影响鱼、虾、贝类等海洋生物氮、磷代谢的主要因素分析
2015-12-14吴丰辉阎波田胜艳
吴丰辉,阎波,田胜艳
(天津科技大学海洋工程与科学学院 天津市海洋资源与化学重点实验室 天津市 300457)
排泄是动物进行能量代谢的基本生理活动之一,能反映动物的正常生理状态,其排泄物成分复杂,主要包括溶解态的氮和磷,其中氨氮占70%以上。在海洋环境中,无脊椎动物的N、P排泄对于海洋生态系统的能量流动和物质循环起到非常重要的作用,特别是在养殖海区,可能对整个海区的生态动力学都会产生较大的影响。同时氮和无机磷是浮游植物的重要营养物质,海洋动物N、P 排泄对浮游植物的生产具有重要意义,特别是再循环无机形态的N(NH4+-N)和P(PO43--P)作为浮游植物可以直接利用的营养元素形态,对研究其在浮游植物和海洋动物之间的循环利用具有重要意义。
在自然环境中,各种自然因素(如温度、盐度、光照、摄食条件)以及人为因素(如投放饲养添加剂)都会对海洋生物的摄食、代谢等生理活动产生一定的影响,这里对影响海洋养殖动物,尤其是鱼、虾、贝类氮、磷代谢的主要因素进行分析,探讨代谢物与环境之间的相互关系,以期为进一步研究海洋生物在氮、磷再循环过程中的作用,减少养殖污染,保护海洋环境提供科学依据。
1 海洋生物氮、磷代谢的主要形式
海洋环境中的氮与磷是重要的生源要素[1],也是海洋动物排泄物的主要成分。对于硬骨鱼类来说,氨和尿素是最主要的排泄产物,同时还有极少量的肌酸、尿酸、嘌呤、氨基酸等物质。在淡水硬骨鱼类的N 排泄物中,氨氮占排泄总量的80%;而一些海洋性鱼类趋于排泄更大比率的其它代谢产物,如尿素、肌酸、嘌呤等其它产物,比例可达到31%-40%[2]。双壳贝类的N 排泄物包括氨、尿素、尿酸、氨基酸等,其中氨的比例最大,达到或超过70%,其余部分因种类不同而不等,同时氨基酸也占据着相当可观的比例,可达5%-50%,但氨基酸的含量与季节有关,并与饥饿条件或配子发生有关。与双壳贝类相类似,在浅海和河口的甲壳动物的N 排泄中,氨占总可溶性氮(TDN)排泄的60%-95%,氨基酸占2%-25%,尿素占1%-5%,但几乎没有尿酸。一些对虾则会排泄一定数量的硝酸盐[3]。
磷存在于海洋生物的所有细胞中,作为核酸和磷脂的重要组分,磷直接参与所有产生能量的细胞内反应,保证细胞膜的结构完整,并维持各种细胞功能。对于养殖的鱼、贝、虾类海洋动物而言,饵料中的磷是促进其生长的主要磷源。被吸收到生物体内的磷只有不到30%能够被生物体同化,而超过70%的磷将随着粪尿被排出体外。日本在网箱养殖鲤鱼方面的研究表明:进入水体的磷超过了投入量的70%,其中5.1%随尿排出体外、65.1%随粪便排出,而这些被排出体外的磷只有24.1%是溶解在水里,其余部分则沉积于底泥中[4]。Smaal 等研究发现贻贝的P 排泄中,有机磷占40%[5]。同样,Atkinson 等[6]观察到底栖无脊椎动物(包括双壳贝类)的P 排泄中,有机磷占总磷排泄的29%。
2 影响鱼、虾、贝类等海洋生物氮、磷代谢的主要因素
在自然环境中,影响海洋生物氮、磷代谢的因素很多,包括生物体自身因素:如种类、性别、体长、体重、饥饿状态等;环境因素如:水温、盐度、光照、水体pH 值、溶解氧水平、水体氨氮含量等;人工养殖因素如:摄食条件、饲料添加剂等,下面对除生物体自身因素外的其它影响因素分别进行分析:
2.1 摄食对海洋生物氮、磷代谢的影响
根据氮、磷元素来源的不同,海洋生物的氮、磷排泄可分为内源性和外源性。前者指非食物来源,由机体组织代谢产生的氮、磷排泄物,后者指来自食物源的氮、磷排泄物。通常内源性排泄的量很少,在进行氮、磷排泄研究时,可以将目标生物饥饿48 h 后测定其氮、磷排泄率作为内源氮、磷排泄率。摄食组的海洋生物在摄食后氮、磷代谢率会明显提高,这一增加的部分称外源氮、磷排泄率,摄食引起的代谢增加称为特殊动力作用,也称为体增热,表现为体热的增加,是在摄入的食物转化为体内可利用的能源物质过程中的能量消耗。外源氮、磷排泄率主要受到食物的营养组成、日粮水平和投饲方式等的影响。由于特殊动力作用主要是由蛋白质代谢引起的,食物中的蛋白质含量对体增热具有相当大的影响,摄食高蛋白质食物的代谢率大于摄食低蛋白质食物的代谢率。
有研究表明摄食量对外源氮中氨氮的含量影响显著,Cui 和Wootton 等[7]提出:氨排泄水平随摄食水平的升高而升高,二者之间的定量关系可用下列方程表达:

其中:N 为氮排泄率,C 为氮吸收率,a、b 为常数。当氮吸收为0 时,a 值则是其内源性排泄率。
除摄食量外,食物中用做能量的蛋白质比例、蛋白质质量(不同氨基酸配比)、可替代蛋白质的碳水化合物、脂肪和维生素的含量等也影响氨氮排泄率。Brett 和Zala[8]发现红大麻哈鱼(Oncorhynchus nerka)的尿素排泄水平在饥饿和摄食情况下都恒定。Kaushik 等[9]也观察到白斑狗鱼(Esox lucius)的尿素排泄水平并没有出现与氨氮排泄相同的摄食后的峰值。他们的研究进一步证实摄食后排氮的增长部分是氨氮的增长,而尿素的排泄很少受食物消化的影响。
氮、磷两种元素经摄食后,由于其对海洋生物的生理作用不同,代谢率也会有所差别。李松青等[10]研究了摄食对凡纳滨对虾(Litopenaeus vannamei)、又称南美白对虾(White Prawn)氮磷代谢率的影响,其使用的饲料中氮含量为6.18%,磷含量为1.20%,而经摄食后,凡纳滨对虾粪便中氮含量仅为4.05%,而磷含量则为3.54%,说明对虾对饲料中磷的消化率低于氮。凡纳滨对虾摄食饲料后氮、磷排泄率的比值(N/P)显著提高,其外源氮、磷比值是内源氮、磷比值的2.5倍,这除了是由于对虾的氮代谢和磷代谢生理不同外,与对虾对饲料中磷的消化吸收率低于氮的消化吸收率亦有关系。
2.2 温度对海洋生物氮、磷代谢的影响
一般情况下,温度对海洋生物氮、磷代谢的影响极显著。随着温度的上升,海洋生物磷、亚硝酸盐氮、硝酸盐氮、氨氮的代谢率都有所提高[11],这说明随着温度的升高,海洋生物组织器官的活动性能提高,体内的各种生化反应速度加快,致使呼吸和代谢加快,对能量的消耗增大[12]。Cui和Wootton 提出温度对氮排泄的影响为线性关系[7],其相关方程为:
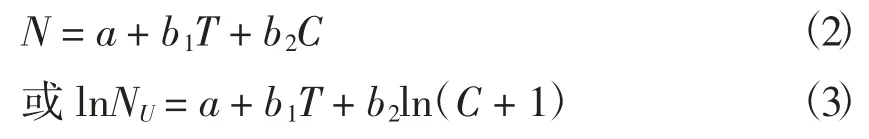
其中:N、Nu 分别表示氨氮和尿素氮的排泄率(mg/d),T 为温度(℃),C 为摄食率(mg/d),a、b1和b2均为常数。
2.3 盐度对海洋生物氮、磷代谢的影响
养殖海水盐度对海洋生物,特别是渗透压调节类动物的生理状况有直接的影响。当外界盐度发生变化时,海洋生物通过改变代谢状况,借助于离子调节和细胞内游离氨基酸库的启动或关闭,来调节体内的渗透浓度,实现有机体与海水之间的水交换以及细胞和组织之间的水分平衡,以适应不同的盐度环境。随着盐度的增加,大部分海洋生物磷、亚硝酸氮的代谢率均提高。硝酸氮、氨氮的代谢率随着盐度的增加而下降[12]。Needham[13]和众多研究者证实甲壳类动物氨氮排泄率是随着盐度的降低而升高的。Pressley 等[14]指出,动物在低盐度中会导致钠离子流失,为了补偿这种流失必须激活钠离子的吸收过程,伴随氨氮排泄的升高。盐度的升高对氨氮排泄的影响则比较复杂,动物体由氨排泄类型转变为尿素排泄类型,氨氮排泄率下降。有研究表明,部分虾类在体液达到等渗点时,其耗氧率最小,此时代谢所需要的能量也是最少的,因而有利于提高摄食能量的利用率[15]。
2.4 光照对海洋生物氮、磷代谢的影响
Dall W 等[16]对虎虾(Tiger Prawns)和食用对虾(Penaeus esculentus Haswell)的研究和姜祖辉[17]对菲律宾蛤仔的研究均表明,这两类动物在黑暗条件下的氮、磷代谢率高于光照条件,这与其生态习性是一致的,虾和双壳贝类等底栖生物终生埋栖生活于海底,适应于在光线较暗的环境中生活,在黑暗的条件下它们的各种活动(如腹足伸出、移动位置等)加强,加快了体内的各种代谢,过强的光照则会抑制其正常的生理活动,导致氮、磷排泄率的降低。
2.5 pH 值对海洋生物氮、磷代谢的影响
pH 值即水体的酸碱度,可直接影响到水中各种生物,包括各种微生物的生长繁殖,调节水体pH 值在水产动物生长最理想的范围内,是水产养殖获得高产稳产的必要条件之一。当水体pH 值过高时,海洋生物对于氨氮的分解代谢增加,会使水中的氨氮含量增加,使水体恶化,不利于海洋生物的生长。而当水体pH 值过低时,海洋生物血液中pH 值会随之下降,血液中氧的分压降低,造成运输氧气的蛋白质功能障碍,使水产动物组织细胞内缺氧,造成呼吸困难,吃食量减少,严重时引起死亡。同时,pH 值的高低与氮磷的排泄也存在一定的联系。随着pH 值的增加,海洋生物磷的代谢率明显下降,氨氮、硝酸氮的代谢率随着pH 值的增加而升高,对亚硝酸氮代谢率的影响不显著[15]。对于虾类养殖来说,虾池水正常pH 范围通常为7.5-8.5,最理想范围为8.0-8.5。
2.6 溶解氧水平对海洋生物氮、磷代谢的影响
溶解氧是鱼、虾、贝类等养殖生态环境中的重要指标之一,它直接或间接地影响着海洋生物的生长。McGraw 等[18]研究发现,在15%-65%的溶解氧饱和度范围内,凡纳滨对虾生长和存活率与溶解氧饱和度呈正相关的关系。崔莹等[19]研究发现,凡纳滨对虾(3.14 cm-3.27 cm)的瞬时耗氧速率随溶氧量的降低而增大。而水体溶解氧水平与水温和水质状况等有一定的相关关系。
2.7 水体氨氮含量对海洋生物氮、磷的影响
海洋环境质量对海洋生物氮、磷排泄量会产生一定程度的影响。而海水中氨氮水平对海洋生物氮、磷排泄的影响较大。赖晓芳等[20]设计了氨氮浓度变化的急变组和缓变组,研究了氨氮浓度对脊尾白虾氮、磷代谢的影响,研究表明因氨分子具有相当高的脂溶性,能穿透细胞膜毒害组织,虾池中氨的积累会增加虾的蜕皮次数,减缓生长,增加耗氧量。当NH4Cl 浓度在0-2.0 mg/L时,脊尾白虾的氮、磷排泄率均随着氨态氮浓度的增加而下降,当NH4Cl 浓度达到2.5 mg/L 时,脊尾白虾的氮、磷排泄率显著降低。而Lin 和Chen[21]研究发现,总氨氮和非离子氨氮对凡纳滨对虾的安全浓度分别为3.55 mg/L 和0.16 mg/L。
2.8 饲料添加剂对海洋生物氮、磷代谢的影响
在进行鱼、虾、贝类等的人工养殖时,通常会采用植物性饲料,植物性饲料中的磷主要是以植酸及其盐的形式存在,难以被生物所利用。已有研究表明[22],向含有植物性原料的鱼类饲料中直接添加适量植酸酶或添加预处理后的植酸酶,能提高鱼类的生长性能。同时添加植酸酶还可以提高鱼类对氮、磷的利用率。一方面,由于植酸酶能催化植物性原料中的植酸,使其水解成无机磷酸(或者磷酸盐)与磷酸肌醇,提高鱼类对植物磷的吸收利用率,降低磷的排泄量;另一方面,植酸酶可以降低植酸与蛋白质的螯合作用,同时提高鱼体内源性蛋白酶的活性,使蛋白质的消化吸收率提高,从而降低氮的排泄率,从而降低了养殖水体氮磷污染的程度。鲁媛媛等[22]研究发现在植物性饲料中添加植酸酶的同时添加非淀粉多糖酶,可以促进植物性饲料中的果胶、非淀粉多糖和纤维素等聚合物分解为小分子物质,降低消化道的粘度,改善鱼类的消化机制,促进鱼类的生长速度,并进一步提高鱼体营养物质的消化吸收,从而降低氮的排泄率,但非淀粉多糖酶的添加对磷排泄率没有显著影响。
在饲料中添加无机盐类物质也可以对海洋生物的生长代谢产生一定影响。如张璐等[23]研究发现在鱼饲料中添加无机盐混合物可以提高鲈鱼对磷的利用率;杨金芳等[24]发现在低盐度水体中添加K+可以提高对虾的渗透压调节能力,显著影响凡纳滨对虾的生长,当水体中K+添加量为10 mg/L 且饲料中K+添加量为0.6%时,凡纳滨对虾的生长和生存状态较好,水体K+添加量为20 mg/L 且饲料K+添加量为0.3%时,凡纳滨对虾耗氧率和排氨率最低,有利于提高摄食能量的利用效率。
3 结 语
随着海水养殖业的不断发展,其带来的一系列氮、磷污染问题逐渐引起人们的重视。而海洋动物的氮、磷排泄对于浮游植物生产力、海洋生态系统营养循环、沉积物-水界面生源要素迁移均产生重要的影响。对影响鱼、虾、贝类等海洋生物氮、磷代谢的主要因素进行分析,对于寻求既能促进生物生长,又可减少氮、磷排泄的养殖方式,保护海洋环境具有一定的科学意义。
[1] 宋金明,李学刚,邵君波,等.南黄海沉积物中氮、磷的生物地球化学行为[J].海洋与湖沼,2006,37(4):370-376.
[2] 侯晓飞.半现场条件下温度、投喂水平和短期盐度胁迫对许氏平鲉(Sebastes schlegeli)氨氮、磷排泄影响的研究[D].青岛:中国海洋大学,2008.
[3] 周毅,杨红生,张福绥.海水双壳贝类的N、P 排泄及其生态效应[J].中国水产科学,2003,10(2):165-168.
[4] 薛飞,周维仁,徐为民.鱼类氮、磷营养与水体环境[J].河北渔业,2004,3:1-3.
[5] Smaal A C,Vonck A P M A.Seasonal variation in C,N and P budgets and tissue composition of the mussel Mytilus edulis[J].Mar.Ecol.Prog.Ser.,1997,153:167-179.
[6] Atkinson M,Smith S V.C:N:P ratios of benthic marine plants[J].Limnol Oceanogr,1983,28:568-574.
[7] Cui Y,Wootton R.J.Bioenergetics of growth of a cyprinid,Phoxinus phoxinus:the effect of ration,temperature and body size on food consumption,faecal production and nitrogenous excretion[J].Fish Biol.,1988,33:431-443.
[8] Brett J R,Zala C A.Daily pattern of nitrogen excretion and oxygen consummationa of sockeye salmon(oncorhynchus nerka)[J].Fish.RES.Board Can.,1975,32:2479-2486.
[9] Kaushik S J,Dabrowski K,Luquet P.Experimental studies on some tropic relationships in juvenile pike[J].Fish Biol.,1985,26:171-180.
[10] 李松青,林小涛,李卓佳,等.摄食对凡纳滨对虾耗氧率和氮、磷排泄率的影响[J].热带海洋学报,2006,25(2):44-48.
[11] 林小涛,梁旭方,吴杰,等.温度对罗氏沼虾亲虾代谢的影响[J].生态学杂志,1999,18(3):15-18.
[12] 宋协法,刘鹏,葛长字.温度、盐度交互作用对凡纳滨对虾耗氧和氨氮、磷排泄的影响[J].渔业现代化,2009,36(2):1-6.
[13] Needham A E.Factors affecting Nitrogen excretion in Crustacea[J].Physiol.Comp.Oecol.,1957,4:209-239.
[14] Pressley T A,Graves J S,Krall A R.Amiloride-sensitive ammonium and Sodium ion transport in the blue crab [J].The American Journal of Physiology,1981,241(5):370-378.
[15] 孙成波,李婷,李义军,等.多因子对日本囊对虾Marsupenaeus japonicus 氮磷代谢的影响[J].中山大学学报(自然科学版).2012,51(2):91-96.
[16] Dall W.Oxygen consumption and ammonian-N excretion in fed and starved tiger prawns,Penaeus esculentus Haswell [J].Aquaculture,1986,55:23-33.
[17] 姜祖辉,王俊.菲律宾蛤仔氮、磷代谢的初步研究[J].青岛大学学报,1999,12(2):68-73.
[18] MeGraw,W.,Teiehert-Coddington,D.R,Rouse,D.B.,et al.Higher minimum dissolved oxygen concentrations increase penaeid shrimp yields in earthen ponds[J].Aquaculnire,2001,199(3-4):311-321.
[19] 崔莹,减维玲,马海娟.南美白对虾瞬时耗氧速率与溶氧水平和海水盐度的相关关系[J].上海水产大学学报,2003,12(l):76-79.
[20] 赖晓芳,沈善瑞,吕海波.氨态氮和pH 对脊尾白虾氮磷代谢的影响[J].安徽农业科学,2013,41(13):5745-5746.
[21] Lin,Y.C.,Chen,J.C.Acute toxieity of ammonia on LitoPenaeus vannamei Boone juveniles at different salinity levels[J].Journal of ExperimentalMarineBiologyandEcology,2001,259(l):109-119.
[22] 鲁媛媛.外源植酸酶和非淀粉多糖酶对大菱鲆幼鱼生长和代谢机能的影响[D].大连:大连海洋大学,2014.
[23] 张璐,艾庆辉,张春晓,等.不同无机盐预混料含量对鲈鱼生长和钙磷代谢的影响[J].水产科学,2008,27(1):1-4.
[24] 杨金芳.添加钾离子对低盐度水体养殖凡纳滨对虾(Litopenaeus vannamei)的生长与生理特性的影响[D].湛江:广东海洋大学,2011.
