判别函数判别得分散布图(图 2), 更直观地印证了上述判别结果的可靠性。

表3 日本囊对虾越冬群和野生群形质特征Fisher分类函数方程组的自变量系数及常数项Tab.3 The independent variable coefficient and constant terms of the Fisher classification functions of morphological traits for all tested groups
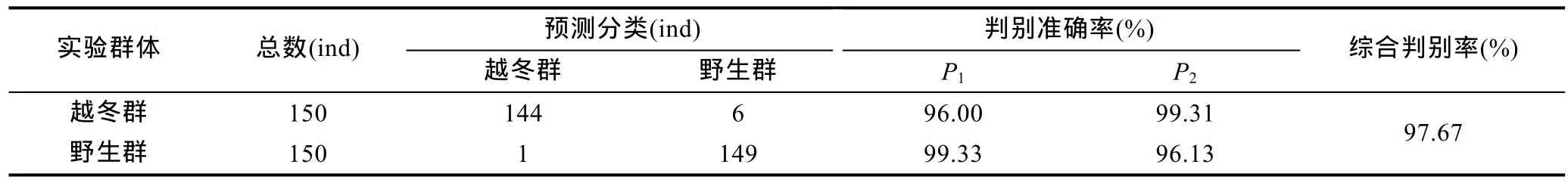
表4 日本囊对虾野生群和越冬群实验样本的判别分类结果Tab.4 The discrimination classification for all the tested groups

图2 日本囊对虾野生群和越冬群实验样本的典型判别函数判别得分散布图Fig.2 The scatter diagram of the typical discriminant function for all the tested groups
3 讨论
3.1 日本囊对虾越冬群与野生群间的形质特征差异及关键判别性状组合的认定
本研究所涉日本囊对虾越冬群与野生群实验样本在表1所列的 18项形质评价指标中, 具显著差异的多达16项(表1), 且两者形质评价指标均值间的标准化欧氏距离也具显著差异(表 2)的情形, 表明两者在形质特征上具较好的区分度。但上述存在显著差异的形质评价指标并非均为区分本研究所涉日本囊对虾越冬群与野生群间形质特征的关键判别变量, 而要找到真正可作为判别两者形质特征差异的关键性评价指标组合尚需借助主成分分析和判别分析才能得以实现。
由表2及图1可见, PC1的贡献率均明显大于其它各主成分, 但仅借助PC1尚无法准确判定本研究所涉日本囊对虾位于该得分轴[–0.5, 0]区间内个体的类群归属问题。由表3及图2可见, 从属于PC1主要影响变量的 X1、X2、X12、X16、X17和从属于 PC2主要影响变量的X4、X5、X10、X11等9个变量共同构成了判别本研究实验对象类群归属问题的关键变量组合。造成上述情形的原因, 无疑既与 PC1贡献率(35.137%)仅占累计贡献率(85.270%)的 41.206%, 致使其所含主要影响变量组合尚未能达到可全貌性地概括本研究所涉日本囊对虾越冬群与野生群个体间的形质特征差异有关, 也与PC1和PC2贡献率加和值(54.704%)占累计贡献率的64.154%, 致使两者所含主要影响变量在全貌性概括越冬群与野生群个体间形质特征差异上实现了信息的有效互补有关。而至于表2中各主成分其它主要影响变量未能被选入关键判别变量组合的原因, 本文作者认为是由于它们和表3中部分关键判别变量间存在信息重叠所致, 即: (1) 由越冬群X2、X3均显著小于野生群(P<0.05), X4、X6、X10、X11和X13均显著大于野生群(P<0.05), 以及X12与野生群无显著差异(P>0.05)(表1)所表露的, X3与X2之间、X6与X10之间以及X13与X11之间均存在一定程度的信息重叠的情形, 是导致与X2、X4同属眼窝距与虾体长轴上特征尺寸性状间比例关系的体型指标X3、与X11、X12同属虾体横截面指数的X13、以及与X10同属虾体纵向尺寸性状与体长之间比例关系的体型指标 X6均未被选入关键判别变量的主因; (2) X5、X7、X8和X9均为表征虾体纵向尺寸性状间比例关系的体型指标。由越冬群 X5、X9均显著小于野生群(P<0.05), X7、X8均显著大于野生群(P<0.05)(表 1)所表露的, X5与 X9之间、X7与 X8之间均存在一定程度的信息重叠, 以及X8与X9间的乘积与X7密切相关的情形, 均是导致X7、X8和X9未能被选入关键判别变量的原因; (3) X14、X15和 X18均未能入选表征营养生理状况关键判别变量的原因, 既缘于三者在 PC1中的载荷值均小于 X16和 X17, 更为重要的是它们均与 X16—X17组合间存在信息重叠有关。综上所述, 可将上述X1、X2、X4、X5、X10、X11、X12、X16、X17等 9个相互之间无信息重叠的变量认定为判别本研究所涉日本囊对虾越冬群与野生群实验样本形质差异的关键变量组合。
3.2 关于日本囊对虾越冬群与野生群间形质特征差异的原因分析
由表 2 可见, 以 X1、X2、X6、X12、X14、X15、X16、X17和 X18为主要影响变量的 PC1系以营养生理状况指数、额剑上缘锯齿数以及腹部肌肉发达程度为主导的,并与尾节和眼窝距宽窄程度相维系的体质强壮程度因子, 以 X3、X4、X5、X6、X8、X10、X11和 X12为主要影响变量的PC2是以寻食视野和胸腔空间大小为主导的, 并与腹节强壮程度和尾节宽窄程度相维系的捕食运动功能因子, Backhaus等(2000)指出主成分越靠后误差就越大, 故将以 X5、X7、X8、X9、X10、X13为主要影响变量的 PC3、PC4和 PC5统归为以尾节发达程度主导的, 并与胸腔大小和视距远近相维系的避敌运动功能因子。上述 5个公共因子所涵盖的体现本研究所涉日本囊对虾实验对象组间差异的体质强壮程度与捕食、避敌运动能力等一系列特征信息, 揭示了机体能量分配策略及其对应的捕食与避敌对策是引起本研究所涉日本囊对虾越冬群与野生群实验样本间产生形质特征差异的主因,从而客观地反映了所处生存环境的严峻度差异在导致本研究所涉日本囊对虾实验群体形质特征出现明显偏离上所具的生态重要性。
Wagner等(2001)认为, 同一物种的不同群体因长期适应不同的生存环境而形成了各自特有的包括生长、发育、繁殖以及对环境因子的适应力等的生物学特性。表1所示的本研究所涉日本对虾越冬群与野生群实验样本间的形质特征差异, 也充分支持了上述机体能量分配策略及其对应的捕食与避敌对策为影响两者间形质特征差异主因的观点, 即: (1) Schulte-Hostedde 等(2005)指出, 动物个体的生理或营养状况与其体内储存的能量物质间, 以及体内存储的能量物质与体重间均常呈正相关。突出于虾类头胸甲前端的额剑, 既具防御敌害及在游泳中起平衡身体等作用(王安利等, 1993; 沈辉等, 2010), 也系表征虾类体质强壮程度和生长代谢旺盛程度的重要体征(王志铮等, 2012)。通径分析表明, 北部湾野生日本囊对虾的额剑上缘锯齿数与体重间的通径系数呈显著水平(P<0.05) (孙成波等, 2008), 本研究所涉日本囊对虾野生群的额剑长与体重之间, 以及额剑上缘锯齿数与肉重之间的通径系数均达到显著水平(P<0.05), 而越冬群个体的额剑长和额剑上缘锯齿数与体重和肉重间的通径系数则均未达到显著水平(P>0.05)(李鸿鹏等, 2015)。无疑, 额剑在表征日本囊对虾野生群个体的体质强壮程度和生长代谢旺盛程度上较越冬群个体更具重要性。本研究中, 野生群个体X1、X2、X3和X5均显著大于越冬群个体(P<0.05), 而越冬群个体X4及所有营养生理状况指数均显著大于野生群个体(P<0.05)的结果, 无疑揭示了越冬群个体因食物保障程度和寻食成功率明显高于野生群个体, 故其采取通过显著缩小眼径和缩短眼窝距以更易于定位周边食物来提高捕食效率, 通过显著减少额剑上缘锯齿数和缩短第二腹节、额剑的长度以弱化食物追捕体征实现节能降耗的生存策略, 从而促使其营养生理状况指数均显著大于野生群个体(P<0.05); (2) 对虾类具通过张开尾扇, 腹部迅速向前弯曲, 使身体向后上方突然跃起, 随即重新伸直腹部, 并展开步足与触角,使身体缓慢下沉的后跃御敌习性(堵南山, 1993), 即尾扇在对虾类遇敌急速逃逸上具重要作用(王克行,1997)。本研究中, 越冬群个体 X6、X7、X8以及 X13均显著大于野生群个体(P<0.05)的结果, 无疑揭示了高密度圈养于露天池塘的越冬群个体, 为减少种内自残机率而采取的通过显著增粗尾节以起到增强逃逸能力的避敌对策; (3) 日本囊对虾系具洄游习性的经济虾种, 其幼体、仔虾与成虾生活于不同海区(堵南山, 1993; 毛锡林等, 1994), 而养殖于冬季露天池塘的日本囊对虾则需耐受持续低温和高pH值的双重胁迫(富裕等, 2014)。甲壳动物的鳃具呼吸、排泄、渗透压调节及病害防御等功能(周双林等, 2001)。本研究中, 越冬群个体 X10、X11均显著大于野生群个体(P<0.05), 以及在 X12与野生群个体相近(P>0.05)情形下, X9显著小于野生群个体(P<0.05)的结果, 无疑揭示了越冬群个体为尽可能消除高严峻度养殖理化环境对其存活和生长带来的负面影响, 而采取的通过扩增可表征鳃室空间大小的胸腔以起到提高耐受逆境能力的生存对策。上述分析表明, 导致日本囊对虾野生群与越冬群个体间形质特征差异的本质, 既为它们对各自所处生境的表型适应(戈峰, 2008), 也是它们基于生境选择压力而就机体能量分配策略及其对应的捕食与避敌对策进行权衡与优化的结果。因此,日本囊对虾秋苗野生亲本形质特征的选择, 应以秋苗越冬养成群体为参照。
王安利, 母学全, 周世梅等, 1993. 中国对虾行为的观察. 海洋科学, (4): 16—18
王志铮, 杨 磊, 施建军等, 2012. 日本沼虾(Macrobrachium nipponensis)池养越冬抱卵亲虾表型性状对个体繁殖力的影响效应. 海洋与湖沼, 43(1): 166—173
王克行, 1997. 虾蟹增养殖学. 北京: 中国农业出版社, 8
戈 峰, 2008. 现代生态学(2版). 北京: 科学出版社, 63
毛锡林, 蒋文波, 1994. 舟山海域海洋生物志. 杭州: 浙江人民出版社, 114
孙成波, 邓先余, 李镇泉等, 2008. 北部湾野生日本囊对虾(Marsupenaeus japonicus)体重和形态性状的关系. 海洋与湖沼, 39(3): 263—268
李义军, 李 婷, 王 平等, 2010. 日本囊对虾(Marsupenaeus japonicus)3个野生种群和 1个养殖种群的形态差异与判别分析. 海洋与湖沼, 41(4): 500—504
李鸿鹏, 富 裕, 任夙艺等, 2015. 舟山近海日本囊对虾野生群体与越冬养成群体形态性状对体重和肉重影响的比较. 海洋与湖沼, 46(5): 1218—1227
何永琴, 苏永全, 毛 勇等, 2012. 用微卫星标记分析不同形态变异类型日本囊对虾的遗传多样性. 水产学报, 36(10):1520—1528
沈 辉, 万夕和, 许 璞等, 2010. 脊尾白虾的行为学观察研究. 海洋科学, 34(10): 53—56
宋晓红, 毛 勇, 董宏标等, 2014. 两种形态变异类型日本囊对虾稚虾高温耐受性的比较. 水产学报, 38(1): 84—90
周双林, 姜乃澄, 卢建平等, 2001. 甲壳动物渗透压调节的研究进展Ⅰ. 鳃的结构与功能及其影响因子. 东海海洋,19(1): 44—51
房继明, 孙儒泳, 刘志龙, 1995. 布氏田鼠肥满度分析和小型兽类肥满度指标 K 与 KWL(重长指标)的比较. 动物学报,41(2): 141—148
堵南山, 1993. 甲壳动物学-下册. 北京: 科学出版社, 742—763
董宏标, 苏永全, 毛 勇等, 2014a. 日本囊对虾2种形态变异类型群体形态性状对体质量的影响效果分析. 厦门大学学报(自然科学版), 53(2): 289—296
董宏标, 苏永全, 毛 勇等, 2014b. 室内环境下日本囊对虾 2种形态变异类型群体生长特性比较研究. 热带海洋学报,33(4): 51—60
富 裕, Fatou D, 陈汉春等, 2014. 密植浒苔对冬季露天池塘池底水温、酸碱度和溶解氧的影响效应. 浙江海洋学院学报(自然科学版), 33(5): 389—393, 413
蔡晓鹏, 游欣欣, 曾凡荣等, 2010. 中国沿海日本囊对虾 5个地理群体间形态差异比较分析. 中国水产科学, 17(3):478—486
Backhaus K, Erichson B, Plinke W et al, 2000. Multivariate Statistical Analysis. Berlin: Springer Berdelberg
Fulton T W, 1911. The Sovereignty of The Sea. London and Edinburgh: William Blackwood and Sons
Li S F, Wang C H, Cheng Q Q, 2005. Morphological variations and phylogenesis of four strains in Cyprinus carpio. 水产学报, 29(5): 606—611
Schulte-Hostedde A I, Zinner B, Millar J S et al, 2005.Restitution of mass-size residuals: validating body condition indices. Ecology, 86(1): 155—163
Tsio K H, Wang Z Y, Chu K H, 2005. Genetic divergence between two morphologically similar varieties of the kuruma shrimp Penaeus japonicus. Marine Biology, 147(2):367—379
Tsio K H, Wang Z Y, Chu K H, 2007. Molecular population structure of the kuruma shrimp Penaeus japonicus species complex in western Pacific. Marine Biology, 150(6):1345—1364
Wagner E J, Arndt R E, Brough M, 2001. Comparative tolerance of four stocks of cutthroat trout to extremes in temperature.Western North American Naturalist, 61(4): 434—444
