大叶藻种子萌发环境条件研究*
2016-01-15唐学玺闫文杰
王 明 郭 馨 唐学玺 臧 宇 闫文杰 周 斌①
(1. 中国海洋大学海洋生命学院 青岛 266003; 2. 厦门大学环境与生态学院 厦门 361102)
海草(Seagrasses)是一种生长于热带和温带浅海水域的单子叶种子植物, 能在海水中完成开花、结果和萌发等生理过程(Fourqurean et al, 2012)。海草床(Seagrass bed)生态系统具有一系列重要的服务功能价值, 成为全球海洋生物多样性保护的焦点(郑凤英等, 2013)。大叶藻(Zostera marina)是北半球分布最广泛的海草类群, 是重要的海洋环境健康指示生物(Krause-Jensen et al, 2005), 其单位面积叶片N含量与生物量的比值也是指示水体早期富养化的污染指标(Lee et al, 2004)。大叶藻在我国分布十分广泛, 主要分布于辽宁、河北、山东等省份的沿海水域(范航清, 2009), 是黄渤海海草分布区的优势种(郑凤英等,2013)。20世纪以来, 由于自然和人为因素的影响, 大叶藻海草床在世界范围内出现严重退化, 且最近几十年退化速率加快(Costello et al , 2011)。叶春江等(2002)的研究指出过去的 30—50年期间我国山东近岸大叶藻分布面积减少了80%以上。大叶藻海草床的退化消失将直接导致生物栖息生境丧失、生物多样性锐减以及生物地球化学过程失衡, 加剧近岸生态系统的不稳定性或脆弱性(王伟伟等, 2013), 因此大叶藻海草床的保护与修复备受关注(Orth et al, 2010;Delefosse et al, 2012; Li et al, 2013)。
海草床的修复方法主要有生境恢复法、成体移植法和种子法, 各有利弊(Orth et al, 2010; Reynolds et al, 2012; Li et al, 2013)。种子法因其成本低、维持种群遗传多样性、对供体草床干扰小等特点, 得到了重视与发展。目前, 海草种子的收集、保存、播种和幼苗发育被公认为是一种具有生态学意义的恢复实践(Kendrick et al, 2012; Reynolds et al, 2012)。虽然海草种子是一种具有高生态适应性和经济可行性的大尺度恢复材料, 但其产量不稳定, 具有明显的时空变化(Orth et al, 2006)。传统种子法将大叶藻种子直接投放到修复海域, 由于受到营养盐、温度、盐度、pH等环境因素的制约, 以及湍流扰动和沉积物破坏等因素的影响, 自然条件下大叶藻种子的萌发率非常低, 仅为5%—15%(Ke et al, 2006; Jarvis et al, 2008)。为了提高修复效果, 我们对种子法进行改进, 预先在实验室条件下进行种子萌发, 再将萌发的种子投放到修复海域。
自20世纪80年代起, 对海草种子萌发的关注在于大叶藻科、丝粉藻科和水鳖科。萌发实验测定的因素包括温度、盐度、基质类型、光照、营养盐、激素和埋藏深度等。研究显示盐度(Conacheret al, 1994)、温度(Hayet al, 2008; Jarviset al, 2008)、光照(Lalet al,1993)及溶解氧(Brenchleyet al, 1998)是影响海草种子萌发的关键因素, 而盐度通常被认为是最重要的单一因子(Orthet al, 2000)。之前的研究多集中在单一或常规环境条件对种子萌发的作用, 而自然条件下海草种子通常受多种因素的交互作用或某些非常规因素的影响。为了保证大叶藻种子在实验室条件下的萌发效果, 有必要深入研究影响种子萌发的环境因素,并寻找可以有效促进种子萌发的处理方式。基于上述目的, 本研究着眼于影响种子萌发的四个环境因素——温度、盐度、pH以及营养盐浓度, 在实验室可控条件下利用正交实验探索影响大叶藻种子萌发的最主要因素并确定较适宜的萌发条件, 并在此基础上比较了多种处理方式(不同光照、UV-B辐射、低温层积和划破种皮)对大叶藻种子萌发的作用。
1 材料与方法
1.1 大叶藻种子的采集及保存
2014年7—8月, 大叶藻种子直接采集于山东省荣成市俚岛湾(122.586°E, 37.246°N), 自然条件下生长成熟的大叶藻生殖枝。种子运抵实验室后, 用滤网处理种子去除杂质和贝类等。选用长椭圆形、黑褐色、坚硬饱满的种子, 在 25°C、盐度 30、自然光、连续充气条件下暂存两周至萌发实验开始。
1.2 种子活力测定
根据ISTA种子检验规程(ISTA, 1996), 采用四唑(TTC, 1.0%)染色法测定种子活力。随机选取50粒种子置于35°C温水中浸泡2d后去皮, 35°C条件下用四唑溶液染色 12h, 根据种胚的着色部位及程度, 按照国际种子检验标准鉴定种子活力。
1.3 种子萌发条件的正交实验
选择温度、盐度、pH以及N/P营养盐浓度四种环境单因素, 设计四因素三水平正交实验, N/P由NaNO3、NaH2PO4和 Na2HPO4提供(表 1)。实验分为9个处理组(表2), 每组种子数量为50粒。以90 mm培养皿作为培养容器, 置于光照培养箱内, 光周期为12:12, 日光灯照度为~3000lx, 培养方法为浸泡培养法(Hootsmanset al, 1987)。种子的实验室培养基质为采集于青岛汇泉湾近岸的灭菌过滤海水(121°C,20min), 通过蒸馏水稀释得到不同盐度水平, 每个培养皿海水用量为30mL, 每两天更换一次, 初始ph由稀盐酸调节。每日记录萌发种子的数量并计算累积萌发率。根据结果筛选出9个处理组中最佳的条件组合,用于后续实验。

表1 正交实验因素水平L9 (34)Tab.1 Factors and levels for L9 (34) orthogonal array design
1.4 优化条件下种子萌发的影响因素
同样采用浸泡培养法, 每个培养皿内加入 30mL培养液和50粒种子, 培养液的盐度、N/P浓度、初始pH及培养温度按正交实验法得出的最优条件设置, 每天观察、记录种子萌发情况, 每两天更换一次培养液。
1.4.1 光质类型及比例对种子萌发的影响 实验使用波长为635nm的红色灯管、波长为460nm的蓝色灯管以及白色灯管各3支, 实验中将两种颜色的灯管分别悬于培养箱两层水培层架的顶部, 另外开启培养箱自带光源, 光周期为 12:12, 照度~3000lx。设置白光灯(W), 红光(R), 蓝光(B), 红蓝组合光1(R:B=1:1), 红蓝组合光 2(R:B=1:2), 红蓝组合光3(R:B=2:1)和黑暗(D)共7个处理组, 每组设置3个平行。红蓝组合光比例由两种光质的照射时长决定。
1.4.2 UV-B辐射对种子萌发的影响 以UV-B辐射的等比强度设置 4个实验组, 每组设置 3个平行,光周期为 12:12, 光周期内每日替代照射 UV-B(~600lx)时长分别为0h, 2h, 4h, 8h, 其余时间置于光照培养箱内。
1.4.3 低温层积对种子萌发的影响 实验种子在萌发实验前首先转入4°C冰箱预处理10d之后取出,按照正交实验中的最佳条件组合进行萌发实验。以无低温层积预处理的种子为对照组, 每个处理设置3个平行。所有实验用种子置于光照培养箱内, 光暗周期为 12:12, 照度~3000lx。
1.4.4 划破种皮对种子萌发的影响 将种子外种皮割破 , 以解剖刀在种子的子叶端、沿纵纹划破种皮, 开口约占纵纹长度的 1/3(Taylor, 1957), 避免伤害胚。对照组则保留完整种, 每个处理设置3个平行。所有实验用种子置于光照培养箱内, 光周期为 12:12,照度~3000lx。
1.5 种子萌发的判定标准
大叶藻种子萌发的判断依据为种子的子叶(内含种胚) 端的种皮发生涨裂且弯曲的 U形棒状子叶发生伸直并突出破裂的种皮(Harrison, 1991)。
1.6 数据处理与分析
萌发率为各处理组中萌发种子粒数占总粒数的百分数, 平均萌发时间(MGT)为:

其中,Nn为观察到的种子开始萌发的天数,n为天数,Gn为第n天观察到的种子萌发数(Ranal, 2006)。
利用SPSS 16.0对数据进行极差分析、方差分析。显著性水平设定为α=0.05, 利用SigmaPlot 12.5作图。
2 结果与分析
2.1 种子活力测定
四唑染色结果显示保存的种子有活力的比例为(91±3.1)%。
2.2 种子萌发的最优环境条件
经过 8周的萌发实验获得各条件下种子的最终萌发率, 结果如表 2所示。由各因素极差大小可知,R盐度>R初始pH>R温度>RN/P浓度, 表明4种不同因素对大叶藻种子萌发的影响大小的次序为: 盐度>初始 pH>温度>N/P浓度。其中,R盐度(44.72)最大, 即盐度对种子萌发率的影响最显著, 因此盐度为主要因素且萌发率最大值出现在第1水平。依次类推, N/P浓度、初始 pH、温度因素的种子萌发率最大值分别出现在第3水平、第3水平和第1水平。因此, 在本实验范围内, 大叶藻种子萌发的较适宜条件为盐度12、N/P浓度 960/60µmol/L、初始pH 6.20和温度 10°C, 此时大叶藻种子萌发率最高。

表2 大叶藻种子萌发率正交实验结果Tab.2 Results of orthogonal test of Z. marina seed germination rate
2.3 光质类型及比例对种子萌发的影响
在优化后的浸泡培养条件下, 比较了不同的光照条对大叶藻种子萌发的影响。培养8周, 黑暗条件下大叶藻种子萌发率最大, 为(92±4.8)%, 而其他光照条件下种子萌发率无显著差异(P>0.05)(图1)。红光条件下种子萌发速度较快, 至第四周萌发基本停止;蓝光条件下种子萌发速度较慢, 但在实验过程中不断进行(图1)。通过多重比较的LSD方法统计检验黑暗条件与其他条件下种子的萌发率差异, 发现黑暗条件与其他条件间的大叶藻种子萌发率影响均存在显著差异(P<0.05, 表 3)。

图1 不同光质类型及比例下种子的萌发率Fig.1 Germination rate of Z. marina seeds in different types of lighting
2.4 UV-B辐射对种子萌发的影响
无UV-B辐射时大叶藻种子的萌发率较低, 在一定强度范围内(每日经UV-B ~600lx连续照射8h), 种子萌发率随着UV-B辐射时长而升高, 每日8 h时萌发率可达(93±5.2)%(图 2)。由单因素方差分析可知,不同UV-B辐射时长下大叶藻种子萌发率存在显著差异(P<0.05), 且经 UV-B辐射的处理组在第 8周仍有萌发趋势, 而对照组种子的萌发则趋于停止。LSD多重比较结果显示: 白光与8h和2h与8h间的种子萌发率均存在显著差异(P<0.05), 说明 UV-B辐射可以有效提高种子萌发率(表4)。

表3 不同光质类型及比例对大叶藻种子萌发率影响的多重比较Tab.3 Multiple comparisons on germination rate in different types of lighting
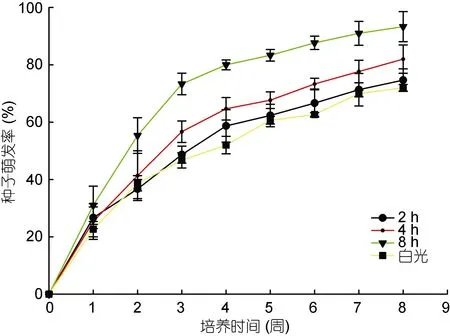
图2 UV-B辐射下藻种子的萌发率Fig.2 Rate of seed germination in different duration of daily UV-B radiation
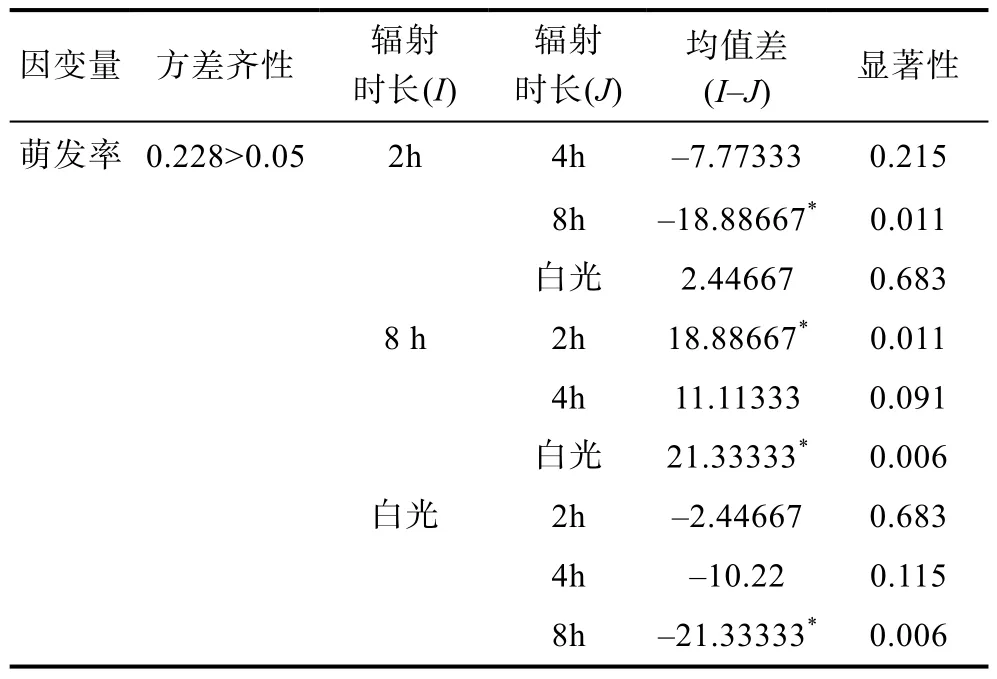
表4 每日UV-B辐射不同时长对大叶藻种子萌发率影响的多重比较Tab.4 Multiple comparisons on rate of germination in different duration of daily UV-B radiation
2.5 低温层积及划破种皮对种子萌发的影响
划破种皮处理组的大叶藻种子萌发率最高, 达(91±2.1)%, 且萌发速度快, 第2周就已达到萌发率最大值(图 3)。低温层积对种子萌发也有一定的促进作用, 萌发率在第 8周达到(83±3.6)%且仍有上升趋势(图 3)。单因素方差分析表明低温层积对种子萌发具有显著影响(P<0.05), 而划破种皮对种子萌发具有极显著影响(P<0.01)。

图3 低温层积及划破种皮时种子的萌发率Fig.3 Rate of seed germination under cold stratification and scarification of seed coat
2.6 不同条件对大叶藻种子平均萌发时间的影响
不同时长的UV-B辐射和不同类型及比例的光质对大叶藻种子的平均萌发时间基本无影响; 而划破种皮可以大大缩短种子平均萌发时间, 为(7.7±0.07)天; 低温层积预处理促进了种子萌发, 使得种子平均萌发时间缩短为(26.78±0.07)天(图 4)。
3 讨论
3.1 正交实验法确定种子萌发最优环境条件
在种间和种内对比的基础上, 通常认为降低盐度能够促进海草种子的萌发(Kahnet al, 2005; Kochet al, 2010; Fernández-Torquemadaet al, 2013), 原因是盐度降低可能干预淀粉的分解代谢导致种皮提早开裂(Sugiuraet al, 2009), 或者促进种子萌发前的水分吸收(Probertet al, 1999)。在恢复实践中常通过提高盐度来抑制存储种子的萌发(Ailstocket al, 2006)。本实验中 3个盐度范围均低于自然海区的正常盐度,之前的研究显示种子萌发率与盐度表现出极明显的相关性, 即随着盐度的升高种子萌发率显著降低。虽然盐度降低能够促进种子萌发, 但有研究表明低盐度会对大叶藻幼苗产生不利影响, 降低光合作用效率和生长速率(Tanner et al, 2010)。因此, 对大叶藻种子萌发条件的优化方案还应该结合后续的幼苗生长状况进行综合分析。

图4 不同条件下大叶藻种子的平均萌发时间Fig.4 Mean germination time (MGT) of seed under different light conditions
有学者认为温度是非休眠种子萌发最主要的影响因素(Probert et al, 1999)。尽管Hootsmans等(1987)和 Harrison(1991)指出大叶藻种子的萌发受到 10—30°C的高温促进(或不存在影响), 更多的观察结果表明高温能够抑制萌发。Abe等(2008)发现大叶藻种子萌发的最适温度为 10—15°C, 且最大萌发率出现在10°C。本实验选出的各处理组中最佳的条件中温度为10°C, 但种子萌发率并没有随着温度的升高而降低,表明温度对大叶藻种子萌发率的作用还受其他条件制约。也有研究认为大叶藻种子萌发的最适水温与当地的最低水温有关 (Orth et al, 1983; Pan et al, 2011);相反地 Loquès等(1990)的研究显示温度对诺氏大叶藻萌发的作用很小。基于上述研究, 我们认为海草种子的萌发可能存在一个与海草所处生境内温度的季节性变化密切相关的阈值, 在这个可能的阈值范围内, 种子萌发具有最适温度并存在种间差异。
目前对大型海藻及某些淡水水生植物的氮磷响应机制有较多研究, 而关于氮磷对大叶藻及其他海草影响的研究鲜有报道。有关研究发现, 氮是大型海藻生长的主要限制因子(Fong et al, 2004), 而在部分海域磷也是限制海藻生长的主要因子(郭卫东等,1998)。许多大型海藻都能够适应营养盐的变动, 在外界营养盐丰富时其最大吸收速率远远大于维持最大生长速率所需要的氮, 从而积累丰富的营养库以备营养盐不足(Mcglathery et al, 1996)。同样地, 程丽巍(2010)的研究中, 氮磷加富均能够显著促进龙须菜和石莼生长。本研究设置了 240/15、480/30以及 960/60µmol/L三个浓度, 各处理组中最佳的条件中氮、磷营养盐浓度为 960/60µmol/L, 但种子萌发率并没有随氮、磷浓度升高, 说明在本研究区域内氮、磷并非大叶藻种子萌发的限制性因素。
pH是影响植物种子萌发和幼苗生长的关键环境因子之一(Rivard, 1989; Arts et al, 1990), 随着酸雨增加及土壤盐碱化, 近年来有关 pH对陆生植物种子萌发的影响的研究已有大量报道。有研究表明, 在高pH条件下环境中易形成HCO3–等离子, 抑制种子萌发与幼苗生长 (马红媛等, 2007), 中性偏酸的条件则可以促进种子萌发(李焰焰等, 2005; 黄婷等, 2007; 何海洋等, 2013)。本实验的结果表明大叶藻种子具有较广泛的 pH适应范围, 不同 pH能够影响种子萌发, pH为6.20的条件更有利于种子萌发。
3.2 不同光质类型及比例对种子萌发的影响
本研究表明, 不同光质类型及比例对大叶藻种子萌发无明显影响。黑暗条件可以提高大叶藻种子的萌发率, 说明大叶藻种子萌发过程可能不需要光照,在无光条件下萌发更加显著。众多物理、化学和生物因素影响大叶藻存活、生长和繁殖并可能影响其群落动态, 光照是影响大叶藻生长和存活最重要的因素(Dennison, 1987; Olesen et al, 1993)。Pons等(1992)认为光照通过光敏色素的活动影响种子萌发, 而Karssen等(1990)认为导致萌发的过程能够在质膜蛋白与活性光敏色素结合时发生, 并得出光照能够促进多种植物种子萌发的结论。但对水生植物川蔓藻(Strazisar et al, 2013)和蓖齿眼子菜(高健等, 2006)的研究却发现光照并不能促进其种子的萌发, 这与本实验中黑暗条件可以提高大叶藻种子的萌发率的结论一致。
本研究发现光照条件虽然不利于提高种子的萌发率, 但在短期内采用红光照射却可在一定程度上加快种子萌发速度, 缩短平均萌发时间, 说明大叶藻种子萌发过程受到光质条件的影响。光质是影响陆生种子植物生长的重要因素。李雯琳等(2013)发现红光能够提高不同品种叶用莴苣种子的发芽活力, 蓝光则能够提高种子的活力指数。而在王维荣等(1991)的研究中红光和绿光对黄瓜种子萌发则表现出完全抑制效应。上述研究结果说明在萌发阶段, 不同种类的植物种子对不同光质的表现有显著差异, 可能是植物适应生境长期进化的结果。
大叶藻主要生长在潮间带和潮下带的浅海区域,与陆生高等植物的生态位具有很大差异, 因此现有的与陆生高等植物种子萌发的研究可能并不适用于海草类群。海洋环境与陆生环境差异明显: 例如红光和远红光在海水中衰减明显, 但陆生高等植物的研究却显示红光和远红光对种子萌发具有促进作用。虽然本研究结果表明蓝光与红光对大叶藻种子萌发率的影响无显著差异, 但是可以发现, 蓝光处理组的种子萌发作用较红光处理组更具有潜力, 在第8周实验结束时仍然有萌发上升趋势, 而红光处理组在第4周左右已达到萌发最大值。且蓝光处理组培养出的幼苗较红光处理组长势更好, 两种光质组合处理时蓝光比例越大幼苗生长状况越好, 红光比例大的处理组幼苗生长受到抑制。在海洋环境中, 光线在透过海水时, 红光被吸收最多而蓝光最少, 后者在海水中具有更强的穿透力。实验结果一定程度上支持了我们的推断, 即在大叶藻进化过程中, 可能形成了更适合海洋环境的萌发与生长策略, 因而对蓝光更加敏感, 能够更好地利用蓝光进行萌发和光合作用。
另外, 本研究结果中光质类型影响不显著的原因可能是受实验条件的设置与实验设备的限制, 比如优化后的条件组合可能对种子在萌发上对于光质的适应造成影响, 以至于一定程度上掩盖了其在自然海水条件下对于光质的适应。今后的研究中可进一步进行极端化处理, 增加单一光质光源照射强度和时间以排除其他光质的影响, 并应进一步探索大叶藻种子萌发对光质的内在适应机制(如光敏色素的成分及比例), 从根本上认识光质对种子萌发的影响。
3.3 UV-B辐射对种子萌发的影响
本研究结果表明, 一定强度的UV-B辐射有利于大叶藻种子萌发。可能的原因: 一是紫外线具有杀菌消毒作用, 能够抑制或杀死影响大叶藻种子萌发的微生物, 这些微生物可能会释放某些化学物质对种子产生不利影响, 也可能与大叶藻争夺生存空间、营养物质、氧气等资源; 二是一定强度的紫外线可以促进大叶藻种子的萌发。但这一结论并不具有一致性:针对陆生植物的研究表明紫外线辐射对种子萌发有抑制作用(胡正华等, 2005), 或对种子萌发影响不显著(方媛等, 2010)。而陈南凯等(1995)报道紫外灯照射种子能提高发芽力, 促进种子成熟且有明显的增产效果。郭彦等(2011)的研究结果同样表明照射10h和20h紫外线的水稻种子发芽率均高于对照组的不经过紫外线照射的种子。本实验结果显示, 在一定范围内随着UV-B照射时长增加, 大叶藻种子萌发率升高。实验结束时, 种子仍有明显的萌发趋势。本实验在一定程度上支持了紫外线辐射能促进种子萌发的观点,然而紫外线辐射促进大叶藻种子及其他种子萌发的作用机理还有待进一步研究。
3.4 划破种皮及低温层积预处理对种子萌发的影响
多数海草的种子具有坚硬种皮且具休眠期, 这有利于海草种群在高度动态性的生境中的长期存在(Orth et al, 2000)。但种皮的机械作用和休眠期使种子萌发受到限制。Loquès等 (1990)的研究证明了种皮在萌发启动上的重要作用, 认为种皮形成了一种可能阻止水分进入和/或降低胚与环境间的气体交换的屏障。Conacher等(1994)的研究表明低盐度下切割种皮能够促进摩羯大叶藻种子快速萌发, 可能是通过种子膨胀或突破外皮, 也可能通过促进水合作用和呼吸作用直接启动胚的发育。由文辉等(1995)对3种沉水植物(菹草, 大茨藻和苦草)种子萌发的研究表明,种皮破裂后发芽率和萌发速率均升高。陈开宁等(2005)研究沉水植物时也发现类似现象, 去除蓖齿眼子菜的种皮可将最终发芽率从6%以下提高到60%左右。本实验的结果同样表明划破种皮可以显著提高种子萌发率, 且大大缩短平均萌发时间。
本实验对大叶藻种子进行了10天的低温层积处理, 萌发率提高了19.07%, 平均萌发时间缩短了3.88天。国外学者开展了大量关于大叶藻种子休眠的研究,但所得结论并不一致。Churchill(1983)的研究表明大叶藻种子不存在休眠且低温(<15°C)是诱导萌发的主要因素。Orth等(1983)进一步的研究表明大叶藻种子不存在原初休眠而种子释放与萌发之间是由高温诱导的休眠期, 但 Hootsmans等(1987)在荷兰西南部的Zandkreek的研究并未发现该物种存在温度强迫性休眠的证据。法国胡安湾的诺氏大叶藻种群(Loquès et al, 1990)和结状丝粉藻种群也存在类似现象(Caye et al, 1992)。大叶藻种子的生理休眠可以在高温和低温层积时破除(Harrison, 1991), 处于打破休眠的理想条件下时种子的萌发时间更短(Kettenring et al, 2007;Vandelook et al, 2008)。低温层积处理时间的长短对种子萌发率的影响有待进一步研究。
4 结论
本实验优化得到大叶藻种子萌发的相对适宜为盐度12、N/P浓度960/60µmol/L、初始pH 6.20和温度 10°C。不同光质类型及比例对大叶藻种子萌发不具显著性作用, 但无光条件下的大叶藻种子具有较高的萌发率92%。UV-B辐射、划破种皮和低温层积处理均可显著提高大叶藻种子的萌发率并缩短平均萌发时间。
陈开宁, 李文朝, 潘继征, 2005. 不同处理对蓖齿眼子菜(Potamogeton pectinatus L.)种子萌发率的影响. 湖泊科学,17(3): 237—242
陈南凯, 王玉兰, 1995. 紫外线处理农作物种子能增产. 云南农业科技, (3): 15—16
程丽巍, 2010. 三种大型海藻对海水中营养盐供应变化的生理响应研究. 南京: 南京农业大学硕士学位论文
方 媛, 于海宁, 程 曦等, 2010. 增强 UV-B 辐射对甘草种子萌发及幼苗形态的影响. 中国农学通报, 26(2):122—126
高 健, 熊 勤, 李 祝等, 2006. 溶氧、pH及其他理化因子对蓖齿眼子菜种子萌发的影响. 水生生物学报, 30(5):524—529
范航清, 石雅君, 邱广龙, 2009. 中国海草植物. 北京: 海洋出版社
郭卫东, 章小明, 杨逸萍等, 1998. 中国近岸海域潜在性富营养化程度的评价. 台湾海峡, 17(1): 64—70
郭 彦, 杨洪双, 梁 明, 2011. 紫外线照射对水稻种子萌发的影响. 江苏农业科学, 39(3): 89—90
何海洋, 胡春芹, 丁强强等, 2013. 不同pH对光皮桦种子萌发及幼苗生长的影响. 西南林业大学学报, 33(5): 29—33, 39
胡正华, 索福喜, 赵晓莉等, 2005. UV-B辐射增加与酸雨复合处理对菠菜种子萌发和幼苗生长的影响. 中国农学通报,21(6): 284—285, 294
黄 婷, 董召荣, 宋 贺等, 2007. 模拟酸雨对紫花苜蓿种子萌发及幼苗生理特性的影响. 种子, 26(4): 21—24
李雯琳, 郁继华, 杨其长, 2013. 不同光质对叶用莴苣种子萌发和幼苗酶活性的影响. 甘肃农业大学学报, 48(3):44—49
李焰焰, 聂传朋, 董召荣, 2005. 模拟酸雨对小黑麦种子萌发及幼苗生理特性的影响. 生态学杂志, 24(4): 395—397
马红媛, 梁正伟, 2007. 不同 pH值土壤及其浸提液对羊草种子萌发和幼苗生长的影响. 植物学通报, 24(2): 181—188
王维荣, 陈 松, 欧阳光察等, 1991. 光质对黄瓜种子萌发过程中过氧化物酶活性及蛋白含量的影响. 上海农业学报,7(4): 17—20
王伟伟, 李晓捷, 潘金华等, 2013. 大叶藻资源动态及生态恢复面临的问题. 海洋环境科学, 32(2): 316—320
由文辉, 宋永昌, 1995. 淀山湖3种沉水植物的种子萌发生态.应用生态学报, 6(2): 196—200
叶春江, 赵可夫. 高等植物大叶藻研究进展及其对海洋沉水生活的适应. 植物学通报, 2002, 19(2): 184—193
郑凤英, 邱广龙, 范航清等, 2013. 中国海草的多样性、分布及保护. 生物多样性, 21(5): 517—526
Abe M, Kurashima A, Maegawa M, 2008. Temperature requirements for seed germination and seedling growth of Zostera marina from central Japan. Fisheries Science, 74(3):589—593
Ailstock S, Shafer D, 2006. Protocol for Large-scale Collection,Processing, and Storage of Seeds of Two Mesohaline Submerged Aquatic Plant Species. Vicksburg, MS, USA: US Army Engineer Research and Development Center
Arts G H P, van der Heijden R A J M, 1990. Germination ecology of Littorella uniflora (L.) Aschers. Aquatic Botany, 37(2):139—151
Brenchley J L, Probert R J, 1998. Seed germination responses to some environmental factors in the seagrass Zostera capricorni from eastern Australia. Aquatic Botany, 62(3):177—188
Caye G, Bulard C, Meinesz A et al, 1992. Dominant role of seawater osmotic pressure on germination in Cymodocea nodosa. Aquatic Botany, 42(2): 187—193
Churchill A C, 1983. Field studies on seed germination and seedling development in Zostera marina L. Aquatic Botany,16(1): 21—29
Conacher CA, Poiner I R, Butler J et al, 1994. Germination,storage and viability testing of seeds of Zostera capricorni Aschers. from a tropical bay in Australia. Aquatic Botany,49(1): 47—58
Costello C T, Kenworthy W J, 2011. Twelve-year mapping and change analysis of eelgrass (Zostera marina) areal abundance in Massachusetts (USA) identifies statewide declines. Estuaries and Coasts, 34(2): 232—242
Delefosse M, Kristensen E, 2012. Burial of Zostera marina seeds in sediment inhabited by three polychaetes: Laboratory and field studies. Journal of Sea Research, 71: 41—49
Dennison W C, 1987. Effects of light on seagrass photosynthesis,growth and depth distribution. Aquatic Botany, 27(1):15—26
Fernández-Torquemada Y, Sánchez-Lizaso J L, 2013. Effects of salinity on seed germination and early seedling growth of the Mediterranean seagrass Posidonia oceanica (L.) Delile.Estuarine, Coastal and Shelf Science, 119: 64—70
Fong P, Fong J J, Fong C R, 2004. Growth, nutrient storage, and release of dissolved organic nitrogen by Enteromorpha intestinalis in response to pulses of nitrogen and phosphorus.Aquatic Botany, 78(l): 83—95
Fourqurean J W, Duarte C M, Kennedy H et al, 2012. Seagrass ecosystems as a globally significant carbon stock. Nature Geoscience, 5(7): 505—509
Harrison P G, 1991. Mechanisms of seed dormancy in an annual population of Zostera marina (eelgrass) from the Netherlands. Canadian Journal of Botany, 69(9):1972—1976
Hay F, Probert R, Dawson M, 2008. Laboratory germination of seeds from 10 British species of Potamogeton. Aquatic Botany, 88(4): 353—357
Hootsmans M J M, Vermaat J E, Van Vierssen W, 1987.Seed-bank development, germination and early seedling survival of two seagrass species from The Netherlands:Zostera marina L. and Zostera noltii hornem. Aquatic Botany, 28(3—4): 275—285
ISTA, 1996. International Rules for seed testing. Seed Science and Technology. Zürich, Switzerland: ISTA
Jarvis J C, Moore K A, 2008. Influence of environmental factors on Vallisneria americana seed germination. Aquatic Botany,88(4): 283—294
Kahn A E, Durako M J, 2005. The effect of salinity and ammonium on seed germination in Ruppia maritima from Florida Bay. Bulletin of Marine Science, 77(3): 453—458
Karssen C M, Hilhorst H W, Koomneef M, 1990. The benefit of biosynthesis and response mutants to the study of the role of abscisic acid in plants. In: Pharis P R P, Rood P S B eds.Plant Growth Substances. Berlin Heidelberg, Germany:Springer, 23—31
Ke X S, Li W, 2006. Germination requirement of Vallisneria natans seeds: implications for restoration in Chinese lakes.Hydrobiologia, 559(1): 357—362
Kendrick G A, Waycott M, Carruthers T J B et al, 2012. The central role of dispersal in the maintenance and persistence of seagrass populations. BioScience, 62(1): 56—65
Kettenring K M, Galatowitsch S M, 2007. Temperature requirements for dormancy break and seed germination vary greatly among 14 wetland Carex species. Aquatic Botany,87(3): 209—220
Koch E W, Ailstock M S, Booth D M et al, 2010. The role of currents and waves in the dispersal of submersed angiosperm seeds and seedlings. Restoration Ecology, 18(4):584—595
Krause-Jensen D, Greve T M, Nielsen K, 2005. Eelgrass as a bioindicator under the European Water Framework Directive.Water Resources Management, 19(1): 63—75
Lal C, Gopal B, 1993. Production and germination of seeds in Hydrilla verticillata. Aquatic Botany, 45(2—3): 257—261
Lee K S, Short F T, Burdick D M, 2004. Development of a nutrient pollution indicator using the seagrass, Zostera marina, along nutrient gradients in three New England estuaries. Aquatic Botany, 78(3): 197—216
Li W T, Kim S H, Kim J W et al, 2013. An examination of photoacclimatory responses of Zostera marina transplants along a depth gradient for transplant-site selection in a disturbed estuary. Estuarine, Coastal and Shelf Science, 118:72—79
Loquès F, Caye G, Meinesz A, 1990. Germination in the marine phanerogam Zostera noltii Hornemann at Golfe Juan, French Mediterranean. Aquatic Botany, 38(2—3): 249—260
McGlathery K J, Pedersen M F, Borum J, 1996. Changes in intracellular nitrogen pools and feedback controls on nitrogen uptake in Chaetomorpha linum (chlorophyta).Journal of Phycology, 32(3): 393—401
Olesen B, Sand-Jensen K, 1993. Seasonal acclimatization of eelgrass Zostera marina growth to light. Marine Ecology Progress Series, 94: 91—99
Orth R J, Carruthers T J B, Dennison W C et al, 2006. A global crisis for seagrass ecosystems. BioScience, 56(12):987—996
Orth R J, Harwell M C, Bailey E M et al, 2000. A review of issues in seagrass seed dormancy and germination:implications for conservation and restoration. Marine Ecology Progress Series, 200: 277—288
Orth R J, Marion S R, Moore K A et al, 2010. Eelgrass (Zostera marina L.) in the Chesapeake Bay region of mid-Atlantic coast of the USA: challenges in conservation and restoration.Estuaries and Coasts, 33(1): 139—150
Orth R J, Moore K A, 1983. Seed germination and seedling growth of Zostera marina L. (eelgrass) in the chesapeake bay. Aquatic Botany, 15(2): 117—131
Pan J H, Jiang X, Li X J et al, 2011. Influence of temperature and salinity on germination of eelgrass (Zostera marina L.) seeds.Journal of Ocean University of China, 10(2): 147—152
Pons A, Guiot J, de Beaulieu J L et al, 1992. Recent contributions to the climatology of the last glacial-interglacial cycle based on French pollen sequences. Quaternary Science Reviews,11(4): 439—448
Probert R J, Brenchley J L, 1999. The effect of environmental factors on field and laboratory germination in a population of Zostera marina L. from southern England. Seed Science Research, 9(4): 331—339
Ranal M A, de Santana D G, 2006. How and why to measure the germination process? Brazilian Journal of Botany, 29(1):1—11
Reynolds L K, Waycott M, McGlathery K J et al, 2012. Eelgrass restoration by seed maintains genetic diversity: case study from a coastal bay system. Marine Ecology Progress Series,448: 223—233
Rivard P G, Woodard P M, 1989. Light, ash, and pH effects on the germination and seedling growth of Typha latifolia(cattail). Canadian journal of botany, 67(9): 2783—2787
Strazisar T, Koch M S, Madden C J et al, 2013. Salinity effects on Ruppia maritima L. seed germination and seedling survival at the Everglades-Florida Bay ecotone. Journal of Experimental Marine Biology and Ecology, 445: 129—139
Sugiura H, Hiroe Y, Suzuki T et al, 2009. The carbohydrate catabolism of Zostera marina influenced by lower salinity during the pre-germination stage. Fisheries Science, 75(5):1205—1217
Tanner C E, Parham T, 2010. Growing Zostera marina (eelgrass)from Seeds in Land-Based Culture Systems for Use in Restoration Projects. Restoration Ecology, 18(4): 527—537
Taylor A R A, 1957. Studies of the development of Zostera marina L.: II. Germination and seedling development.Canadian Journal of Botany, 35(5): 681—695
Vandelook F, Van Assche J A, 2008. Temperature requirements for seed germination and seedling development determine timing of seedling emergence of three monocotyledonous temperate forest spring geophytes. Annals of Botany, 102(5):865—875
