封山育林对不同发育阶段杉木林植物多样性的影响
2015-12-30吴初平沈爱华袁位高朱锦茹戚连忠
张 骏,葛 滢,吴初平,沈爱华,江 波,袁位高,朱锦茹,戚连忠,常 杰
(1. 浙江林业科学研究院,浙江 杭州 310023;2. 浙江大学,浙江 杭州 310058)
封山育林对不同发育阶段杉木林植物多样性的影响
张 骏1,葛 滢2,吴初平1,沈爱华1,江 波1,袁位高1,朱锦茹1,戚连忠1,常 杰2
(1. 浙江林业科学研究院,浙江 杭州 310023;2. 浙江大学,浙江 杭州 310058)
将浙江省杉木公益林划分为3个发育阶段:≤10 a为幼龄林;11 ~ 20 a为中龄林;> 20 a为成熟林。对杉木(Cunninghamia lanceolata)优势林和含杉木混交林的三个发育阶段的乔木层、下木层和草本层的植物多样性进行比较研究。结果表明:杉木优势林的乔木层植物多样性指数都随时间缓慢上升(< 15%),成熟林乔木层的异质性和均匀性高于草本层,幼龄林下木层多样性不高,物种数和异质性也都随时间缓慢上升(< 5%),下木层均匀性在中龄以后稳定,杉木优势林随着林龄的增加,郁闭度相应增加使得草本层的Gleason指数从2.62下降到1.68;含杉木混交林封山育林后幼龄林下木层植物多样性最高,中龄林的物种数、异质性最低,均匀度随时间逐渐降低,各层植物多样性指数在各个发育阶段都表现为下木层 > 乔木层 > 草本层,仅有草本层Gleason指数中幼龄林阶段较高例外;在各个发育阶段杉木优势林的乔木层和下木层的植物多样性都不如含杉木混交林,优势林草本层的物种丰富度、异质性和均匀性指数幼龄林阶段都高于含杉木混交林,中龄林阶段接近,在成熟林阶段都比含杉木混交林低;总体而言,杉木优势林的植物多样性不如相同发育阶段的含杉木混交林。
生态公益林;杉木;植物多样性;优势林;浙江省
在当今生态环境日益恶化,人民群众对生态产品要求日益提高的大背景下,生态公益林的建设已成为改善生态环境的重要组成部分[1~2]。生态公益林的效益包括生态效益、经济效益和社会效益。生物多样性是恢复生态学研究的主要内容之一,也是体现公益林生态效益的重要方面[3~4]。物种多样性是揭示植被组织水平的生态基础,反映生物群落在组成、结构、功能和动态等表现出的异质性[5],因此植物群落的多样性作为显示植物群落组成结构的重要指标,一直受到生态学家的关注[6~8]。对杉木(Cunninghamia lanceolata)人工林不同发育阶段发展过程中植物多样性变化的研究,可探明杉木林植物多样性现状,为开展生物多样性保护和指导杉木林的人工管理提供理论依据[4]。
浙江省是我国经济发达省份之一,高速的经济发展和人口增长带来了土地利用与土地覆盖的剧烈变化,也对生态环境造成显著影响[9]。建国初期由于社会改革和经济建设的发展,对木材需求量大,20世纪50年代起杉木造林面积迅速增加[10]。但是随着社会发展,杉木的经济价值已经越来越小,有些地区的杉木林已经超过更新年龄却无人砍伐;部分地区为了提高杉木林生产力而加强人为管理,从而对植物多样性产生影响。从2001年起,随着公益林建设工程推进,分布在水源涵养林区、生态保护植被恢复和森林生态治理区等的杉木林作为公益林进行封育,面积达10.53万hm2,占全省公益林面积的12.8%[11]。这些杉木林封山育林已近15年,不能再按经济利用方式进行砍伐,但其较低的生物多样性特征必将严重限制公益林的生态效益。
为了定量分析封山育林后杉木林的生态效益,本研究以不同发育阶段的杉木林为研究对象,调查和计算不同林型的植被多样性指数,进而预测杉木林封育后林下植被多样性恢复潜力,并探讨定向改造的方法,从而为提高杉木公益林生态效益提供理论依据和实践手段。
1 研究区自然概况
浙江省位于长江中下游沿海地区,省内山脉多是西南—东北走向,高山基本集中于西南部,平均海拔为800 m。浙江省属于典型的亚热带气候,水热基本同期[12];年均气温15.3 ~ 18.5℃,最低月均温2.7 ~ 7.9℃,最高月均温27.0 ~ 29.5℃,≥10℃年积温4 800 ~ 5 800℃,全年无霜期225 ~ 280 d;年降水量1 000 ~ 2 000 mm,以春雨、梅雨、台风雨为主,7-8月有伏旱[1]。土壤类型多以红壤、红黄壤和黄壤为主,还有少量的石灰土、紫色土等,适合于森林群落的生长,故境内森林群落类型丰富,植物种类繁多。浙江全省范围均属于亚热带常绿阔叶林区域—东部(湿润)常绿阔叶林亚区—中亚热带常绿阔叶林地带,故地带性植被为常绿阔叶林、常绿阔叶和落叶混交林;除此之外还有落叶阔叶林、针叶林、竹林等多种植被类型[11];同时浙江省还位于杉木栽培区域中带,杉木栽培历史悠久[10]。
2 研究方法
2.1 样地设置
在代表浙江省各种环境条件的23个县里,选取杉木林样地67个,其中测定林龄的样地45个。样地分布在浙西北、浙中、浙南和浙东南沿海,海拔50 ~ 650 m,坡度10 ~ 40°,平均胸径4.3 ~ 14.0 cm,林龄6 ~ 30 a[1]。群落类型除杉木优势林(简称优势林)外还包括杉木—苦槠混交林、杉木—木荷混交林、杉木—苦楝混交林和杉木—拟赤杨混交林等含杉木混交林(简称混交林)[13]。
对选定的杉木林样地采用方精云等[14]在中国山地植物物种多样性中的方法[1]进行群落植被结构、数量特征以及海拔高度和坡度等指标的调查。其中乔木层样地面积为600 m2,即20 m×30 m,调查乔木树种的每株胸径、树高、冠幅等;下木层样带面积36 m2,即在乔木层20 m×30 m的样地对角线的1 m宽样带中,调查每一树种的株数、树高并统计各树种的数量与平均高度;草本层在样地对角线端部设2个2 m×2 m的小样方,调查小样方内各草本植物种数量和盖度。
2.2 多样性测定方法
物种多样性是一个群落结构和功能复杂性的度量,有3个参数与之密切相关,即物种丰富度、物种间差异度和物种均匀度[15]。物种多样性的测度方法国内外研究很多[16 ~ 18],本文采用如下:
物种丰富度R:R = S
Gleason指数Gl:Gl= S/lnA。Gini指数D:D = 1-∑Pi2
Shannon-Wiener指数H’:H’ =-∑PilnPi(∑Pi= 1)。
式中,A为样方面积,Pi为物种i的相对重要值[19],乔木层物种相对重要值为乔木的相对密度、相对胸高断面积之和和相对频度的平均值,灌木和草本层物种的相对重要值为各自的相对密度、相对盖度和相对频度的平均值[20],数据的其他统计分析采用Excel进行。
2.3 数据处理
杉木林发育阶段的划分方式各有差异,秦建华等[21]以1 ~ 10 a为幼龄林,11 ~ 20 a为中龄林,21 ~ 25 a为近熟林,> 25 a为成熟林分4个发育阶段;陈楚莹等[22]从杉木林的生长发育特点出发划分为4个阶段:幼树阶段、速生阶段、干材阶段和成熟阶段。参照浙江林业自然资源的林龄划分标准[11]和对生态公益林的要求,本研究将浙江省杉木公益林划分为3个发育阶段:≤10 a为幼龄林;11 ~ 20 a为中龄林;> 20 a为成熟林[1]。
本文从杉木占乔木层的相对密度以及相对重要值考虑[23],将杉木相对密度70%以上或相对重要值0.60以上的样地划为优势林,其他则为混交林[1]。共67个杉木样地划分为优势林地28个,混交林地39个。
3 结果与分析
3.1 乔木层植物多样性
优势林乔木层中杉木的相对密度在幼龄林和中龄林阶段都很高,占75%以上,成熟林阶段降低到了35.73%;从幼龄林到中龄林阶段物种数和多样性指数均缓慢上升(幅度小于15%),成熟林阶段植物多样性指数有较大幅度上升(幅度大于77 %)。混交林乔木层的物种数和Gleason指数增长迅速,三个发育阶段间几乎成倍递增;Shannon-Wiener指数也逐渐上升但变化幅度较小不到30 %;Gini指数在0.73水平保持稳定。
相同发育阶段的杉木优势林乔木层的各个多样性指数在20年前只有混交林的四分之一左右;成熟林阶段两者乔木层的物种数差距增大,其他多样性指数的差距减小。各个阶段优势林乔木层的物种丰富度、异质性和均匀度都低于混交林乔木层(表1)。

表1 不同发育阶段杉木优势林和含杉木混交林乔木层植物多样性比较Table 1 Plant diversity at tree layer of pure C. lanceolata forest and mixed fores in different development stage
3.2 下木层植物多样性
优势林下木层的S、Gl和D都在幼龄林阶段最小,随林龄增加呈上升趋势,上升幅度逐渐减小;H’在成熟林阶段保持稳定略有下降。混交林的各个指数随年龄的变化与优势林相反,均在幼龄林阶段出现最大值,中龄林和成熟林阶段变化较小(表2)。
相同发育阶段的下木层各个多样性指数,都是优势林低于含杉木混交林,但两者下木层的差距不如乔木层大,不到 1.5倍。优势林下木层多样性指数随着年龄增长的变化规律与混交林相反,所以到成熟林阶段,优势林下木层多样性指数与混交林的下木层多样性指数之间的差距在减小。
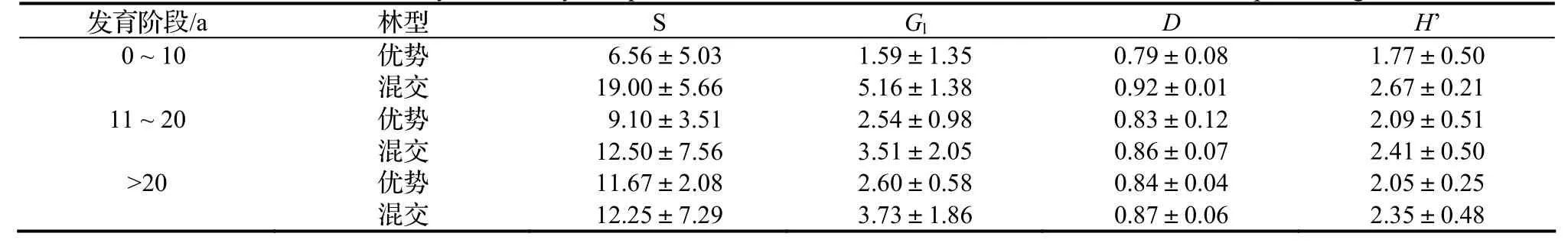
表2 各发育阶段杉木优势林和含杉木混交林下木层植物多样性比较Table 2 Plant diversity at shrub layer of pure C. lanceolata forest and mixed forest in different development stage
3.3草本层植物多样性
优势林草本层的物种丰富度、异质性和均匀性都在中龄林最大。混交林中龄林草本层的物种丰富度、异质性和均匀性比幼龄林增长近一倍(> 74%),成熟林与中龄林接近,略微下降。
草本层Gleason指数在10 a以前一直优势林高于混交林,到成熟林阶段优势林草本层的S只有混交林的58.1%。D和H’在幼龄林阶段优势林高于混交林,到中龄林阶段两者差距缩小,成熟林阶段后者超过前者(表3)。
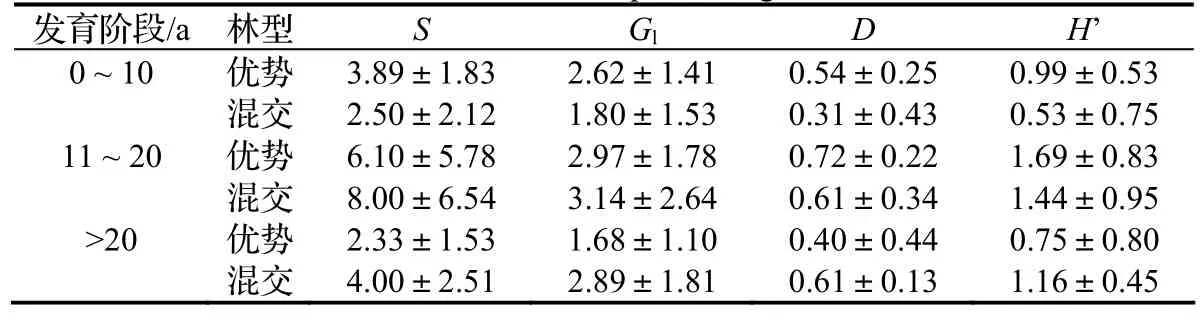
表3 各发育阶段杉木优势林和含杉木混交林草本层植物多样性比较Table 3 Plant diversity at herb layer of pure C. lanceolata forest and mixed forest in different development stage
3.4群落各层次之间比较
优势林和混交林的乔木层各个多样性指数平均值都低于下木层(表4),经显著性分析,两种林型的乔木层与下木层植物多样性差异极显著(P < 0.01),表明两者差异较大,即乔木层与下木层的乔木幼树、幼苗物种雷同程度极小。
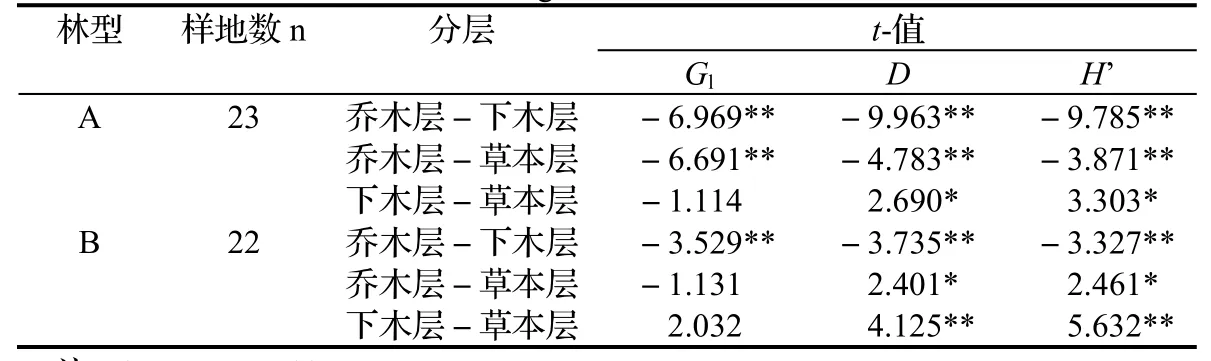
表4 杉木优势林和含杉木混交林型各层次间植物多样性的显著性检验(t-检验)Table 4 T-test of biodiversity in each layer of C. lanceolata dominanteded forest and mixed forest containing C. lanceolata
4 讨论
浙江省杉木公益林中的优势林封山育林后,乔木层植物多样性指数都随时间缓慢上升(< 15%),下木层幼龄林阶段多样性不高,物种数和异质性也都随时间缓慢上升(< 5%),林龄>20 a乔木层的异质性和均匀性高于草本层,下木层均匀性在中龄以后稳定。优势林随着林龄的增加,郁闭度相应增加使得草本层的Gleason指数从2.62下降到1.68,但光照仍然相对充足,草本植物丰富,所以草本层植物多样性一直高于乔木层[24]。优势林在>20 a后,杉木相对密度降低到了35.73,郁闭度的减少使得草本层的Gleason指数达到2.97(表3),但其他如木荷、柳杉、杨梅、油茶、山樱桃等常绿树种迅速进入乔木层使得郁闭度加大,这可能受到人为干扰(如间伐)影响所致[25~26];到了成熟林阶段,下木层旺盛的生长发育一定程度上抑制了草本层,故草本层的异质性和均匀性不如相同发育阶段的乔木层和下木层[27]。
杉木公益林中的混交林封山育林后,下木层幼龄林阶段植物多样性最高,中龄林阶段的物种数、异质性最低,均匀度随时间逐渐降低;各层植物多样性指数在各个发育阶段都表现为下木层>乔木层>草本层,仅有草本层Gleason指数中幼龄林阶段高于乔木层例外,这是因为混交幼、中龄林的上层郁闭度小,下层有一定比例的灌木和草本生长[10],故混交林的下木层物种比乔木层丰富。林龄 > 20 a的混交林,由于许多阔叶树种进入乔木层使得郁闭度增大和林下光照减少,引起草本植物稀疏,不仅各物种个体数分配不均匀,而且物种在群落中的空间分布也很不均匀,因此草本层的各个多样性指数都不如乔木层。
封山育林后的优势林不论乔木层还是下木层的植物多样性,在各个发育阶段都不如混交林;而草本层优势林的物种丰富度、异质性和均匀性指数幼龄林阶段都高于混交林,中龄林阶段接近,在成熟林阶段都比混交林低。因此总体而言,优势林的植物多样性不如相同发育阶段的混交林。
根据多数学者观点,尽管物种多样性与系统稳定性并非永远表现为正相关,但较高的物种多样性一般都能提高系统稳定性[28~30]。田大伦等[4]认为杉木林群落的植物多样性与稳定性相关,物种的多样性和复杂性增加了群落的稳定性,为此仅从生态效益的角度来说,非常有必要对杉木优势林进行以含杉木混交林为目标的定向改造。盛炜彤等[26]认为适度间伐对生态系统干扰较小,择伐后补种苦槠、青冈、木荷、檵木、柃木和冬青等其他常绿树种容易迅速恢复亚热带地区森林生态系统的功能[1];这和王祖华等[31]、蔡道雄等[32]的研究一致,认为间伐显著提高了林下植被的物种丰富度,有利于林下植被的恢复和优势群落的建立。因此对杉木公益林的管理,建议结合林下抚育改造,通过间伐补种加快形成多种植物组成、多种层次的顶极群落或稳定性较强的混交次生林,从而实现杉木纯林的近自然改造。
[1]张骏,葛滢,江波,等. 浙江省杉木生态公益林碳储量效益分析[J]. 林业科学,2010,46(6):22-26.
[2]周国逸,闫俊华. 生态公益林补偿理论与实践[M]. 北京:气象出版社,2000.
[3]方奇. 不同密度杉木幼林系统生产力和生态效益研究. 林业科学,2000,36(1):28-35.
[4]田大伦,康文星,文仕知,等. 杉木林生态系统学[M]. 北京:科学出版社,2003.
[5]兰思仁. 武夷山国家级自然保护区植物物种多样性研究[J]. 林业科学,2002,39(1):36-43.
[6]Magurran A E. Ecological diversity and its Measurement[M]. New Jersey: Princeton University Press, 1988.
[7]马克平. 生物多样性的测定方法[M]. 北京:中国科学技术出版社,1994.
[8]易俗,黄忠良,欧阳学军. 鼎湖山生物圈保护区层间植物物种多样性的研究[J]. 生物多样性,2001,9(1):56-61.
[9]张骏,高洪娣,应宝根,等. 浙江省仙居县公益林生物量动态分析[J]. 南京林业大学学报(自然科学版)2011,35(5):147-150.
[10]俞新妥,范少辉,林思祖,等. 杉木栽培学[M]. 福州:福建科学技术出版社,1997.
[11]刘安兴,张正寿,丁衣冬,等. 浙江林业自然资源(森林卷)[M]. 北京:中国农业科学技术出版,2002.
[12]浙江森林编辑委员会. 浙江森林[M]. 北京:中国林业出版社,1993.
[13]黄承才,张骏,江波,等. 浙江省杉木生态公益林凋落物及其与植物多样性的关系[J]. 林业科学,2006,42(6):7-12.
[14]方精云,沈泽昊,唐志尧,等. “中国山地植物物种多样性调查计划”及若干技术规范[J]. 生物多样性,2004,12(1):5-9.
[15]常杰,葛滢. 生态学[M]. 杭州:浙江大学出版社,2001.
[16]Michaĕl A, Didier A, Fabrice B. Diversity of plant assemblages in managed temperate forests: a case study in Normandy (France)[J]. For Ecol Manage, 2003(175):321-337.
[17]贺金生,陈伟烈,李凌浩. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征[J]. 植物生态学报,1998,22(4):303-311.
[18]刘世荣,蒋有绪,史作民,等. 中国暖温带森林生物多样性研究[M]. 北京:中国科学技术出版社,1998.
[19]宋永昌. 植被生态学[M]. 上海:华东师范大学出版社,2001.
[20]姜汉侨,段昌群,杨树华,等. 植物生态学[M]. 北京:高等教育出版社,2004.
[21]秦建华,姜志林. 杉木林生物量极其分配变化的规律[J]. 生态学杂志,1996,135(1):1-7.
[22]陈楚莹,廖利平,汪思龙. 杉木人工林生态学[M]. 北京:科学出版社,2000.
[23]金则新. 浙江天台山落叶阔叶林优势种群结构与动态分析[J]. 浙江林学院学报,2001,183(3):245-251.
[24]胡正华,于明坚,丁炳扬,等. 古田山国家级自然保护区常绿阔叶林类型及其群落物种多样性研究[J]. 应用与环境生物学报,2003,9 (4):341-345.
[25]叶镜中,姜志林. 苏南丘陵杉木人工林的生物量结构[J]. 生态学报,1983,3(1):7-14.
[26]盛炜彤,范少辉. 杉木人工林的育林干扰对长期立地生产力的影响[J]. 林业科学,2003,39(5):37-43.
[27]林开敏,俞新妥,黄宝龙,等. 杉木人工林林下植物物种多样性的动态特征[J]. 应用与环境生物学报,2001,7(1):13-19.
[28]Bengtsson J, S G Nilsson, Franc A, et al. Biodiversity, disturbances, ecosystem function and management of European forests[J]. Forest Ecology and Management, 2000(32):39-50.
[29]Loreau M, Naeem S, Inchausti P, et al. Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges[J]. Science, 2001 (294):804-808.
[30]Tilman D, Reich P B, Johannes Knops, et al. Diversity and Productivity in a Long-Term Grassland Experiment[J]. Science , 2001(294):843 -845.
[31]王祖华,李瑞霞,王晓杰,关庆伟. 间伐对杉木人工林林下植被多样性及生物量的影响[J]. 生态环境学报,2010,19(12):2 778-2 782.
[32]蔡道雄,卢立华,贾宏炎,等. 封山育林对杉木人工林林下植被物种多样性恢复的影响[J]. 林业科学研究,2007,20(3):319-327.
Influences of Conservation for Regeneration on Plant Diversity of Ecological Cunninghamia lanceolata Forest at Different Development Stage in Zhejiang
ZHANG Jun1,GE Ying2,WU Chu-ping1,SHEN Ai-hua1,JIANG Bo1,YUAN Wei-gao1,ZHU Jin-ru1,QI Lian-zhong1,CHANG Jie2*
(1. Zhejiang Forestry Academy, Hangzhou 310023, China; 2. Zhejiang University, Hangzhou 310058, China)
Ecological Cunninghamia lanceolata forest was divided into three development stages(≤10-year as young, 11-20-year as half-mature and > 20-year as mature) and two forest types(pure C. lanceolata forest and mixed forest with C. lanceolata) in Zhejiang province. 67 sample plots in 23 counties were established for monitoring plant diversities at tree, shrub and herb layer in different forest types. The results showed that at tree layer of pure C. lanceolata forest, plant diversity index increased slowly (<15%), and heterogeneity and homogeneity was higher than that at herb layer in mature forest, while at shrub layer, plant diversity index of pure C. lanceolata forest was not high at young stage, species numbers and heterogeneity increased slowly (<5%). Gleason index at herb layer decreased from 2.62 to 1.68 because of the increase of closure. Mixed young forest had the highest plant diversity index at shrub layer, half-mature one had the lowest species numbers and heterogeneity. Plant diversity index at different layer in different development stages was ordered by shrub layer>tree layer> herb layer, Gleason index of herb layer was an exception. Plant diversity at tree and shrub layer of pure C. lanceolata forest with different development stage was lower than mixed forest. Species richness, heterogeneity andevenness index of herb layer in pure young C. lanceolata forest was higher than that in mixed forest, and got close in half-mature one, but lower in mature than that in mixed forest. The study found that plant diversity of pure C. lanceolata forest was lower than that of mixed forest in the same developmental stage.
ecological forest; Cunninghamia lanceolata; plant diversity; Zhejiang
S718
A
1001-3776(2015)05-0011-05
2015-05-09;
2015-08-10
浙江省自然科学基金项目(LQ13C0301);森林生态科技创新团队(2011R50027);浙江省省级定位站管理维护及技术支撑项目
张骏(1981-),男,浙江龙游人,博士,副研究员,从事森林生态研究;*通讯作者。
