叉角厉蝽对黄野螟幼虫的捕食功能反应
2015-12-09李文华贾彩娟陈惠平陈科伟
李文华,贾彩娟,陈惠平,付 浪,温 健,陈科伟*
(1.深圳市梧桐山风景区管理处,深圳 518004;2.华南农业大学农学院,广州 501640)
黄野螟Heortia vitessoides (Moore)属鳞翅目Lepidoptera,螟蛾科 Pyralidae,齿螟亚Odontiinae (Munroe,1977),主要取食沉香属Aquilaria 的少数植物,在我国仅发现为害土沉香Aquilaria sinensis (又名白木香) (苏跃平,1994)。土沉香是我重要的香料与药用资源植物,曾遭到大肆损毁和采伐,被列为国家2 级珍稀濒危保护植物。近年来,随着土沉香人工种植面积的日益扩大,黄野螟种群数量急剧增加,在局部地区暴发成灾。如2008年以来,在广东中山市及化州市均有黄野螟大规模暴发为害(陈志云等,2011;乔海莉等,2013),2012-2013年期间,在深圳梧桐山土沉香种植区发现该虫的发生为害。黄野螟大发生时,单株虫口数量可从几百到上千头,数天内便将整株叶片取食殆尽,造成光秃无叶,严重削弱植株长势,影响其经济价值。有关黄野螟的防治也逐渐成为土沉香种植区亟待解决的问题。
目前,有关黄野螟的研究资料较少,仅有的几篇文献报道,主要侧重于其发生规律以及生物学、生态学特征方面(苏跃平,1994;陈志云等,2011;乔海莉等,2013),甚少涉及其防治方法(陈志云等,2012)。由于除人工种植基地外,大部分土沉香野生于杂树林之中,零星分散,且植株高大,给用药带来诸多不便。为寻找治理黄野螟的有效方法,笔者曾在室内发现叉角厉蝽Eocanthecona furcellate (Wolff)对黄野螟幼虫有较强的攻击能力,在林间释放一定数量的叉角厉蝽后,在野外也观察到其捕食黄野螟幼虫的现象,表现出较好的应用前景。为进一步定量评估叉角厉蝽对黄野螟的捕食效能,本文开展了叉角厉蝽对黄野螟的捕食功能反应研究,可为田间释放与应用提供相应的科学依据。
1 材料与方法
1.1 供试昆虫
叉角厉蝽E.furcellate (Wolff):采自华南农业大学附近菜场番薯地,室内以斜纹夜蛾Spodoptera litura 及黄粉虫Tenebrio molitor 幼虫饲养约20 代。饲养条件:利用人工气候箱设置温度26±1℃,相对湿度60%~70%,光照L∶D=14∶10。供试的叉角厉蝽各龄若虫及成虫均经饥饿处理24 h。
黄野螟H.vitessoides (Moore)各龄幼虫:试验所需的黄野螟各龄幼虫直接从深圳梧桐山风景区土沉香植株上采集,在室内用新鲜土沉香叶片饲养。饲养条件同上。
1.2 试验方法
1.2.1 叉角厉蝽成虫对黄野螟幼虫的捕食行为观察
选取黄野螟3 龄幼虫1 头,放置于洁净的培养皿(Φ=15 cm)中,同时提供2-3 片新鲜的土沉香叶片供其取食,然后移入1 头经饥饿24 h 的叉角厉蝽成虫,与黄野螟幼虫相距约10 cm。观察叉角厉蝽对黄野螟幼虫的搜索与捕食行为,每次持续观察2 h,重复10 次。当将取食对象更换为黄野螟5 龄幼虫时,重复上述步骤。
1.2.2 叉角厉蝽对黄野螟幼虫的捕食功能反应
本试验分别研究叉角厉蝽3 龄、5 龄若虫以及成虫这3种虫态,对黄野螟3 龄与5 龄幼虫的捕食功能反应。以透明塑料盒(15 cm×10 cm×5 cm)作为测试容器,盒盖上挖一5 cm×5 cm 开口,并粘上金属纱网,用于通风透气。猎物密度设置为3、6、9、12 及15 头/盒5 个梯度水平。将相应虫态及数量的黄野螟幼虫连同少量土沉香叶片放入测试容器,然后移入经饥饿24 h 处理的相应虫态的叉角厉蝽1 头,24 h 后统计其捕食量。各密度处理重复5 次。
1.2.3 捕食者自身密度对功能反应的影响
分别以1、2、4、6、8、10 头/盒密度的叉角厉蝽5 龄若虫与50 头黄野螟5 龄幼虫组合,统计24 h 后的捕食量,设5 次重复。
1.2.4 空间异质性对捕食效应的影响
试验设置养虫盒(15 cm×10 cm×5 cm)、笼罩植株(带虫枝条)和野外这3 个不同的生态环境空间,其中笼罩的空间大小为30 cm×20 cm×10 cm。野外则选择距地面高度约2 m,长约2.5 m的土沉香枝条,在枝条中间位点放置相应数量的黄野螟3 龄幼虫(野外观察表明,黄野螟3 龄幼虫仍有较强的聚集取食习性,4-5 龄幼虫才开始逐渐扩散为害)。在这3 个不同空间分别放入5、10、15 头黄野螟3 龄幼虫,每个试验空间放入叉角厉蝽5 龄若虫1 头,观察24 h 的捕食结果。试验设5 个重复。
1.3 数据处理
(1)不同虫态叉角厉蝽对黄野螟捕食的功能反应采用Holling-Ⅱ圆盘方程进行拟合,其方程为:。其中Na 为被捕的猎物数(头),N 为初始猎物数量(头),a'为叉角厉蝽的瞬时攻击率,T 是试验处理时间,Th为叉角厉蝽处理猎物的时间(d)(徐汝梅,1987)。
(2)叉角厉蝽自身密度对其寻找效应的影响根据Hassell 和Rogers (1972)提出的模型进行拟合:E=Q×P-m。其中E 为寻找效应,Q 为寻找参数,m 为相互干扰参数,P 为叉角厉蝽密度。而E可由公式E=Na/(N×P)估算(丁岩钦,1980)。
本研究所有数据分析均用SAS 9.0 数据处理软件,涉及单因素方差分析,非线性回归,用Duncan 氏新复极差法进行多重比较。
2 结果与分析
2.1 叉角厉蝽对黄野螟幼虫捕食行为
将叉角厉蝽放入培养皿后,叉角厉蝽会先静止休息一下(或直接爬行,再休息),然后在培养皿内爬行,期间触角会以一定频率地上下左右摆动。当接近黄野螟幼虫时,触角停止摆动,口器前伸并试探性接触虫体。对于黄野螟低龄(3 龄)幼虫,叉角厉蝽通常以口针直接刺入虫体。对于高龄(5 龄)幼虫,当叉角厉蝽口器接近虫体时,黄野螟幼虫会用头部抵抗叉角厉蝽的进攻,但叉角厉蝽保持口器前伸状态(或不完全收回),停顿在虫体附近,等黄野螟停止反抗后,继续进攻,直到刺入虫体为止;一旦叉角厉蝽将口针刺入黄野螟幼虫体内,虽然黄野螟幼虫会继续抵抗,但叉角厉蝽通常不会拨出口针,20-30 s 后,黄野螟幼虫逐渐处于静止,2-3 mins 后叉角厉蝽开始进行取食。叉角厉蝽的口针通常从黄野螟低龄幼虫体侧或腹部插入(偶尔从背部刺入),而捕食黄野螟高龄幼虫时,经常从其腹部末端插入。
2.2 叉角厉蝽对黄野螟幼虫捕食作用的功能反应
在试验设置的猎物密度梯度范围内,3种虫态的叉角厉蝽对黄野螟幼虫的捕食量均随猎物密度的增加而增大,但随着猎物密度的不断增加,叉角厉蝽捕食量增幅逐渐减缓(表1,表2)。另外,由于猎物个体大小的差异,同一虫态的叉角厉蝽对黄野螟3 龄幼虫的捕食数量高于5 龄幼虫(表1,表2)。
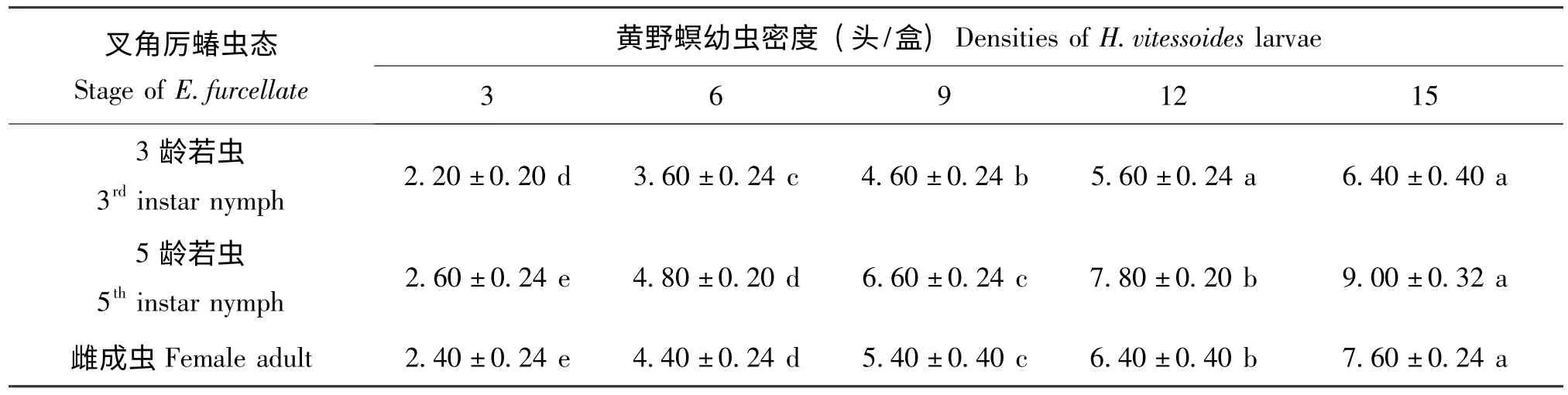
表1 叉角厉蝽对黄野螟3 龄幼虫的捕食量(头/d)Table 1 Predatory amount of Eocanthecona furcellate on the 3rd instar larvae of Heortia vitessoides
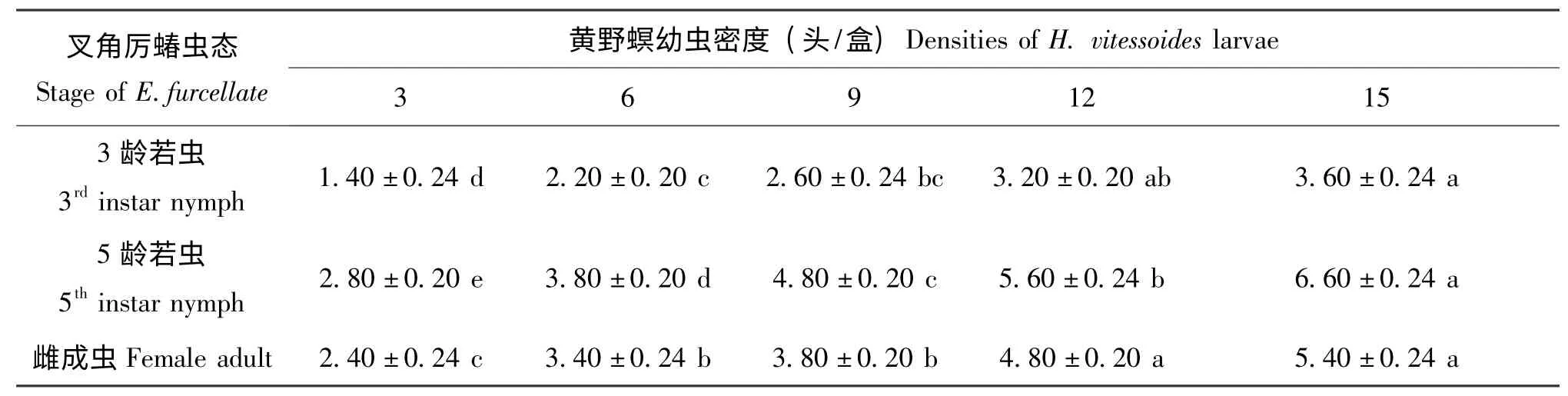
表2 叉角厉蝽对黄野螟5 龄幼虫的捕食量(头/d)Table 2 Predatory amount of Eocanthecona furcellate on the 5th instar larvae of Heortia vitessoides
不同虫态的叉角厉蝽对黄野螟幼虫的捕食功能反应均符合Holling Ⅱ模型(表3)。从不同虫态叉角厉蝽对黄野螟幼虫的攻击率来看,其捕食能力的大小顺序为:高龄若虫(5 龄) >雌成虫>低龄若虫(3 龄),这与不同虫态叉角厉蝽的实际捕食量及根据理论模型所计算出的日最大捕食量的结果相一致(表1,表2,表3)。
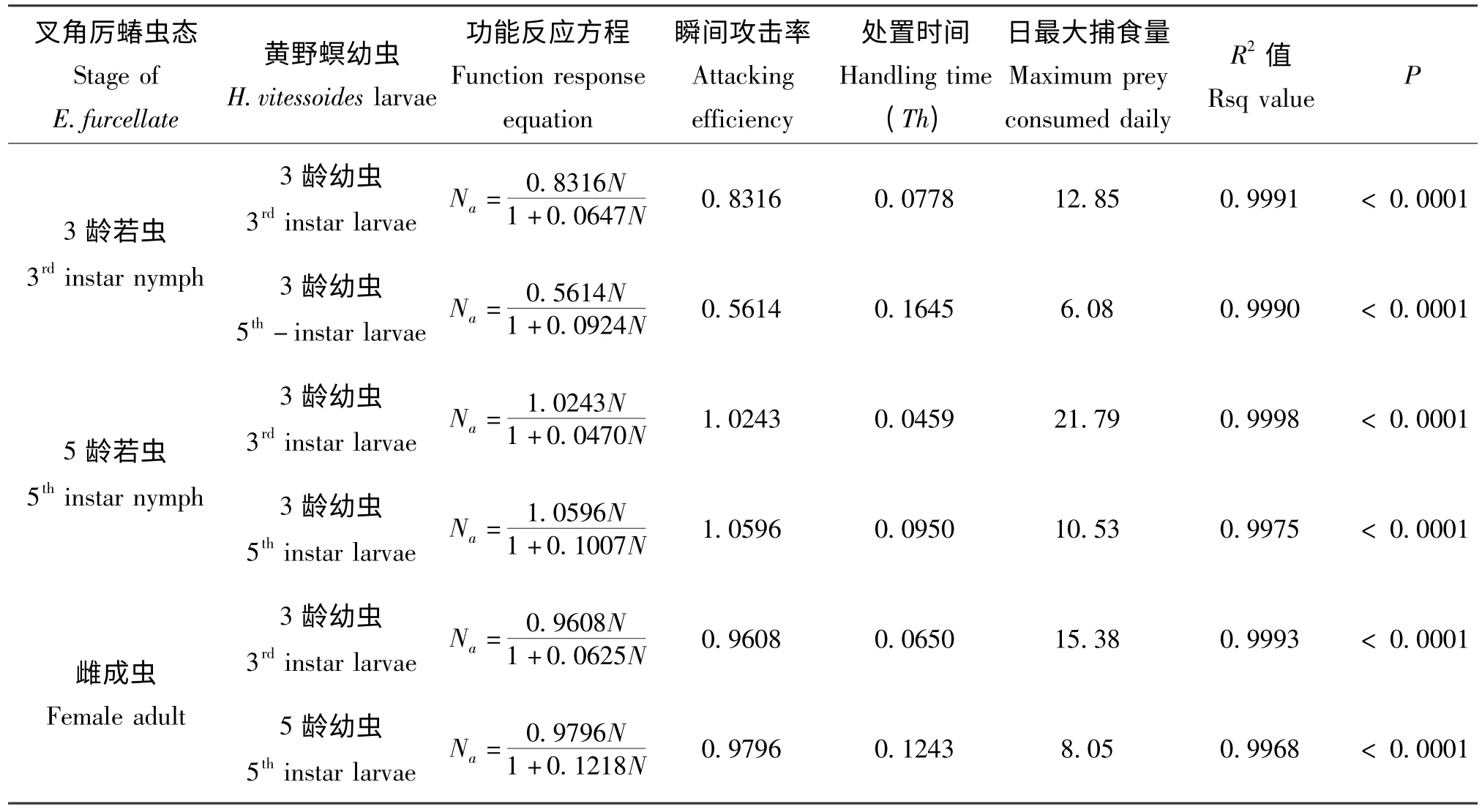
表3 叉角厉蝽对黄野螟幼虫的捕食功能反应参数Table 3 Functional response parameters of Eocanthecona furcellate on the larvae of Heortia vitessoides
2.3 叉角厉蝽自身密度对猎物寻找效应的影响
当猎物密度为50 头黄野螟5 龄幼虫时,随着叉角厉蝽自身密度的增加,其捕食量也显著上升。但从叉角厉蝽对猎物的寻找效应来看,随着自身密度的增加,其对猎物的寻找效应也逐步下降,其寻找效应方程为:E=0.1452×P-0.3923(R2=0.99,P<0.0001),说明在有限的空间范围内,叉角厉蝽种群内不同个体之间存在相互干扰作用。

表4 不同密度的叉角厉蝽对黄野螟幼虫的捕食量Table 4 Predatory amount of Eocanthecona furcellate at different densities on Heortia vitessoides larvae
2.4 空间异质性对叉角厉蝽捕食效应的影响
叉角厉蝽对黄野螟幼虫的捕食效能除受到猎物及自身密度影响外,其在不同空间的捕食量也存在一定的差异(表5)。在养虫盒中,空间狭小且环境条件稳定,叉角厉蝽的捕食率最高。笼罩的空间比养虫盒,大大增加了叉角厉蝽搜寻猎物的范围,其捕食率开始下降。而在空旷的大田间环境时,由于受不稳定的气候因子及复杂的生物因子的影响,叉角厉蝽的捕食效能急剧下降,其理论日最大捕食量为10.91 头黄野螟3 龄幼虫,不到养虫盒环境条件下捕食量的一半(见表5)。

表5 叉角厉蝽在不同空间对黄野螟幼虫的捕食效应Table 5 Functional response of Eocanthecona furcellate to Heortia vitessoides larvae at different space
3 结论与讨论
叉角厉蝽是农林作物上一种重要的捕食性天敌,在我国华南地区广泛分布,其捕食范围涉及鳞翅目、膜翅目、鞘翅目、半翅目等40种以上的幼虫,其中尤喜鳞翅目幼虫(朱涤芳,1990),如对斜纹夜蛾(Rani et al.,1993;蒋杰贤等,2001)、甜菜夜蛾(何旭诺等,2013)、菜青虫(张敏玲等,1996)、尺蠖(谢钦铭等,2001)等均有很强的捕食作用。但目前尚未见任何有关叉角厉蝽取食黄野螟幼虫的研究报道。通过对叉角厉蝽的捕食行为观察表明,叉角厉蝽对黄野螟低龄(3 龄)与高龄(5 龄)幼虫均表现出良好的搜索与捕食能力,说明黄野螟幼虫是其适宜的捕食对象。
功能反应是评价捕食天敌对害虫控制作用的重要方法。本研究结果表明,不同虫态的叉角厉蝽对黄野螟幼虫的捕食功能反应均符合Holling Ⅱ模型。不同虫态叉角厉蝽对黄野螟3 龄幼虫的捕食量均高于5 龄幼虫,这可能有两方面的原因。一是猎物个体大小的差异,黄野螟3 龄幼虫的个体明显小于5 龄幼虫,显然叉角厉蝽需要捕食较多的3 龄幼虫才能满足其营养需求;二是叉角厉蝽对黄野螟低龄幼虫有较强的攻击力,而黄野螟高龄幼虫受到攻击时会有一定的抵抗行为,这在一定程度上增加了叉角厉蝽处置猎物的时间,从而降低其捕食率。从不同虫态叉角厉蝽的捕食量来看,高龄若虫的捕食能力最强,其次为雌成虫,低龄若虫的捕食能力最弱。低龄若虫的捕食量较低主要受其自身捕食能力的制约,而高龄若虫的捕食量超过雌成虫,这可能是高龄若虫需要获取较多的能量用于进一步的生长发育,为进入成虫阶段贮备较多的物质基础。
在一定的空间范围内,当猎物数量固定时(但仍超过捕食者的最大捕食量),天敌对猎物的功能反应常常表现出自身密度干扰效应(林克剑等,2006;张彦龙等,2012)。从本实验的结果来看,叉角厉蝽的捕食效能也受到其自身密度的影响。虽然叉角厉蝽捕食猎物的总体数量会随同伴数的增加而逐渐增加,但其捕食量的增加与自身密度的增大不是线性关系,而是呈现一种减速增长,其对猎物的寻找效应会因自身密度的增加而逐步下降,表明个体之间可能因觅食竞争而产生一定的干扰作用,导致其整体捕食效能的下降。
另外,叉角厉蝽的捕食效率会因猎物所处环境空间的扩大而急剧下降,尤其是在野外条件下,会受不稳定的气候因子(如光照、温湿度、雨水、风等)及复杂的生物因子(如其他捕食性天敌的竞争或其他一些猎物的存在而转移对目标害虫的攻击)的影响,从而大大影响其对猎物的搜寻效率(秦玉洁等,2002)。因而,在田间进行天敌释放时,需综合考虑天敌自身的捕食能力、猎物的种群数量、外部的环境条件等因素,以确定适宜的释放方式与释放量。
总得来说,叉角厉蝽对黄野螟幼虫表现出较强的捕食能力。虽然叉角厉蝽的食谱范围非常广泛,在一定程度上会削弱其对目标害虫的控制作用,但相比一些寄主专化性强的天敌,其野外定殖及生存的能力较强,在某一种猎物缺乏时可以通过捕食其他猎物而增加存活的机率,这对于害虫种群数量的持续控制是有利的一面。如果在黄野螟的发生期,通过在土沉香植株上定点、定时补充释放一定数量的叉角厉蝽,可望获得较为理想的防治效果。
References)
Chen ZY,Li DW,Wang L,et al.Studies on biological characteristics of Heortia vitessoides Moore on Aquilaris sinensis [J].China Plant Protection,2011,31 (11):10-14.[陈志云,李东文,王玲,等.土沉香黄野螟生物学特性研究[J].中国植保导刊,2011,31 (11):10-14]
Chen ZY,Wang L,Li DW,et al.A screening experiment on insecticides against Heortia vitessoides Moore [J].China Forest Science and Technology,2012,26 (1):117-119.[陈志云,王玲,李东文,等.黄野螟防治药剂筛选试验[J].林业科技开发,2012,26 (1):117-119]
Ding YQ.Principles and Application of Insect Population Mathematical Ecology[M].Beijing:Science Press,1980,197-213.[丁岩钦.昆虫种群数学生态学原理与应用[M].北京:科学出版社,1980,197-213]
Hassell MP,Rogers DJ.Insect parasite responses in the development of population models [J].Journal of Animal Ecology,1972,41 (3):661-667.
He XN,Xian JD,Chen R,et al.Effects of four insect feed on development and reproduction of Cantheconidea furcellata(Hemiptera:Asopinae) [J].Journal of Environmental Entomology,2013,35 (6):799-803.[何旭诺,冼继东,陈然,等.4种昆虫饲料对叉角厉蝽生长发育和繁殖的影响[J].环境昆虫学报,2013,35 (6):799-803]
Jiang JX,Liang GW.The selective predation of Cantheconidea furcellate Wolff on the different instar larvae of Spodoptera litura in coexistence of three age type[J].Acta Ecologica Sinica,2001,21 (4):684-687.[蒋杰贤,梁广文.叉角厉蝽对斜纹夜蛾不同龄幼虫的选择捕食作用[J].生态学报,2001,21 (4):684-687]
Lin KJ,Wu KM,Liu SB,et al.Functional responses of Chrysopa sinica,Propylaea japonica and Leis axyridis to Bemisia tabaci [J].Chinese Bulletin of Entomology,2006,43 (3):339-343.[林克剑,吴孔明,刘山蓓,等.中华草蛉、龟纹瓢虫和异色瓢虫对B 型烟粉虱的捕食功能反应[J].昆虫知识,2006,43 (3):339-343]
Munroe E.Synopsis of Heortza lederer,with descriptions of a new species and a new subspecies (Lepidoptera:Pyralidae:Odontiinae)[J].The Canadian Entomologist,1977,109 (3):429-441.
Qiao HL,Lu PF,Chen J,et al.Biological characteristics and occurrence patterns of Heortia vitessoides [J].Chinese Journal of Applied Entomology,2013,50 (5):1244-1252.[乔海莉,陆鹏飞,陈君,等.黄野螟生物学特性及发生规律研究[J].应用昆虫学报,2013,50 (5):1244-1255]
Qin YJ,Wu WJ,Liang GW,et al.The functional response of Campylomma chinesis to Thrips palmi [J].Journal of South China Agricultural University,2002,23 (2):27-29.[秦玉洁,吴伟坚,梁广文,等.中华微刺盲蝽对节瓜蓟马的捕食效应[J].华南农业大学学报(自然科学版),2002,23 (2):27-29]
Rani PU,Wakamura S.Host acceptance behaviour of predatory Pentatomid,Eocanthecona furcellata (Wolff) (Heteroptera:Pentatomidae)towards larvae of Spodoptera litura (Lepidoptera:Noctuidae)[J].Insect Science and its Application,1993,14 (2):141-147.
Su YP.The biology of Heortia vitessoides [J].Journal of Chinese Medicinal Materials,1994,17 (12):7-9.[苏跃平.白木香黄野螟生物学特性[J].中药材,1994,17 (12):7-9]
Xie QM,Liang GW,Luo S,et al.Observation of the predacious function of a bug Cantheconidae furcellata on litchi looper Thalassodes proquadria [J].Jiangxi Science,2001,19 (1):21-23.[谢钦铭,梁广文,罗诗,等.叉角厉蝽对绿额翠尺蛾幼虫的捕食作用的初步研究[J].江西科学,2001,19 (1):21-23]
Xu RM.Insect Population Ecology [M].Beijing:Beijing Normal Universtity Press,1987,331-338.[徐汝梅.昆虫种群生态学[M].北京:北京师范大学出版社,1987,331-338]
Zhang ML,Lu CC.Rearing of Eocanthecona furellata [J].Natural Enemies,1996,18 (1):74-77.[张敏玲,卢传权.叉角厉蝽的饲养[J].昆虫天敌,1996,18 (1):74-77]
Zhang YL,Yang ZQ,Wang XY,et al.Functional response of the parasitoid Sclerodermus sp.(Hymenoptera:Bethylidae)to the third instar larvae of host Monochamus alternatus (Coleoptera:Cerambycidae)[J].Acta Entomologica Sinica,2012,55 (4):426-434.[张彦龙,杨忠歧,王小艺,等.松褐天牛肿腿蜂对寄主松褐天牛三龄幼虫的功能反应[J].昆虫学报,2012,55 (4):426-434]
Zhu DF.Studies on the characteristics of Cantheconidea furcellata(Hemiptera:Pentatomidae,Asopinae)[J].Natural Enemies,1990,12 (2):71-74.[朱涤芳.叉角厉蝽生物学特性研究[J].昆虫天敌,1990,12 (2):71-74]
