不同种源栓皮栎种子表型性状的变异分析
2015-12-22李迎超丁可君刘国军厉月桥中国林业科学研究院林业研究所国家林业局林木培育重点实验室北京0009北京中林林业科技开发公司北京0009新余学院江西新余00北京市科学技术委员会农村发展中心北京0009邢台市林业局河北邢台0000唐山市嘉城园林绿化工程有限公司河北唐山0000中国林业科学研究院亚热带林业实验中心江西分宜00河北农业大学林学院河北保定0000
许 洋,李迎超,冯 慧,王 迪,丁可君,郭 超,刘国军,厉月桥,宁 超,贺 磊,郭 芳 (.中国林业科学研究院林业研究所国家林业局林木培育重点实验室北京0009;.北京中林林业科技开发公司北京0009;.新余学院 江西新余00;.北京市科学技术委员会农村发展中心 北京0009;.邢台市林业局 河北邢台0000;.唐山市嘉城园林绿化工程有限公司 河北唐山0000;.中国林业科学研究院亚热带林业实验中心江西分宜00;.河北农业大学林学院河北保定0000)
栓皮栎(Quercus variabilis)为高大乔木,隶属壳斗科(Fagaceae)栎属(Quercus Linn.)植物,是我国暖温带海拔1 600 m以下地区主要的地带性植物组成树种,树高可达30 m,胸径可达1 m,是我国分布极为广泛的树种之一。栓皮栎的木材有光泽,具有花纹美丽、致密坚实、耐冲击、强度大、耐腐蚀等优良特性。栓皮栎还是传统的薪炭资源树种,其木材和枝丫是木耳、香菇、天麻等食用菌生产的优质原料[1-3]。栓皮栎和栓皮槠(Quercus suber)的栓皮层通常称为“软木”(cork或phellogen),与针叶材软木(softwood)不同,其独特的细胞结构及化学成分使其具有阻燃、防水、防潮、隔水、隔音、隔热、弹性好、质轻柔软、耐磨等优良特性,素有“软黄金”之称[4],软木在欧美被认为是最理想的室内装修和装饰材料。栓皮栎果实内淀粉含量可达51% ~59%[5],其乙醇生产潜力可满足我国目前E10乙醇汽油需求的0.63%,是发展生物质燃料乙醇的重要补充原料[6]。
国内对栓皮栎的研究涉及多个方面,包括生物学特性[7-9]、地理分布[10-14]、资源培育[15-19]、群落特征[20]、种群动态[21-24]、群落物种多样性[25]、生态位[26]、橡实淀粉燃料酒精生产工艺[27]等方面,而对栓皮栎遗传多样性以及种子表型变异的研究较少,加之栓皮栎分布范围广造成地理种源变异,使得在种子调拨育苗方面存在盲目性,影响了规模化引种栽培及其发展。种子表型变异具有较高的稳定性,是生物遗传变异最直接的表现,因此,了解栓皮栎种子表型变异规律,可以在较短时间内获得该树种变异趋势的一般规律,并且是种子鉴别、种子区划、种子检验和播种育苗等工作的基础和前提,可为种源试验采样和试验设计提供重要依据,同时也是遗传育种的基础。该研究在全面踏查栓皮栎自然分布区的基础上,对来自8个省15个地理种源栓皮栎种子表型性状变异进行分析,探讨种子性状变异规律及其与环境因子的关系,为确定适宜的优良种源及开展栓皮栎的遗传改良和品种选育提供试验依据。
1 材料与方法
1.1 种子来源及采样地自然条件 该研究所用材料是来自栓皮栎自然分布区的北京、安徽、陕西、河北、山东、河南、云南、湖南等8个省15个地理种源,基本覆盖栓皮栎的自然分布区[28]。分别于2013、2014年2年中8~11月开展种子收集工作,所采种源样本均为当年成熟的种子。具体采集地点、主要气候因子及代号参考李迎超[29]对栓皮栎地理种源的调查。
1.2 试验方法 将从各个种源所采集的种子,分别采用四分法随机抽取300粒种子,每100粒为1组,3次重复,用游标卡尺分别测定种子的种宽和种长,以腹面横向最大宽度代表种宽,以种子纵轴代表种长,精确到0.01 mm,计算种长和种宽的比值。用1/100电子天平测定单果重,精确到0.01 g,记录数据。
1.3 数据分析 用Excel 2010软件进行基础数据的整理、统计,建立栓皮栎种子性状原始数据文档,运用SPSS 18.0软件进行方差分析、多重比较、相关分析以及聚类分析。
2 结果与分析
2.1 不同栓皮栎种源间种子的表型变异及多重比较分析 不同栓皮栎种源间种子表型性状变异分析见表1。从表1可以看出,在种子表型特征方面,15个种源栓皮栎种子宽度最小的是陕西镇坪种源(12.50 mm),最大的是陕西黄龙种源(21.34 mm),后者为前者的1.71倍,15个种源栓皮栎种子宽度变异幅度为12.50~21.34 mm;种子长度最小的是安徽滁州皇甫山种源(20.17 mm),最大的是山东牟平种源(24.30 mm),最大值为最小值的 1.20倍,15个种源栓皮栎种子长度变异幅度为20.17~24.30 mm;种子长宽比最小的是安徽滁州皇甫山种源(1.06),最大的是陕西镇坪种源(1.79),后者是前者的1.69倍,15个种源栓皮栎种子长宽比变异幅度为1.06~1.79;单果重最小的是陕西镇坪种源(1.55 g),最大的是山东牟平种源(4.98 g),后者是前者的3.21倍,15个种源栓皮栎单果重变异幅度为1.55 ~4.98 g。方差分析结果表明,不同种源间栓皮栎种子长、种子宽、种子长宽比、单果重等性状差异均达到极显著水平。
变异系数表示性状的离散程度,在种子的4个表型性状中,单果重的变异系数最大,为17.06%,种子宽、种子长及种子长宽比的变异系数在6.66% ~7.68%之间,说明种子长和宽指标的离散程度较小,具有一定的稳定性,同时也表明不同地域的环境因子对表型性状变异具有一定的影响。
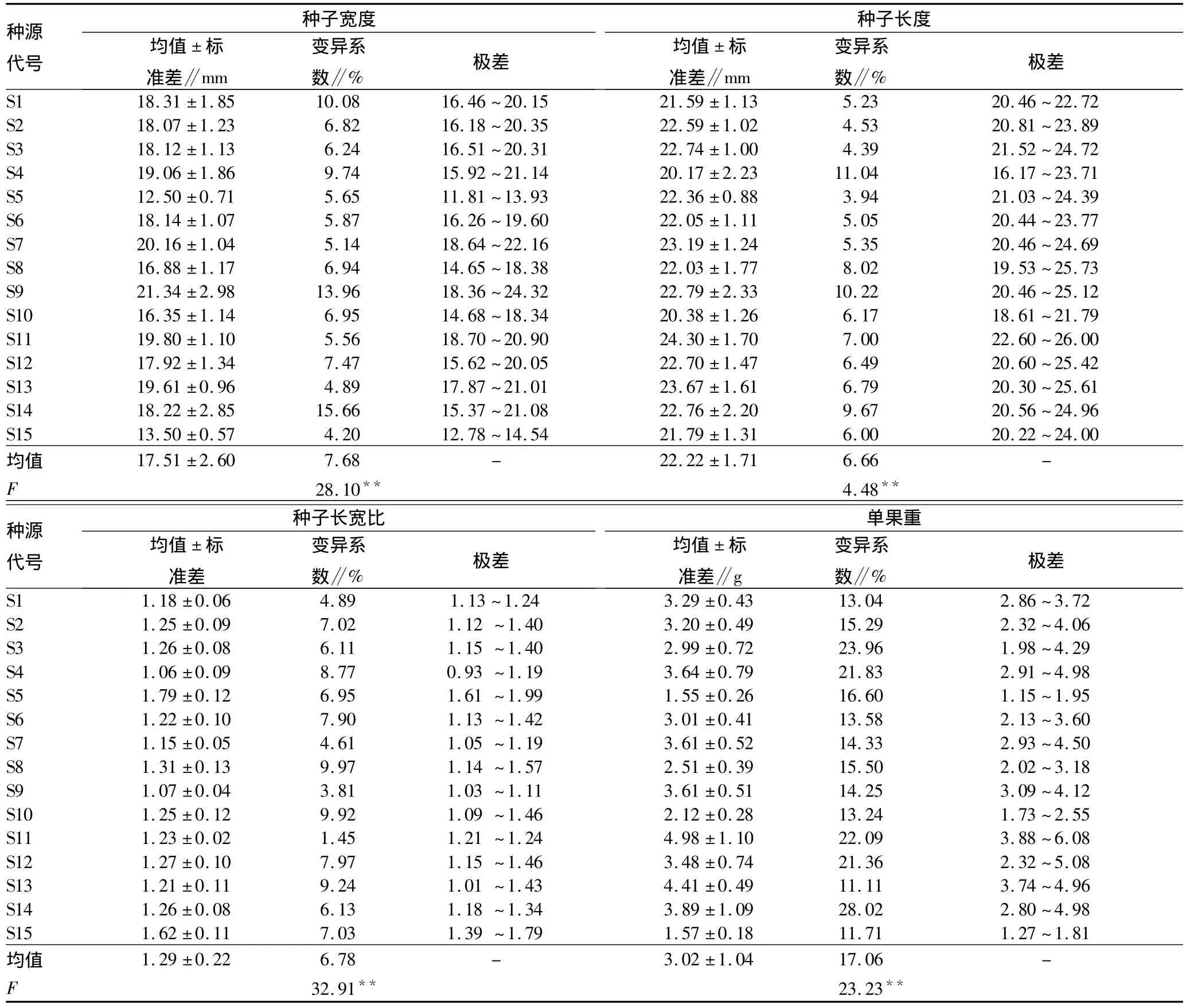
表1 栓皮栎种源间种子表型性状变异
2.2 栓皮栎种源间种子各表型性状的相关性分析 供试栓皮栎种源种子表型性状的相关系数见表2。由表2可知:在栓皮栎种子4个表型性状中,种子宽和单果重与种子长宽比呈极显著负相关(P<0.01),其他性状两两间呈正相关,其中种子宽与单果重呈极显著正相关(P<0.01),说明随着种子宽度的增加,种子单果重显著增加。种子长度与单果重呈显著正相关(P<0.05),而种子其他性状间均未达显著性相关水平(P≥0.05)。
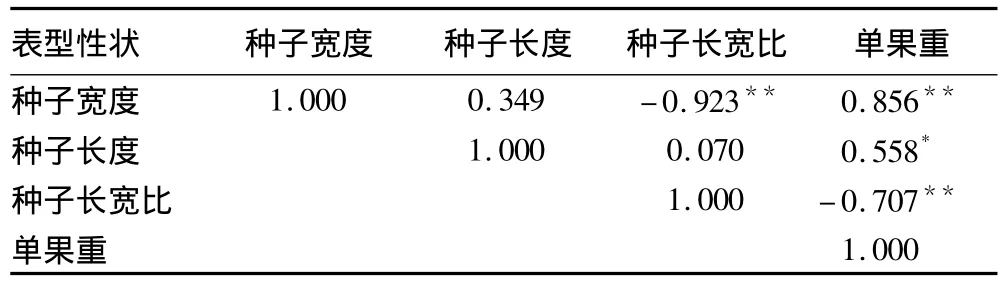
表2 栓皮栎种源种子表型性状间的相关系数
2.3 栓皮栎种子表型性状与生态因子的地理变异分析 栓皮栎种子表型性状与生态因子的地理变异分析见表3。由表3可知,种子宽度与经度、纬度和日照时数呈正相关,与海拔、年均温、1月均温、7月均温、年降水量、无霜期呈负相关,但种子宽度与所有生态因子指标相关性未达到显著水平;种子长度与经度、纬度、7月均温、日照时数呈正相关,与海拔、年均温、1月均温、年降水量、无霜期呈负相关,但都没有达到显著水平;单果重与经度、纬度、7月均温、年降水量、日照时数呈正相关,与海拔、年均温、1月均温、无霜期呈负相关,其中单果重与经度呈显著正相关(P<0.05),即自西往东,种子单果重逐渐增大;种子长宽比与海拔、年均温、1月均温、7月均温、年降水量、无霜期呈正相关,与经度、纬度、日照时数呈负相关,但均未达到显著水平。

表3 栓皮栎种子表型性状与生态因子的相关系数
2.4 栓皮栎不同种源种子表型性状的聚类分析结果 以栓皮栎种子长、种子宽、种子长宽比、单果重为变量,基于欧氏距离对15个栓皮栎种源进行聚类分析(图1)。从图1可以看出,供试15个栓皮栎种源可分为3大类,第1类包括北京门头沟、安徽滁州大柳、湖南郴州、河南窑店、北京平谷、陕西太白、河北邢台、安徽滁州皇甫山、云南安宁9个种源;第2类包括山东牟平、河南泌阳、陕西延安、陕西黄龙4个种源;第3类包括陕西镇坪和河南灵宝2个种源。
3 结论与讨论
(1)15个不同种源栓皮栎的种子宽、种子长变异幅度分别为12.50 ~21.34 mm 和 20.17 ~24.30 mm,种子长宽比变异幅度为1.06~1.79,果实平均单果重变异幅度为1.55~4.98 g。栓皮栎不同种源间种子的种子长、种子宽、种子长宽比、单果重等性状差异均达到极显著水平(P<0.01)。一般认为,遗传和环境协同作用导致种源间的差异,栓皮栎不同种源间种子表型性状存在差异性,表明不同种源的遗传差异和环境压力造成的差异,使之产生了不同的表型变异,为接下来开展其优良种源及家系选择以及遗传改良工作提供了基础条件。国内一些科研学者利用种子性状差异已开展了沙拐枣[30]、文冠果[31]、麻疯树[32]、红榉[33]、山核桃[34]、花曲柳[35]、红松[36]、西藏光核桃[37]等优良种源及家系的选育工作,并选育出一批优良种源及家系。
(2)在栓皮栎种子4个表型性状中,种子宽和单果重与种子长宽比呈极显著负相关(P<0.01),其他性状两两间呈正相关,其中种子宽与单果重间呈极显著正相关(P<0.01),说明种子越宽,单果重越大。种子长度与单果重呈显著正相关(P<0.05),而种子其他性状间均未达显著性相关水平(P≥0.05)。对栓皮栎种子性状与种源地的地理、气候等生态因子的相关性分析表明,单果重与经度呈显著正相关(P<0.05),其他生态因子与种子性状相关性不显著(P≥0.05),说明栓皮栎种子性状受到地理、气候等生态因子影响之外,还可能受到遗传信息、栽培方法等因子的共同影响。
(3)聚类分析结果表明,供试栓皮栎15个种源可以划分为3大类,第1大类包括9个种源,第2大类包括4个种源,第3大类包括2个种源。从各大类地理分布的种源来看,聚类分析结果与各种源地理分布无显著相关性。由于表型性状特征在不同环境下差异很大,用种子表型性状的4个变量检测种源的遗传变异具有一定的局限性[38],因而仅基于表型性状变异的聚类分析结果不能整体反映栓皮栎种源内在的遗传差异。
[1]JAIR G G,GUADALUPE W L.Edge effect on acorn removal and oak seedling survival in Mexican lower montane forest fragments[J].New Forests,2006,31:487 -495.
[2]漆民楷,陈红.栓皮栎薪炭林研究[J].四川林业科技,1992,13(1):72 -76.
[3]高根虎,卢从样.陕西省软木工业发展的优势及对策[J].陕西林业科技,2002(1):63-65.
[4]郑志峰.软木资源及其利用[J].云南林业,2005,26(3):23 -24.
[5]谢碧霞,陈训,张东林,等.中国木本淀粉植物[M].北京:科学出版社,2008:33-45.
[6]李迎超,于海燕,付甜,等.中国栓皮栎资源生产燃料乙醇的潜力及其空间分布[J].林业科学,2013(11):129-134.
[7]周建云,郭军战,杨祖山.栓皮栎天然群体过氧化物酶同工酶遗传变异分析[J].西北林学院学报,2003,18(2):33 -36.
[8]武康生.栓皮栎苗木的水分关系[J].北京林业大学学报,1990,12(3):26-33.
[9]谢会成,朱西存.水分胁迫对栓皮栎幼苗生理特性及生长的影响[J].山东林业科技,2004(2):6-7.
[10]吴征镒.中国植被[M].北京:科学出版社,1980:261 -262.
[11]郑万钧.中国树木志:2卷[M].北京:中国林业出版社,1985:23-30.
[12]雷明德.陕西植被[M].北京:科学出版社,1999:163 -164.
[13]张仰渠.陕西森林[M].西安:陕西科学技术出版社,1989:235-240.
[14]傅焕光,于光明.栓皮栎栽培与利用[M].北京:中国林业出版社,1978:7-10.
[15]张存旭,宋敏,赵忠.栓皮栎茎段离体培养的研究[J].西北植物学报,2004,24(7):1260 -1265.
[16]张存旭,姚增玉,赵忠.栓皮栎体胚诱导关键影响因素研究[J].林业科学,2005,41(2):174 -178.
[17]张存旭,张瑞娥,张文辉,等.不同群体栓皮栎栓皮性状变异分析[J].西北林学院学报,2003,18(3):34 -36.
[18]郑均宝,于力,王德艺.影响栓皮栎等造林成活因素的研究[J].河北林果研究,1990,5(1):33 -36.
[19]张理宏.栓皮栎扣杯直播造林技术[J].林业实用技术,1992(4):26-27.
[20]朱志诚.秦岭北麓的栎林[J].陕西林业科技,1979(4):1 -5.
[21]吴明作,姜志林,刘玉萃.栓皮栎种群的年龄结构和稳定性研究[J].河南科学,1999,17(1):69 -73.
[22]吴明作,刘玉萃.栓皮栎种群数量动态的谱分析与稳定性[J].生态学杂志,2000,19(4):23 -26.
[23]吴明作,姜志林.栓皮栎种群的生命进程与稳定性研究[J].南京林业大学学报,1999,23(5):55 -59.
[24]刘彩云.栓皮栎种群结构与动态规律研究[D].南京:南京林业大学,2008:23 -28.
[25]程瑞梅,肖文发.河南宝天曼栓皮栎林群落特征及物种多样性[J].植物资源与环境,1998,7(4):8 -13.
[26]吴明作,杨玉珍.河南省栓皮栎林主要种群的生态位研究[J].西北植物学报,1999,19(3):511 -518.
[27]李安平,谢碧霞,田玉峰.橡实淀粉生料发酵生产燃料酒精工艺研究[J].中国粮油学报,2011(3):91 -94.
[28]李迎超,厉月桥,王利兵,等.木本淀粉能源植物栓皮栎与麻栎的资源调查以及分布规律[J].林业资源管理,2013(20):94-101.
[29]李迎超.木本淀粉能源植物栓皮栎与麻栎资源调查及地理种源变异分析[D].北京:中国林业科学研究院,2013:45 -87.
[30]蔡亮,孙鹤,王菊萍,等.河西走廊不同群体沙拐枣种子表型性状变异研究[J].现代园艺,2013(18):10-11.
[31]侯元凯,黄琳,高巍,等.不同种源文冠果果实及种子表型性状变异的研究[J].中南林业科技大学学报,2013(7):20-24.
[32]惠文凯,陈晓阳,刘明骞.麻风树种源间种实性状变异研究[J].北京林业大学学报,2014(3):110-114.
[33]王旭军,张日清,许忠坤,等.红榉不同种源种子形态性状变异[J].中南林业科技大学学报,2015(1):1-7.
[34]张辉,董雷鸣,曾燕如,等.山核桃天然群体家系种实和幼苗性状变异分析[J].林业科技开发,2014(3):93 -95.
[35]张鹏,沈海龙.花曲柳优树种子性状变异及其子代苗期生长表现[J].林业科技开发,2013(2):18-22.
[36]张振,张含国,周宇,等.红松多无性系群体的种实性状变异研究[J].北京林业大学学报,2015(2):67-78.
[37]周淑香,符亚茹,王超,等.西藏光核桃果实及种子表型性状变异研究[J].北方园艺,2013(23):38 -40.
[38]XU F,GUO W H,XU W H,et al.Habitat effects on leaf morphological plasticity in Quercus acutissima[J].Acta biologica cracoviensia:Series botanica,2008,50(2):19 -26.
