广州市典型森林植被的结构及多样性研究
2015-12-08翟石磊陈步峰林娜史欣潘永军邹志谨
翟石磊,陈步峰,林娜,史欣,潘永军,邹志谨
中国林业科学研究院热带林业研究所,广东 广州 510520
广州市典型森林植被的结构及多样性研究
翟石磊,陈步峰*,林娜,史欣,潘永军,邹志谨
中国林业科学研究院热带林业研究所,广东 广州 510520
分别遴选广州城市尺度上帽峰山、白云山、天河公园样点的常绿阔叶林各0.48、0.48、0.24 hm2,采用群落生态学的方法对比研究了3种植被类型的群落结构、物种多样性和主要种群间的关联性。结果表明3种植被类型的物种数、林分密度、郁闭度均表现为常绿阔叶天然次生林(EBSF)>常绿阔叶人工-天然混交林(EBANF)>常绿阔叶人工林(EBP);乔木层、灌木层及所有木本植物的Shannon-Wiener指数、Gleason丰富度指数也表现出相同的趋势,且EBSF表现为:灌木层>乔木层,EBANF和EBP表现为:乔木层>灌木层,而Pielou均匀度指数则表现出多变的特征;EBSF、EBANF、EBP的径级结构分别表现为标准、微双峰、多峰的倒“J”型;EBSF与EBANF的20个优势木本植物间整体呈现不显著的正关联,20个优势种构成的190个种对的关联性在χ2检验、PC值及AC值上均表现出与总体一致的不显著正关联,且EBSF物种间的正关联性更强。结果反映了广州市局部地区常绿阔叶天然次生林、人工-天然混交林、人工林在群落结构、物种多样性以及主要种群间的关联性上的差异,以典型代表整体,旨在为广州近自然林及美丽城镇森林景观的建设及评价提供理论依据。
广州;城市森林;群落结构;物种多样性;种间联结
城市化快速发展的过程带来了自然环境的破坏,给城市生态平衡带来了严峻的考验。城市森林作为城市生态系统的重要组成成分,在美化环境(陈勇等,2013;张永利等,2010)、净化空气(邵海荣等,2005;Alvey,2006)、调节小气候(廖建华等,2013)、减弱热岛效应(Alberti,2005;Fisher et al.,2006;Takebayashi et al.,2007)、固定碳素(Nowak et al.,2002;Myeong et al.,2006)等方面具有独特的生态功能;而且,在人们逐渐由物质享受转向精神享受的今天,城市森林更是人们休闲游憩、强身健体的重要场所。城市森林群落的物种组成、结构和物种多样性是认识、管理森林进而保证城市生态健康的基础。种群联结是指不同物种在空间分布上的相互关联性,包括空间分布关系与功能依赖关系(陈玉凯等,2011),能有效的反映群落发育过程中物种与物种、物种与环境之间的相互关系,对认识群落的结构、功能、演替发展具有重要意义(奇凯等,2010)。物种多样性是物种丰富度与均匀度的综合反映,它不仅可以反映群落与生态系统的特征,也能间接的反映群落的组织水平、稳定程度、演替阶段及生境差异(郭连金等,2007),较高的物种多样性有助于维持群落稳定,促进可持续发展(Ares et al.,2010)。
广州市现有的森林多为人工林,保存比较完整的天然次生林较少。为了更好的发挥森林在改善广州生态环境中的作用,有必要对广州地区有代表性的天然次生林、人工林、人工-天然混交林的群落特征进行对比研究,找出差异,以期为改进或建立优质林分提供建议。近些年来,国内学者(苏志尧等,2001;彭少麟等,1996;林伟强等,2006;张璐等,2003)对白云山和帽峰山的天然次生林和人工-天然混交林的群落结构、演替动态及种间关系进行一定的研究,但对不同植被类型的种群、群落特征的对比研究鲜有报道。本研究以帽峰山常绿阔叶次生林、白云山常绿阔叶人工-天然混交林、天河公园和白云山人工林为研究对象,对其物种组成、径级结
构、物种多样性、主要木本植物的种间联结性进行对比研究,旨在找出人工群落或人工促进演替的半天然群落同天然次生群落的差距,为广州美丽城镇森林的建设与管理提供理论依据。
1 研究区及样地概况
广州位于广东省东南部、珠三角洲北缘,地理位置界于东经112°57′~114°03′,北纬22°26′~23°56′。属南亚热带季风气候,气候温和,雨量充沛,年均温度22.0 ℃,年均降雨量1800.0~2200.0 mm,多集中在4─9月份,占全年降雨的82.1%,年平均相对湿度79.0%,土壤类型主要有赤红壤、山地草甸土、山地黄壤、山地红壤,地带性土壤为赤红壤;样地分别布设在天河公园、白云山、帽峰山,其自然条件等基本情况见表1。

表1 森林监测区的基本情况Table 1 Basic characteristics of forest monitoring area
2 研究方法
2.1 样地调查方法
采用典型抽样的方法,在全面踏查的基础上,于帽峰山布设常绿阔叶次生林20 m×20 m的样方12个、白云山布设常绿阔叶人工-天然混交林 20 m×20 m的样方9个以及人工林20 m×20 m的样方3个、天河公园布设常绿阔叶人工林20 m×20 m的样方6个,总计1.2 hm2。将每个样方视为资源序列中的一个N维资源状态的综合,在每个样方内进行每木检尺,测量记录DBH(胸径)≥1.0 cm的各木本植株的种名、坐标、胸径、树高、冠幅、活枝下高、郁闭度等指标,同时记录样地的经纬度、海拔、坡度、坡向、土壤状况等环境因子。
2.2 数据分析
2.2.1 重要值的计算
重要值(IV)=(相对密度+相对优势度+相对频度)/3,范围为0%~100%。
式中,植株的相对优势度用林木胸高断面积来表示,相对密度、相对优势度、相对频度的计算公式如下:相对密度=某个种的个体数/全部种的个体总数×100%;相对优势度=某个种的胸高断面积/全部种的总胸高断面积×100%;相对频度=某个种的频度/全部种的总频度×100%。
2.2.2 物种多样性的计算
各类型森林不同层的物种多样性指数值取同一层不同群落算术平均值(李军玲等,2006;任国学等,2011)。目前,能较好衡量南亚热带森林群落物种多样性的指标是Shannon-Wiener指数(汪殿蓓等,2003),因此本文以Shannon-Wiener指数为主要衡量指标,Pielou均匀度指数与Gleason丰富度指数作为补充。
物种多样性的3个指标的公式(马克平,1994;马克平等,1994)如下:
Gleason丰富度指数:

Shannon-Wiener指数:

Pielou均匀度指数:

式中,S为物种总数;Pi=Ni/N,为第i个种的相对多度;N为所有物种个体数之和,Ni为第i个种的个体数。
2.2.3 总体关联性测度
种间联结性测度的最适取样数目为30~50个,样方面积为50~100 m2(王伯荪等,1985),样方数越多,种间联结性的计算结果越精确(王国梁等,2002)。本研究把样方分为10 m×10 m的小样方,其中,帽峰山常绿阔叶次生林 46个,白云山常绿阔叶-人工天然混交林36个。
基于出现-不出现数据的方差比率测定多物种间的总体联结性。计算公式(Schluter,1984)为:

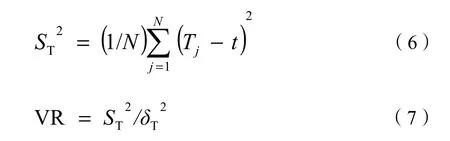
式中:VR为物种总体联结性指数;S为总物种数;ni为物种 i出现的样方数;N为总样方数;Tj为样方j内出现的研究物种的总数;t为样方中物种的平均数,t=(T1+T2+…+TN)/N。
独立性零假设条件下,VR=1;当VR>1时,物种间呈正关联;当VR<1时,物种间呈负关联。物种间的正负关联可以相互抵消。故采用统计量W来检验VR值偏离1的程度(显著性),W=N·VR,若W落在χ2分布界限(χ20.95N 2.2.4 种间联结性测度 χ2检验能够相对客观、准确的判断种对联结的显著性,但其在一定程度上模糊了相关性判定。AC和PC值可以反映出χ2检验证明不显著的联结性。以χ2检验为基础,AC、PC值为辅助参数共同分析南亚热带森林的种间联结性效果会更好(王伯荪等,1985;张倩媚等,2006;李军玲等,2010)。 2.2.4.1 种间联结性χ2检验 种间联结性的 χ2检验一般用于确定实测值与机率基础上的预期值之间偏差的显著程度。建立2×2列联表,采用连续性校正公式(Yates et al.,1972)进行计算: 式中,a为两物种同时出现的样方数;b和 c分别为仅有1个物种出现的样方数;d为两物种均未出现的样方数;χ2没有负值,种间联结性的正负以ad与bc值来判断,ad>bc时,为正联结,ad 2.2.4.2 种间联结系数AC的测度 用AC值进一步检验χ2统计量检测的结果,并且进一步说明种间的联结程度。对AC出现+1、-1的情形,把a、b、c、d为0的都加权为1,这样测度的结果便于在两个林型间进行比较,更能体现种间关系的动态变化和延续性(张倩媚等,2006)。 2.2.4.3 共同出现百分率PC的测度 共同出现百分率PC,即Jaccard指数,用来表示种对相伴出现的概率和联结程度。其可在一定程度上克服AC值受到d影响大而造成的偏差。其公式(史作民等,2001)如下: PC值域为[0, 1],越趋近1,表示种对的正联结越紧密。 表2 群落的基本特征Table 2 Basic characteristics of communities 3.1 群落类型的划分 采用组间平均联结法对 30个样地基础数据进行聚类分析,将3个区的调查样地划分为6个不同的群落类型,以乔、灌木优势种命名,分别为:短序润楠(Machilus breviflora)+楠木(Phoebe zhennan)-九节(Psychotria rubra)、润楠(Machilus pingii)+藜蒴(Castanopsis fissa)-银柴(Aporosa dioica)、朴树(Celtis sinensis)+润楠-黄牛木(Cratoxylum cochinchinense)、荷木(Schima superba)+楠木-降真香(Dalbergia benthamii)、蒲 桃(Syzygium jambos)+尾叶桉(Eucalyptus urophylla)-鱼尾葵(Caryota ochlandra)、阴香(Cinnamomum burmanni)+白兰花(Michelia alba)-九节。以《中国植被》的分类原则与系统为纲,参照《广东森林》的植被分类原则,将以上群落归为常绿阔叶人工林(EBP)、常绿阔叶人工-天然混交林(EBANF)、常绿阔叶次生林(EBSF)3种植被类型(表2)。 3.2 群落结构特征 3.2.1 群落的物种组成 调查资料的统计可知,C01的木本植物由 47个物种组成,隶属于26科35属,IV>2.0%的优势乔木有短序润楠、楠木、黄樟(Cinnamomum porrectum)等 10个种,重要值的和占所有种的56.0%;C02的木本植物由40个物种组成,隶属于21科 32属,IV>2.0%的优势乔木有藜蒴、润楠、中华锥(Castanopsis chinensis)等11个种,重要值的和占64.9%;C03的木本植物由20个物种组成,隶属于12科17属,IV>2.0%的优势乔木有朴树、润楠、楠木等15个种,重要值的和占89.6%;C04的木本植物由24个物种组成,隶属于17科23属,IV>2.0%的优势乔木有荷木、楠木、短序润楠等 7个种,重要值的和占74.8%;C05的木本植物由12个物种组成,隶属于7科11属,IV>2.0%的优势乔木有蒲桃、尾叶桉、落羽杉(Taxodium distichum)等10个种,重要值的和占91%;C06群落的木本植物由20个种组成,隶属于15科17属,IV>2.0%的优势乔木有阴香、白兰花、大叶紫薇(Lagetstroemia speciosa)等10个种,重要值的和占65.2%。 群组水平上,物种数及其科属数表现为:EBSF>EBANF>EBP,林分密度与郁闭度也表现出相同的趋势(表2),表征出人工林向天然林的正向演替过程中物种数、林分密度、郁闭度逐渐增大,而平均胸径、树高却减小。 3.2.2 群落的径级结构 3种植被类型EBSF、EBANF、EBP的木本植物的平均胸径分别为 8.9、17.6、19.3 cm,呈递增的趋势。EBSF、EBANF的径级分布呈倒“J”型(图1),EBSF表现为标准的单峰趋势,个体数随径级的增加平稳的连续下降,而EBANF的“J”型表现为微双峰趋势,在9~13 cm范围内个体数相对较少,体现出人工-天然混交林的特点,同时也是人为干扰的结果。EBP的径级分布呈不规则的多峰倒“J”型,在15~23、31~37 cm、>45 cm中大径级范围内的个体数相对较多,反映出人工种植的特点。 图1 3种植被类型木本植物的径级分布Fig. 1 Tree size distribution of woody plants in three vegetation types 图2 3种植被类型所有木本植物同径级个体群重要值随径级结构的分布Fig. 2 Importance value of woody individual groups in the same tree size of three vegetation types 以5 cm为1径级,将个体分为9个组。把同一径级所有个体看作同一物种,借鉴重要值的计算方法得出3种植被类型木本植物同径级个体群重要值随径级结构的变化曲线(图2)。比较发现,EBSF与EBANF的同径级个体群的重要值随径级的增加呈下降的趋势,但中小径级个体(1.0 cm≤DBH﹤20 cm)在EBSF中的优势程度相对较高,而中大径级个体(DBH≥20 cm)在EBANF中的优势程度较高。EBP的同径级个体群的重要值随着径级的增加呈三峰分布,中径级(18 cm≤DBH≤24 cm)及大径级(DBH≥45 cm)个体的优势程度明显高于其他两个林型,而小径级(1.0 cm≤DBH≤15 cm)和中径级(25 cm≤DBH≤32 cm)的个体优势程度较小。 3.3 群落的物种多样性 3种植被类型的乔、灌层Shannon-Wiener指数、Gleason丰富度指数表现出相同的趋势:EBSF>EBANF>EBP,而Pielou均匀度指数却表现出多变的特征。EBSF的多样性指数、丰富度指数均表现为:灌木层>乔木层,而均匀度指数则相反,说明天然次生林林下植被物种数多,但分布不均匀,优势种与伴有种个体数相差较大。EBANF与EBP的多样性与丰富度指数表现为:乔木层>灌木层,均度指数表现为:EBANF:灌木层>乔木层,EBP:乔木层>灌木层(表 3)。这反映出广州常绿阔叶林中人工林与人工-天然混交林灌层的物种多样性及丰富度较差的特点,尤其是人工林中的灌木层。 表3 3种植被类型的乔灌层物种多样性指数Table 3 Species diversity indices of tree layer and shrub layer in three vegetation types CO4之外的 5种森林群落乔、灌木层的Shannon-Wiener指数与Gleason丰富度指数表现出基本一致的趋势(图3a、图3b):CO1和CO2:灌木层>乔木层;其他3个群落型:乔木层>灌木层,Pielou均匀度指数均表现为:乔木层>灌木层(图3c)。体现出群落物种多样性由丰富度与均匀度共同作用的特点。群落4的Shannon-Wiener指数乔木层明显小于灌木层(H′=3.659>3.069),这主要是因为其物种均匀度偏小的缘故(图3c)。该群落乔木层优势种比较突出,荷木、楠木、短序润楠3个优势种的重要值之和占乔木层的60%以上,优势种与稀有种的物种数差异较大,使得其均匀度偏小。 图3 6个森林群落乔灌层的物种多样性Fig. 3 Species diversity of three and shrub layers in six forest communities 3.4 主要木本植物的空间关联性 3.4.1 主要木本植物间的总体关联性 人工林更新时间短,主要木本植物多为人为配置树种,研究其种间关联性的意义不大,故不进行关联性的比较。依据两种植被类型木本植物的重要值,最终确定参与常绿阔叶次生林和常绿阔叶人工-天然混交林的木本植物各 20种,其重要值均在1.0%以上(山矾Symplocos sumuntia为0.78),分别占所有木本植物重要值的86.3%、92.4%。EBSF、EBANF植被类型主要木本植物的 VR值分别为1.33、1.02(表4),均大于1,表明物种间总体为正关联。其对应的统计检验量W分别为60.98、36.76,均落在χ2临界值范围内,说明物种间总体的关联性不显著。两种植被类型的对比发现,EBSF的总体物种间正关联性更强。 3.4.2 主要木本植物的种对关联性 图4a可知,EBSF、EBANF主要木本植物间的关联性表现出明显的不显著性,比例分别为87.4%和90%,反映出物种相对独立的格局。其20个优势种构成的190个种对的正关联、负关联、无关联的种对数百分比分别为 52.6%和 35.8%、46.9%和62.1%、0.5%和2.1%。其中,显著和极显著正关联 有14对和8对,显著和极显著负关联有9对和7对。帽峰山EBSF植被类型的种对主要为正关联,但其显著性(7.4%)较低,这与物种间的总体关联性一致。白云山EBANF植被类型的种对主要为负联结,这与物种间的总体正关联相矛盾,可能是群落人为干扰严重致使物种分布不均(尤其是下层灌木)的缘故。两种植被类型在显著-极显著水平上,均是正关联种对略占优势,但EBSF更多,体现了常绿阔叶次生林物种间正联结性更强的特点。阔叶次生林比人工-天然混交林木本植物相伴出现率高,种对正联结性更强的特点。 图4 两种植被类型的种间关联性χ2检验(a)、种间共同出现百分率(b)及种间联结系数(c)的种对数百分比Fig. 4 Percentage of species-pairs of χ2correction test, PC value and AC value in two vegetation types 表4 两种植被类型主要木本植物的总体关联性Table 4 Overall association of dominate woody plants in two vegetation types 图 4c可知,两种植被类型的种对联结系数在[-0.2,0.20)范围占据绝对优势,种对数比例为44.2%和40.5%。说明种对间整体虽然存在一定联系,但仍呈现独立分布的格局。在|AC|≥0.5范围内,EBSF的种对数占优势,占总对数的 23.7%。在AC∈[-0.5,-0.2)和[0.2, 0.5)内,EBANF的种对数占优势,占总对数的37.9%。表征出常绿阔叶次生林种间联结性更强的特点。 两种植被类型的种对共同出现百分率PC<0.1、0.1≤PC<0.3、0.3≤PC<0.5、PC≥0.5的种对百分比分别为 25.8%和 62.6%、54.7%和 28.4%、12.1%和6.3%、7.4%和2.7%(图4b)。EBSF在PC≥0.1的比例显著大于EBANF,其中,0.1≤PC<0.3范围内的种对数更是占绝对优势(54.7%)。而 EBANF则在PC<0.1内占绝对优势(62.6%),其中,PC=0的种对数占总对数的44.2%。表征出广州地区常绿 植被分类一直是植物学领域中争论的热点,而亚热带森林植被的划分也没有统一的标准(汪殿蓓等,2002)。不同林分起源的森林在物种组成、群落结构及外貌上会有显著的差异(庄雪影等,2002),而且主要优势种的差异在一定程度上能反映群落的结构特征(汪殿蓓等,2003)。本研究基 于以上理论再结合乔木种的重要值,用SPSS软件的组间平均联接法进行聚类分析,将样地划分为 6个群落和3个植被类型。其结果与深圳市主要群落类型(陈勇等,2013)的划分有一定的相似性,但把人工-天然混交林单独作为一个整体进行研究。 径级结构是判断植被群落稳定性和生长发育状况的重要指标。EBSF木本植物的径级结构表现为标准的倒“J”型,优势种的个体集中分布于中小径阶范围,林分结构合理,更新能力强,体现出群落稳定与正常的生长状态。而EBANF与EBP木本植物的径级结构分别呈微双峰、多峰的倒“J”型,中小径级个体分布相对较少,林分更新能力较差,群落相对不稳定。群落中个体的树高与胸径呈正相关关系,物种的径级结构能间接反映垂直分布情况,相比EBSF,EBANF与EBP的中低层乔木个体有缺失,这主要是人为管理与干扰的缘故。从不同径级个体的优势程度看,EBSF中DBH≤10 cm的个体数居多,占72.15%;DBH>10 cm的个体数占27.85%,高于南亚热带季风气候下的广西大青山次生林(21.05%)(农友等,2015)和鼎湖山大样地次生林(16.42%)(叶万辉等,2008),低于 CTFS中热带季雨林区的Mudumalai大样地(58.97%),近似等于HKK大样地(27.1%)(Condit et al.,2000),体现了帽峰山的南亚热带季风气候,同时也反映出其林分向热带过渡的特点。 在物种多样性方面,帽峰山常绿阔叶次生林的平均Shannon-Wiener指数与Pielou均匀度指数值分别为 4.15、0.76。与自然条件下形成的南亚热带常绿阔叶林的 Shannon-Wiener多样性指数(4-5)与均匀度指数(0.7~0.8)(彭少麟,1987;彭少麟,1996)一致,反映出该植被类型典型的近自然性。比张璐等(2003)的帽峰山次生林的物种多样性指数3.30和均匀度指数0.43偏大,说明林分经过10 a演替,群落结构趋于稳定,系统发育更加良好。而常绿阔叶人工-天然混交林与人工林的多样性指数则相对较小。群落的物种多样性是由物种的丰富度与均匀度共同决定的,对比3种植被类型的乔、灌木的物种多样性可知,常绿阔叶人工-天然混交林的乔灌层的物种丰富度相对较低(DG=2.75,2.14)是其多样性偏低的主要原因。该植被类型分布于白云山,地处城市腹地的孤立山地,许多南亚热带树种由于地理阻隔难以进入白云山,故群落中植被丰富度偏小,而白云山地区作为5A级旅游景点,人流量大,人为干扰也会导致植被破坏,物种丰富度减少。常绿阔叶人工林较低的乔灌木物种丰富度以及灌木层较差的均匀度使得物种多样性偏低。该植被类型分布于天河公园,其建群树种主要是人为种植的柠檬桉、白兰花等,物种相对单一,而且地理阻碍与人为干扰使得更新物种多为公园原有物种,如山矾、阴香等。林下优势成分九节、阴香等个体数量过多,据外业调查显示,林下九节、阴香、山矾的密度占林下总密度的89.9%,在竞争中占据绝对优势,影响了其他小灌木的生长,使灌木分布不均匀,林下植被均匀度较差。 一般情况下,由于灌木中存在一定比例的幼树,灌木层植物多样性指标要高于乔木层。本研究帽峰山的常绿阔叶次生林表现出与此一致的结果。而白云山与天河公园的常绿阔叶人工-天然混交林与人工林却表现出相反的结果,这与广州市珠江公园、创业公园、荔湾湖公园等多样性研究(陈雷等,2015)以及厦门城市绿地植被调查(洪志猛,2009)的结果一致。白云山与天河公园地处广州市区,人流量大,人为干扰严重,另外为了维持森林景观的美景度,管理处会定期清除林下的部分植被,这使得林下灌层多样性小于乔木层。 种间关联是群落形成、演替的重要数量和结构特征,也是种间关系和群落分类的依据(岳永杰等,2009),优势种间的关联度能反映种群间的空间依赖性和一定的功能关系(汤孟平等,2006)。广州帽峰山常绿阔叶次生林与白云山常绿阔叶人工-天然混交林主要木本植物间总体呈现不显著的正相关性,且190个优势种对的关联性在χ2检验、PC值、AC值上分别表现出不显著性,这说明这两种植被类型各自的木本植物间呈现相对独立的分布状态。一般情况下,群落在演替初期,物种间的关联程度往往较低,甚至表现为较大的负关联,种间的竞争排斥较为激烈(郭忠玲等,2004)。随着群落演替过程中物种与物种、群落、环境间的相互作用逐渐增强,种间的正关联性会逐渐增大;当进化到顶级阶段时,生态位分化,各物种协调共存,表现出明显的正关联性(胡文强等,2013)。以往研究发现,广州帽峰山次生林乔、灌层主要种群间的联结性分别呈不显著、显著的负关联(林伟强等,2004),本研究中该地区次生林木本植物间呈现不显著的正关联,说明经过11 a的演替,物种间的竞争排斥作用减弱,尤其是灌木层,其密度由 5240株·hm-2下降为1596株·hm-2,正关联性变大,群落结构也趋于复杂化,但现有群落的种间正联结性仍不显著,群落结构及物种组成仍不稳定,与南亚热带的顶级群落(王伯荪等,1985)相比,乔木层物种组成较为单一,更新层缺乏地带性顶级群落建群种。白云山人工-天然混交林的木本植间的正联结性更差,χ2检验的负联结的种对数甚至超过正联结,反映了其林分结构与物种组成的不稳定性,主要是 由于人为扰动导致林下更新层的破坏。 总之,与南亚热带顶级森林群落相比,帽峰山常绿阔叶次生林的乔木层和更新层物种组成相对单一,林下植被密度过大,需要补植一些优良的生态乡土树种和进行一定程度的林下抚育间伐。以帽峰山常绿阔叶次生林为标准,白云山人工-天然混交林的物种组成单一,中小个体数目较少,需要补植一些林下有地带性特色的中小植被,并且对林分进行适当的保护,减少人为破坏;白云山和天河公园人工林的林下灌木层的丰富度与均匀度均较差,且种群分布过于聚集化,某一区域特别密集(可达3867株·hm-2),某一区域特别稀少,需要在稀疏区域补植优良的林下植被,同时对林下密集的九节、阴香、山矾进行一定程度的间伐。森林种间的正联结关系可以作为森林恢复的重要依据,在对森林进行保护、间伐与补植时,可以将正联结性强的树种作为目标树种,以缩短森林演替的时长,全面发挥森林的多效功能(张明霞等,2015)。 ALBERTI M. 2005. The effects of urban patterns on ecosystem function [J]. International regional science review, 28(2): 168-192. ALVEY A A. 2006. Promoting and preserving biodiversity in the urban forest [J]. Urban Forestry & Urban Greening, 5(4): 195-201. ARES A, NEILL A R, PUETTMANN K J. 2010. Understory abundance, species diversity and functional attribute response to thinning in coniferous stands [J]. Forest Ecology and Management, 260(7): 1104-1113. CONDIT R, ASHTON P S, BAKER P, et al. 2000. Spatial patterns in the distribution of tropical tree species [J]. Science, 288(5470): 1414-1418. FISHER J I, MUSTARD J F, VADEBONCOEUR M A. 2006. Green leaf phenology at Landsat resolution: Scaling from the field to the satellite [J]. Remote sensing of environment, 100(2): 265-279. MYEONG S, NOWAK D J, DUGGIN M J. 2006. A temporal analysis of urban forest carbon storage using remote sensing [J]. Remote Sensing of Environment, 101(2): 277-282. NOWAK D J, CRANE D E. 2002. Carbon storage and sequestration by urban trees in the USA [J]. Environmental pollution, 116(3): 381-389. SCHLUTER D. 1984. A variance test for detecting species associations, with some example applications [J]. Ecology, 65(3): 998-1005. TAKEBAYASHI H, MORIYAMA M. 2007. Surface heat budget on green roof and high reflection roof for mitigation of urban heat island [J]. Building and Environment, 42(8): 2971-2979. YATES F E, MARSH D J, IBERALL A S. 1972. Integration of the whole organism: A foundation for a theoretical biology [J]. Challenging biological problems: Directions towards their solution. Oxford Univ. Press, New York: 110-132. 陈雷, 孙冰, 谭广文, 等. 2015. 广州公园植物群落物种组成及多样性研究[J]. 生态科学, 34(5): 38-44. 陈勇, 孙冰, 廖绍波, 等. 2013. 深圳市主要植被群落类型划分及物种多样性研究[J]. 林业科学研究, 26(5): 636-642. 陈玉凯, 杨小波, 李东海, 等. 2011. 海南霸王岭海南油杉群落优势种群的种间联结性研究[J]. 植物科学学报, 29(3): 278-287. 郭连金, 张文辉, 刘国彬. 2007. 黄土丘陵区沙棘人工林发育过程中物种多样性及种间关联变化[J]. 应用生态学报, 18(1): 9-15. 郭垚鑫, 康冰, 李刚, 等. 2011. 小陇山红桦次生林物种组成与立木的点格局分析[J]. 应用生态学报, 22(10): 2574-2580. 郭忠玲, 马元丹, 郑金萍, 等. 2004. 长白山落叶阔叶混交林的物种多样性, 种群空间分布格局及种间关联性研究[J]. 应用生态学报, 15(11) 2013-2018. 洪志猛. 2009. 厦门城市公园植物群落的物种丰富度调查分析[J]. 南京林业大学学报: 自然科学版, 33(2): 51-54. 胡文强, 黄世能, 李家湘, 等. 2013. 南岭石坑崆山顶矮林乔木优势种群的种间关联性[J]. 生态学杂志, 32(10): 2665-2671. 李军玲, 张金屯. 2006. 太行山中段植物群落物种多样性与环境的关系[J]. 应用与环境生物学报, 12(6): 766-771. 李军玲, 张金屯. 2010. 太行山中段植物群落草本植物优势种种间联结性分析[J]. 草业科学, 27(9): 119-123. 廖建华, 陈月华, 覃事妮. 2013. 湖南烈士公园植物群落的观赏效果及生态效益比较分析[J]. 中南林业科技大学学报, 33(8): 143-146. 林伟强, 贾小容, 陈北光, 等. 2006. 广州帽峰山次生林主要种群生态位宽度与重叠研究[J]. 华南农业大学学报, 27(1): 84-87. 林伟强, 刘惠明, 张璐. 2004. 广州帽峰山次生林主要种群种间联结性研究[J]. 生态科学, 23(1): 42-46. 马克平,刘玉明. 1994. 生物群落多样性的测度方法.Ⅰ:α多样性的测度方法(下)[J]. 生物多样性, 2(4): 231-239. 马克平. 1994. 生物群落多样性的测度方法.Ⅰ:α多样性的测度方法(上)[J]. 生物多样性, 2(3): 162-168. 农友, 郑路, 贾宏炎, 等. 2015. 广西大青山次生林的群落特征及主要乔木种群的空间分布格局[J]. 生物多样性, 23(3): 321-331. 彭少麟, 方炜. 1996. 广州白云山次生常绿阔叶林的群落结构动态[J].应用与环境生物学报, 2(1): 22-28. 彭少麟. 1987. 广东亚热带森林群落的生态优势度[J]. 生态学报, 7(1): 36-42. 彭少麟. 1996. 南亚热带森林群落动态学[M]. 北京: 科学出版社: 84, 98-99. 奇凯, 张春雨, 侯继华, 等. 2010. 赤峰市沙地油松林草本植物多样性及种间关联动态[J]. 生态学报, 30(18): 5106-5112. 任国学, 刘金福, 徐道炜, 等. 2011. 戴云山国家级自然保护区黄山松群落类型与物种多样性分析[J]. 植物资源与环境学报, 20(3): 82-88. 邵海荣, 杜建军, 单宏臣, 等. 2006. 用空气负离子浓度对北京地区空气清洁度进行初步评价[J]. 北京林业大学学报, 27(4): 56-59. 史作民, 刘世荣. 2001. 宝天曼落叶阔叶林种间联结性研究[J]. 林业科学, 37(2): 29-35. 苏志尧, 陈北光. 2001. 广州白云山几种森林群落的物种丰富度和多样性[J]. 华南农业大学学报, 22(3): 5-8. 汤孟平, 周国模, 施拥军, 等. 2006. 天目山常绿阔叶林群落最小取样面积与物种多样性[J]. 浙江林学院学报, 23(4): 357-361. 汪殿蓓, 暨淑仪, 陈飞鹏, 等. 2003. 深圳南山区天然森林群落多样性及演替现状[J]. 生态学报, 23(7): 1415-1422. 汪殿蓓, 暨淑仪. 2002. 深圳市南山区山地植被和主要植物群落类型[J].东北林业大学学报, 30(6): 97-101. 王伯荪, 彭少麟. 1985. 南亚热带常绿阔叶林种间联结测定技术研究——Ⅰ.种间联结测式的探讨与修正[J]. 植物生态学报, 9(4): 274-285. 王国梁, 刘国彬. 2002. 黄土丘陵沟壑区铁杆蒿群落种间联结性研究[J].中国草地, 24(3): 1-6. 叶万辉, 曹洪麟, 黄忠良, 等. 2008. 鼎湖山南亚热带常绿阔叶林 20公顷样地群落特征研究[J]. 植物生态学报, 32(2): 274-286. 岳永杰, 余新晓, 李钢铁. 2009. 北京松山自然保护区蒙古栎林的空间结构特征[J]. 应用生态学报, 20(8): 1811-1816. 张璐, 林伟强, 陈北光, 等. 2003. 广州帽峰山次生林群落结构特征[J].华南农业大学学报, 24(3): 53-56. 张明霞, 王得祥, 康冰, 等. 2015. 秦岭华山松天然次生林优势种群的种间联结性[J]. 林业科学, 51(1): 12-21. 张倩媚, 陈北光, 周国逸. 2006. 鼎湖山主要林型优势树种种间联结性的计算方法研究[J]. 华南农业大学学报, 27(1): 79-83. 张永利, 杨锋伟, 王兵, 等. 2010. 中国森林生态系统服务功能研究[M].北京: 科学出版社: 1-2. 庄雪影, 甄荣东. 2002. 增城市主要森林群落植物多样性研究[J]. 林业科学研究, 15(2): 182-189. Structure and Species Diversity of Typical Forests in Guangzhou City ZHAI Shilei, CHEN Bufeng, LIN Na, SHI Xin, PAN Yongjun, ZOU Zhijin 0.48, 0.48 and 0.24 hm2evergreen broad-leaf forests of Maofeng Mountain, Baiyun Mountain and Tianhe park in Guangzhou urban scale were selected respectively. Communities structure, species diversity and interspecific association of dominant populations of three vegetation types were comparative studied by the method of community ecology. The result indicated that species number, stand density and canopy density are characterized by Evergreen Broadleaved Secondary Forest (EBSF) >Evergreen Broadleaved Artificial and Natural Forest (EBANF) >Evergreen Broadleaved Plantation (EBP). The same character was showed on the Shannon-Wiener (SW) index and Gleason index of tree layer, shrub layer and all woody plants. The SW index and Gleason index of shrub layer had a higher value than tree layer in EBSF, and it was converse in EBANF and EBP. While the Pielou evenness index showed a changeable feature. The diameter-class structure of EBSF, EBANF and EBP were recognized as Standard, bimodal and peaky inverse “J” type respectively. 20 dominate populations of woody plants showed insignificant positive association in EBSF and EBANF. The interspecific association of 190 species-pairs were the similar conclusion of insignificant positive association analyzed by χ2 correction test, percentage co-occurring (PC) and association coefficient (AC). Moreover, the interspecific association of EBSF species was more significant. The paper revealed the difference in communities structure, species diversity and main interspecific relevance of three vegetation types in Guangzhou local area, which would provide a good theoretical basis for the construction and evaluation of the near-natural forest and beautiful city forest landscape as a typical example. Guangzhou; urban forest; community structure; species diversity; interspecific association 10.16258/j.cnki.1674-5906.2015.10.006 Q948;X176 A 1674-5906(2015)10-1625-09 翟石磊,陈步峰,林娜,史欣,潘永军,邹志谨. 广州市典型森林植被的结构及多样性研究[J]. 生态环境学报, 2015, 24(10): 1625-1633. ZHAI Shilei, CHEN Bufeng, LIN Na, SHI Xin, PAN Yongjun, ZOU Zhijin. Structure and Species Diversity of Typical Forests in Guangzhou City [J]. Ecology and Environmental Sciences, 2015, 24(10): 1625-1633. 国家林业公益性行业科研专项(201404030105);广州市城市森林生态环境效益监测分析研究项目(2015.01-2015.12);广东珠江三角洲森林生态系统国家定位观测研究站运行补助项目(2015-LYPT-DW-012) 翟石磊(1990年生),男,硕士研究生,主要从事森林生态学研究。E-mail: 18620842880@163.com *通信作者:陈步峰(1958年生),男,研究员,硕士,主要从事森林生态学研究。E-mail: zsjcsdwcbf@126.com 2015-09-20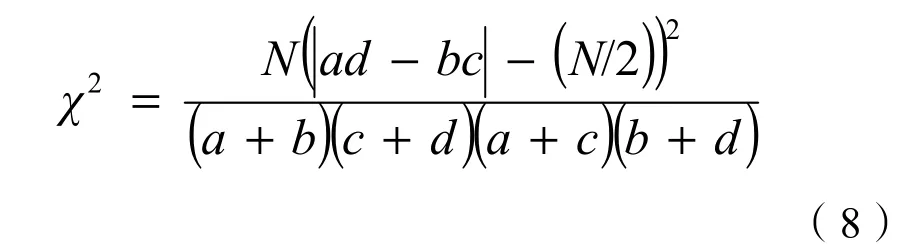




3 结果与分析






4 结论与讨论
Research Institute of Tropical Forestry, Chinese Academy of Forestry, Guangzhou 510520, China
