一种新型训练蜜蜂装置的初步探索
2015-12-16狄纳言
狄纳言,高 洁
(1.中国科学院西双版纳热带植物园,昆明 650223;2.中国科学院大学,北京 100049)
虽然蜜蜂Apis 的脑部结构简单,但是却具有比较强的学习及记忆能力。例如,意大利蜜蜂Apis mellifera 通过学习记忆能够识别花粉与花蜜的味道或者颜色,形成永久记忆(Kandel,2001)。蜜蜂的嗅觉记忆是研究嗅觉的经典模型,嗅觉在蜜蜂学习、觅食过程中起重要作用(Breed,1983),意大利蜜蜂拥有163个嗅觉受体基因(Robertson and Wanner,2006;Weinstock et al.,2006),具有很强的嗅觉记忆能力(Laska et al.,1999)。蜜蜂的正常活动半径为2-3 km,通过学习和记忆飞行途径的地形、地貌,能从巢外复杂的环境中回到蜂巢(Pahl et al.,2011)。蜜蜂作为研究学习记忆行为的重要模式昆虫,针对蜜蜂的学习与记忆机制已经被各国各科研工作者用于研究(Menzel,2001),已被作为研究动物学习与记忆的神经基础的标准模型(Hammer and Menzel,1998;郑火青和胡福良2009;Gronenberg and Couvillon,2010)。
在研究蜜蜂的学习和记忆过程中,通常需要训练蜜蜂。早在1986年Getz 等人依据巴甫洛夫条件反射原理,在Bitterman 等人的基础上(1983),建立条件刺激(CS)结合非条件刺激(US)的蜜蜂模式(Getz et al.,1986),训练方法为:用气体A(CS+,即conditioned stimulus)对着蜜蜂触角吹3 s,若蜜蜂伸吻则给予一滴糖浆(US+,即unconditioned stimulus)。蜜蜂均受训一次后,用另一种气体B(CS-)对着蜜蜂触角吹3 s,若蜜蜂对气体B 无反应则给予一滴氯化钠(US-)溶液。如果气体A 与气体B 对比足够大,则可以实现训练工蜂对气体A 有反应而对气体B 无反应。其中,条件刺激(CS)和非条件刺激(US)控制是关键,关系训练效果。
本文提出利用软件控制条件刺激和非条件刺激的时间、强度(流量)等参数,研制了一轮可训练10 头蜜蜂的自动训练仪,实现了对训练过程的精确控制,为蜜蜂学习记忆行为提供一种更为便捷、高效、精确的方法。
1 材料与方法
蜜蜂训练装置的主要由刺激系统和蜜蜂旋转训练台、控制器、红外检测器以及计算机软件组成。基于计算机控制给予条件刺激(气味)的气压系统、非条件刺激(糖浆)的机械系统和变换受训蜜蜂位置的旋转台。气压系统包括:气瓶、气泵、气管和流量计;机械系统包括:电机、注射器、饲喂盘和液管。整个装置的实物图和示意图如下:
刺激系统,即条件刺激部分与非条件刺激部分结构如图3 所示,条件刺激利用气压系统传输:气泵与流量计相连,流量计控制气流大小;控制器与旋转训练台上的出气孔相连,通过电磁阀的开闭控制出气时间以及出气孔(A 孔或B 孔),同时训练气与风扇相连,通过控制器控制风扇抽气,确保训练前、后旋转训练台上没有残留气味,影响下一只蜜蜂受训。由计算机软件控制气体的开关信号,并通过USB7408N 连接控制器软件上可设定气体通道、气体时间、以及抽气时间等训练参数。非条件刺激利用液压传输系统,由计算机软件提供糖浆喂食信号,通过USB7408N 连接控制器控制喂食电机运动并同步联动糖浆电机,将无菌注射器中的蔗糖糖浆通过糖浆管运输到旋转训练台上的糖浆滴管,通过控制台控制糖浆时间以及糖浆流速,糖浆从滴管滴落在糖浆盘上,供蜜蜂取食,以此达到自动喂食蜜蜂的效果。

图1 蜜蜂训练装置实物图Fig.1 The stimulation apparatus for bee trained

图2 蜜蜂训练装置示意图Fig.2 The diagram of stimulation apparatus for bee trained
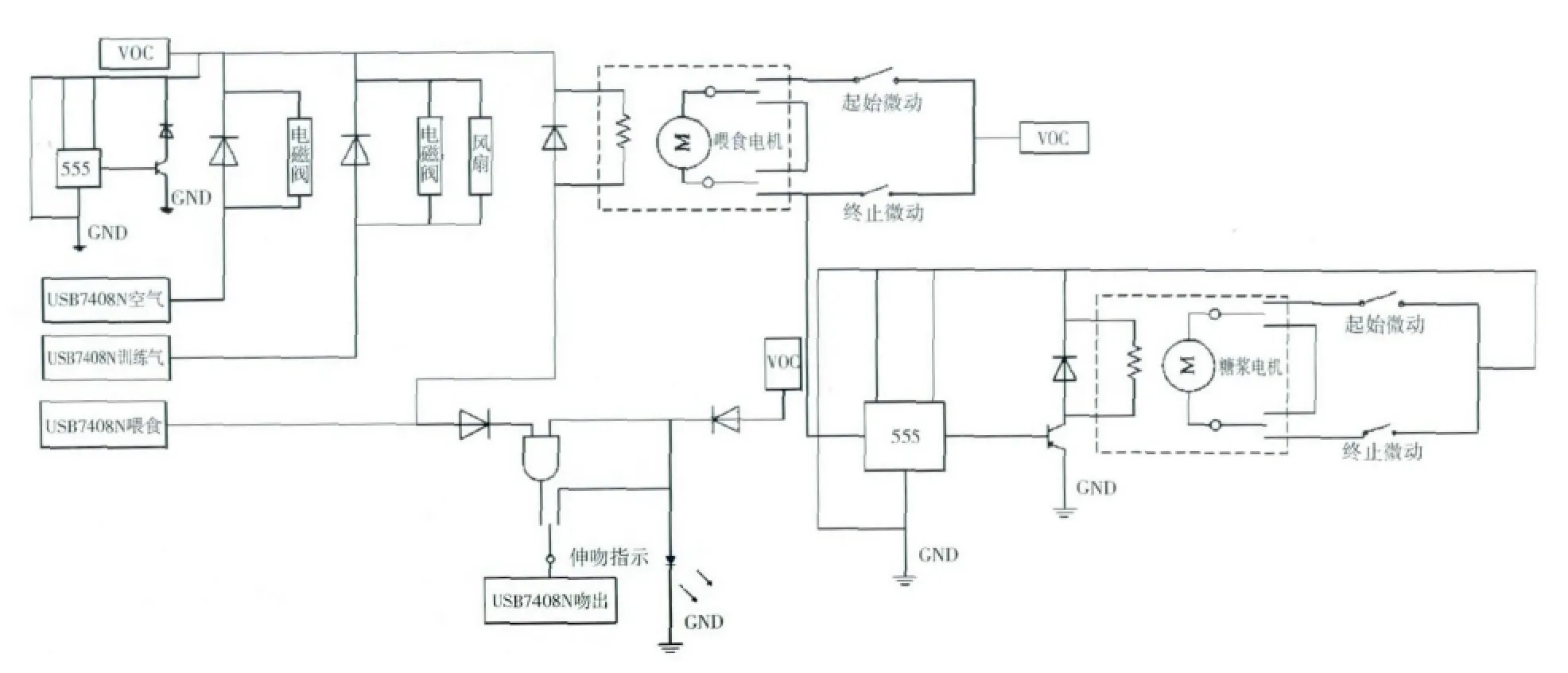
图3 蜜蜂训练装置条件刺激与非条件刺激部分电路图Fig.3 The circuit diagram that indicates the conditioned stimulus and the unconditioned stimulus
2 结果与分析
2.1 训练蜜蜂准备
选择一群蜜蜂,在蜂巢门口,于8∶00-10∶00拦截出巢工蜂15-20 头,将其放入小玻璃瓶内(每个小玻璃瓶内放入1-2 头),4℃冰水混合物中麻痹至蜜蜂无活动后,取出将其固定在专用的金属支架上,只留有头部和触角可自由活动。待蜜蜂苏醒后,用糖浆轻触蜜蜂的触角,诱导蜜蜂伸吻后让蜜蜂吸食糖浆,直至吃饱。把喂饱的蜜蜂放入暗室2 h(温度25℃、湿度85%)后取出蜜蜂,糖浆轻触蜜蜂的触角检测是否伸吻,如果伸吻,表明该蜜蜂具有良好的反应能力,可用于训练。每次选择10 头放到旋转训练台上用于训练。
2.2 蜜蜂训练方法
采用目标气体和空白(空气)训练重复的方法训练蜜蜂。所用供试蜜蜂随机分为2 组,一组接受手动训练,另一组接受仪器自动训练,训练操作方法如下:
2.2.1 自动训练方法
目标气体训练:在黑暗通风厨内,首先打开自动训练装置上的红光灯罩,然后打开Bee Training 程序,选择气体通道并设定训练前等待时间、给气时间、给糖时间,以及抽气时间等参数后,仪器将自动将1 头蜜蜂旋转到训练位置(即同时施于条件刺激和非条件刺激位置),后停留40 s 让其熟悉训练台,接收条件刺激(CS)5 s(气体流速33 mL/s),在条件刺激的第3 秒,给予非条件刺激3 s(溢出糖浆20μL),让蜜蜂吸吮糖浆,然后让蜜蜂在训练位置再停留15 s,下待训1 头蜜蜂自动旋转到训练位置,重复以上训练。单只蜜蜂的训练时间为1 min,10 头蜜蜂完成训练的时间为10 min。
空白气体训练方法:当所有蜜蜂都经过一次训练后,所有训练参数不变,但条件刺激的气体为空气,让所有蜜蜂都接受一次仅有空气流的空白训练,这个过程是为了消除气流对蜜蜂的机械刺激。
每次让10 头蜜蜂接受3 次目标气味和3 次空白气流的刺激训练。训练好的蜜蜂喂饱后放入暗室,待用。
2.2.2 手动训练方法
手动训练参数同上,由训练人员手动给予蜜蜂条件和非条件刺激。
2.3 训练效果检测
2 h 后从暗室拿出蜜蜂,用糖浆检测是否伸吻,淘汰没有伸吻的蜜蜂。将10 头蜜蜂放入旋转控制台,软件设定仅给气,不给糖浆。用目标气味刺激这些蜜蜂,观察其是否伸吻,再用空气刺激确定其是否伸吻,如果对目标气味刺激伸吻且对空白气流刺激不伸吻,表明这些蜜蜂已学会并记住了该气味,记录学会并记住气味的蜜蜂数量。
2.4 数据分析
利用上述2种方法,每次训练50 头蜜蜂,重复训练6 次,共训练600 多头蜜蜂。统计每次50 头中学会和记住目标气味的蜜蜂数量N,计算训练的成功率:P=N/50×100%。之后得到6 次训练成功率的平均值。

图4 蜜蜂训练装置与手动训练成功率比较Fig.4 The success rate of apparatus and manually operated for training bees
使用蜜蜂训练装置自动训练蜜蜂数共301 头;手动训练蜜蜂数共304 头,蜜蜂训练装置的平均成功率为58.47%,手动训练蜜蜂的平均成功率为49.67%。试验结果使用SPSS19.0 进行χ2检验,结果表明:使用蜜蜂训练装置,训练成功率显著高于手动训练(χ2=9.822,P=0.002),说明蜜蜂训练装置对蜜蜂的训练效果优于手动训练。
3 结论与讨论
蜜蜂训练装置作为一种新型的训练蜜蜂仪器,具有更为精准、快捷的优势。蜜蜂训练装置的优势表现为:
首先,通过计算机软件控制条件和非条件刺激,能控制施放刺激方式,单独或同时,以及刺激蜜蜂的时间和强度,让受训蜜蜂建立有效的条件反射。并且位于糖滴管下方红外检测器自动记录蜜蜂展吻问数,如是否展吻和展吻次数,不仅实现了训练、检测的自动化,并且训练结果自动保存,极大提高训练效率。
其次,蜜蜂训练装置可对气体通道自动选择,通过软件控制2个气瓶阀门开关。使用空气消除蜜蜂对气流、位置的感知,控制给气量可以降低试验误差,避免因给气量的不同而导致蜜蜂记忆差异,让蜜蜂更快建立“气味—奖励”这一条件反射。
再次,蜜蜂训练装置缩短训练时间,蜜蜂训练装置1 min 针对1 头蜜蜂进行训练,一轮10 头蜜蜂训练一次需10 min,提高训练效率。
此外,旋转训练台可加载LED 红光灯罩,消除视觉对蜜蜂的影响,减少环境对训练结果的影响。在下一步工作中,将增加自动刺激触角装置,在器件的选择、电路的设计上进行改进,特别要注意非条件刺激控制部分的改进,提高训练蜜蜂的精度和成功率。
References)
Bitterman ME,Menzel R,Fietz A,et al.Classical conditioning of proboscis extension in honeybees(Apis mellifera)[J].Journal of Comparative Psychology,1983,97(2):107.
Breed MD.Nest mate recognition in honeybees[J].Animal Behavior,1983,31(1):86-91.
Getz WM,Brückner D,Smith KB.Conditioning honeybees to discriminate between heritable odors from full and half sisters[J].Journal of Comparative Physiology A,1986,159(2):251-256.
Gronenberg W,Couvillon MJ.Brain composition and olfactory learning in honey bees[J].Neurobiology of Learning and Memory,2010,93(3):435-443.
Hammer M,Menzel R.Multiple sites of associative odor learning as revealed by local brain microinjections of octopamine in honeybees[J].Learning & Memory,1998,5(1):146-156.
Kandel ER.The molecular biology of memory storage:a dialogue between genes and synapses[J].Science,2001,294(5544):1030-1038.
Laska M,Galizia CG,Giurfa M,et al.Olfactory discrimination ability and odor structure-activity relationships in honeybees[J].Chemical Senses,1999,24(4):429-438.
Menzel R.Searching for the memory trace in a mini-brain,the honeybee[J].Learning & Memory,2001,8(2):53-62.
Pahl M,Zhu H,Tautz J,et al.Large scale homing in honeybees[J].PLoS One,2011,6(5):e19669.
Robertson HM,Wanner KW.The chemoreceptor superfamily in the honey bee,Apis mellifera:expansion of the odorant,but not gustatory,receptor family[J].Genome Research,2006,16(11):1395-1403.
Weinstock GM,Robinson GE,Gibbs RA,et al.Insights into social insects from the genome of the honeybee Apis mellifera[J].Nature,2006,443(7114):931-949.
Zheng HQ,Hu FL.Honeybee:a newly emerged model organism[J].Acta Entomologica Sinica,2009,52(2):210-215.[郑火青,胡福良.蜜蜂:新兴的模式生物[J].昆虫学报,2009,52(2):210-215]
