Neuronal gene transcription modulates demyelination and remyelination in a mouse model of multiple sclerosis
2015-12-15SofiaAnastasiadou,BerndKnll
Neuronal gene transcription modulates demyelination and remyelination in a mouse model of multiple sclerosis
The pathological hallmark of MS is the occurrence of multifocal demyelinated lesions in the brain and spinal cord of patients. At early stages of the disease, active lesions are associated with infl ammation, de- and remyelination. As disease progresses, remyelination fails and axons devoid of myelin eventually degenerate. Clinical symptoms are mainly caused by the ongoing neurodegeneration of demyelinated axons in the grey and white matter tracts (Ellwardt and Zipp, 2014). Hence, current therapeutic approaches do not only concentrate on hindering demyelination, but also on promoting oligodendrocyte precursor cell (OPC) maturation and remyelination.
Infl ammatory processes play a key role in disease pathogenesis, thus, MS is commonly considered to be caused by an autoimmune response. For example, breakdown of the blood-brain barrier, autoreactive T cells, monocyte infi ltration and autoantibodies are commonly found in patients suff ering from MS (Yadav et al., 2015). Beside the components of the peripheral immune system, activation of brain resident infl ammatory cells (i.e., astrocytes and microglia) is a classical feature of MS and infl uences disease progression considerably (Claycomb et al., 2013).
Astrocytes are the most abundant cell type in the CNS with functions ranging from building the blood-brain barrier to establishing a scaff old for migrating neurons during brain development. One of the most intriguing functions of astrocytes is the regulation of the adaptive and innate immune system in the CNS. Activated astrocytes in lesions of MS patients can infl uence the activation and infi ltration of T cells into demyelinated lesions for example by controlling the permeability of the blood-brain barrier, by presenting antigens or by secreting pro-infl ammatory chemokines. Additionally, an essential function of astrocytes is the activation of microglia (Claycomb et al., 2013; Skripuletz et al., 2013).
Myelin and cell debris within MS lesions are mainly cleared away by microglia, the primary phagocytic cells in the CNS. Microglia activity can be regulated by astrocytes either by direct cell-cell contacts, or through chemokine secretion. Depending on the mode of activation, microglia can develop into pro-infl ammatory M1 cells promoting further infl ammation and eventually leading to neurotoxicity. On the other hand, microglia can polarize to anti-infl ammatory M2 cells, which have neuroprotective functions and promote maturation of OPCs. Interestingly, M2 microglia are only present in active MS lesions and at the border of chronic lesions, where remyelination is still ongoing (Ellwardt and Zipp, 2014). This possibly indicates an essential role of M2 microglia in promoting remyelination. Taken together, the exact regulation of astrocyte and microglia activation seems to be critical for keeping the balance between neuroprotection and neurotoxicity in pathological situations like MS.
In addition to the demyelination and neuroinflammation, axonal degeneration is a typical characteristic of MS lesions. Neuronal pathology like axonal swellings, Wallerian degeneration and axonal transections have been repeatedly described in MS patients and MS mouse models and are commonly accepted to be secondary to axonal demyelination and infl ammation (Trapp and Nave, 2008). Recent research has increasingly focused on the role of the neuronal compartment in MS development and progression. Interestingly, studies in MS patients with relapsing remitting MS (RR-MS) receiving immunomodulatory drugs have shown that while suppression of the immune system alleviates clinical symptoms during relapses, it does not slow down disease progression (Shirani et al., 2012). Additionally, experiments with mice lacking the oligodendroglial protein 2′,3′-cyclic nucleotide phosphodiesterase (CNPase) have shown axonal pathology without myelin impairment (Lappe-Siefke et al., 2003). Those fi ndings indicate that the etiology of neurodegeneration cannot solely be explained by demyelination or neuroinfl ammation. In sum, recent research has identifi ed the neuronal compartment and axon-glia interactions as key players in MS development. Hence, it is considerable that demyelination and neuroinfl ammation are (at least in some cases) secondary to neuronal damage.
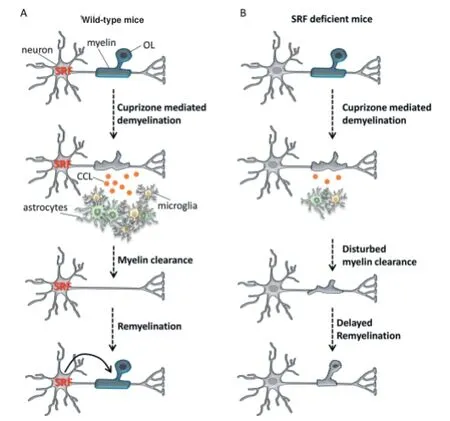
Figure 1 Model for the regulation of infl ammation, demyelination and remyelination by serum response factor (SRF).
Paracrine regulation demyelination and remyelination by neuronal serum response factor (SRF) gene transcription: In our laboratory, we have focused on the role of the transcription factor SRF, which is highly expressed in neurons. SRF regulates the transcription of immediate early genes (IEGs), cytoskeletal genes and genes related to actin dynamics (Knöll and Nordheim,
2009). Thereby, it can control diverse neuronal processes in the developing brain, including neuronal migration, axon guidance and regeneration and growth cone dynamics. Besides those neuron specifi c processes, we analyzed the role of SRF in myelin development in mice. We demonstrated that mice lacking SRF in forebrain neurons only, fail to generate myelin upon birth. Further, we identifi ed a mechanism by which neurons regulate the maturation of pre-myelinating oligodendrocytes in a paracrine fashion (Stritt et al., 2009).
采空区垂直投影的平面范围界线根据调查资料、物探解译资料与验证资料综合分析适当外推,圈定采空区垂直投影总面积约4341m2,采空区容积约19793m3,见图2。
In our recent study, we aimed at analyzing the role of SRF in demyelination and remyelination in young adult mice with an already established myelinated neuronal circuit. Our data show that the deletion of Srf in neurons of adult mice can infl uence demyelination and infl ammation in the cuprizone mouse MS model of toxic demyelination. In fact, whereas wild-type mice had a robust demyelination upon cuprizone treatment, the lack of SRF in forebrain neurons resulted in higher amounts of non-removed damaged myelin. Since oligodendroglia (OLs) of Srf defi cient mice where similarly depleted by cuprizone as OLs of wildtype mice, we concluded that Srf mutant mice had problems effi ciently clearing away the remaining myelin debris (Figure 1; Anastasiadou et al., 2015). Moreover, we demonstrated that SRF depletion in neurons results in disturbed induction of brain resident infl ammatory cells (i.e., microglia and astrocytes). Previous reports have already shown that astrocytes and microglia play an essential role in removing cell and myelin debris. Therefore, this defective induction of infl ammation is a compelling explanation for the impaired myelin debris clearance observed in Srf mutant mice. In our mouse model, the deletion of Srf was restricted to neurons, indicating the involvement of a paracrine mechanism. Hence, our results support the importance of the crosstalk between OLs, neurons, astrocytes and microglia for the regulation of demyelination and infl ammation in pathological conditions. Interestingly, in contrast to neuronal SRF depletion, the deletion of Srf only in OLs did not infl uence demyelination or infl ammation upon cuprizone treatment. This indicates that mainly neurons (but not OLs) use a SRF mediated paracrine pathway for the induction of brain resident immune cells.
Besides demyelination and infl ammation, a major issue in MS pathology is the compromised remyelination. Hitherto, it is not known why OPCs eventually stop diff erentiating, resulting in remyelination failure. Considering that SRF can regulate OPC diff erentiation during development (Stritt et al., 2009), we investigated whether SRF also infl uences OL function during remyelination. In wild-type mice, we detected rapid generation and maturation of OPCs during remyelination. In line, the expression of myelin genes, which had been strongly reduced during demyelination, showed an overshooting reaction reaching even higher levels than untreated control mice. In contrast, generation and maturation of OPCs during remyelination were delayed in mice lacking SRF in forebrain neurons. Additionally, the expression of genes encoding for myelin proteins was reduced in those mice (Anastasiadou et al., 2015). Hence, we demonstrated that neuronal SRF can influence OL function during remyelination in a paracrine fashion.
In order to shed some light on the interaction of the involved cell types, we searched for mediators of the neuron-glia crosstalk. In this search, we considered molecules that fulfi ll two criteria: i) they can be secreted by neurons and ii) they are related to infl ammation. By gene expression analysis, we identifi ed a group of chemokines, the CC chemokine ligands (CCLs), which are highly induced during demyelination. Particularly CCL2, CCL3 and CCL5 show reduced expression in SRF defi cient mice (Anastasiadou et al., 2015). CCLs have previously been reported to activate and attract astrocytes and microglia and are also known to be secreted by neurons. Therefore, we suggest that SRF activity in neurons can modify the expression of CCLs during demyelination, thereby infl uencing the communication of neurons with astrocytes and microglia (Figure 1). Nevertheless, the contribution of diff erent CCLs to the neuron-glia crosstalk still has to be elucidated e.g., by the use of CCL2, CCL3 or CCL5 defi cient mice.
Taken together, our results underline the importance of a well-orchestrated communication between neurons and diff erent types of glia cells for the regulation of essential processes in pathological demyelination and in remyelination. Our observations suggest that neurons are actively involved in activation/recruiting of brain resident infl ammatory cells, thereby regulating myelin debris clearance. Additionally, our data emphasize the delicate balance of infl ammation leading either to neurodegeneration or neuroprotection. Current therapeutic strategies for MS aim at suppressing CNS infl ammation and enhancing neuroprotection. Therefore, molecular and cellular insights obtained in rodent MS models, highlighting the importance of the neuronal compartment for disease progression, might prove useful for the understanding of mechanisms involved in the onset of human MS disease. Finally, the investigation of neuronal signaling mechanisms, such as SRF mediated gene transcription, identifi ed in MS mouse models, might provide new valuable molecular targets relevant to human MS therapy.
Sofi a Anastasiadou*, Bernd Knöll
Institute of Physiological Chemistry, Ulm University, Ulm, Germany
*Correspondence to: Sofi a Anastasiadou,
sofi a.anastasiadou@uni-ulm.de.
Accepted: 2015-08-18
orcid: 0000-0001-9459-4409 (Sofi a Anastasiadou)
Anastasiadou S, Liebenehm S, Sinske D, Meyer zu Reckendorf C, Moepps B, Nordheim A, Knöll B (2015) Neuronal expression of the transcription factor serum response factor modulates myelination in a mouse multiple sclerosis model. Glia 63:958-976.
Claycomb KI, Johnson KM, Winokur PN, Sacino AV, Crocker SJ (2013) Astrocyte regulation of cns infl ammation and remyelination. Brain Sci 3:1109-1127.
Ellwardt E, Zipp F (2014) Molecular mechanisms linking neuroinfl ammation and neurodegeneration in MS. Exp Neurol 262:8-17.
Knöll B, Nordheim A (2009) Functional versatility of transcription factors in the nervous system: the SRF paradigm. Trends Neurosci 32:432-442.
Lappe-Siefke C, Goebbels S, Gravel M, Nicksch E, Lee J, Braun PE, Griffi ths IR, Nave KA (2003) Disruption of Cnp1 uncouples oligodendroglial functions in axonal support and myelination. Nat Genet 33:366-374.
Shirani A, Zhao Y, Karim ME, Evans C, Kingwell E, van der Kop ML, Oger J, Gustafson P, Petkau J, Tremlett H (2012) Association between use of interferon beta and progression of disability in patients with relapsing-remitting multiple sclerosis. JAMA 308:247-256.
Skripuletz T, Hackstette D, Bauer K, Gudi V, Pul R, Voss E, Berger K, Kipp M, Baumgärtner W, Stangel M (2013) Astrocytes regulate myelin clearance through recruitment of microglia during cuprizone-induced demyelination. Brain 136:147-167.
Stritt C, Stern S, Harting K, Manke T, Sinske D, Schwarz H, Vingron M, Nordheim A, Knöll B (2009) Paracrine control of oligodendrocyte diff erentiation by SRF-directed neuronal gene expression. Nat Neurosci 12:418-427.
Trapp BD, Nave KA (2008) Multiple sclerosis: an immune or neurodegenerative disorder? Annu Rev Neurosci 31:247-269.
Yadav SK, Mindur JE, Ito K, Dhib-Jalbut S (2015) Advances in the immunopathogenesis of multiple sclerosis. Curr Opin Neurol 28:206-219.
10.4103/1673-5374.165507 http://www.nrronline.org/
Anastasiadou S, Knöll B (2015) Neuronal gene transcription modulates demyelination and remyelination in a mouse model of multiple sclerosis. Neural Regen Res 10(9):1401-1402.
猜你喜欢
杂志排行

中国神经再生研究(英文版)的其它文章
- Lactulose enhances neuroplasticity to improve cognitive function in early hepatic encephalopathy
- Elastic modulus aff ects the growth and diff erentiation of neural stem cells
- Optimal concentration and time window for proliferation and diff erentiation of neural stem cells from embryonic cerebral cortex: 5% oxygen preconditioning for 72 hours
- Stem Cell Ophthalmology Treatment Study (SCOTS) for retinal and optic nerve diseases: a case report of improvement in relapsing auto-immune optic neuropathy
- Repair of peripheral nerve defects with chemically extracted acellular nerve allografts loaded with neurotrophic factors-transfected bone marrow mesenchymal stem cells
- Polylactic-co-glycolic acid microspheres containing three neurotrophic factors promote sciatic nerve repair after injury