念青唐古拉山东南坡植被群落数量生态分析及群落多样性
2015-12-08曲广鹏参木友赵景学陈晓鹏张涛白彦福
曲广鹏,参木友,赵景学,陈晓鹏,张涛,白彦福
1. 西藏自治区农牧科学院草业科学研究所,西藏 拉萨 850000;2. 中国科学院青藏高原研究所,北京 100101;3. 中国科学院水利部成都山地灾害与环境研究所,四川 成都 610041;4. 兰州大学草地农业科技学院,甘肃 兰州 730020
念青唐古拉山东南坡植被群落数量生态分析及群落多样性
曲广鹏1*,参木友1,赵景学2,陈晓鹏3,张涛4,白彦福4
1. 西藏自治区农牧科学院草业科学研究所,西藏 拉萨 850000;2. 中国科学院青藏高原研究所,北京 100101;3. 中国科学院水利部成都山地灾害与环境研究所,四川 成都 610041;4. 兰州大学草地农业科技学院,甘肃 兰州 730020
研究沿海拔梯度对植被群落的影响,有助于人们进一步理解未来气候变化背景下,高寒生态系统结构和功能的响应模式。选择念青唐古拉山东南坡的典型生境,包括从海拔4 775 m到5 305 m全部范围内的植物群落。沿着海拔梯度每隔约30 m设置1个样带,并在各个样带上随机设置样方,调查样方内物种数、物种高度、物种盖度、物种频度。采用群落多样性指数、应用双向指示种分析法(TWINSPAN)和除趋势对应分析(DCA)等方法,对西藏念青唐古拉山东南坡高山草甸植物群落进行了分析。TWINSPAN将所有植物群落划分为5种类型,分别为:香柏(Sabina pingii var.wilsonii)+高山嵩草(Kobresia pygmaea)灌丛草甸群落;高山嵩草+圆穗蓼(Polygonum macrophyllum)草甸群落;高山嵩草+矮生嵩草(Kobresia humilis)草甸群落;高山嵩草草甸群落;流石滩冰缘植被群落。这些群落分别位于不同的海拔高度。分类结果很好地反映了植物群落类型分布与地形、海拔的关系,并在DCA二维排序图上得到了较好的验证。念青唐古拉山东南坡草地植物群落多样性在高海拔、低海拔地区较低,中间海拔高度地区较高。植物群落多样性指数呈现出草本层>灌丛层>流石滩冰缘的特征。多样性指数与海拔高度之间的趋势模拟均呈负二次函数关系,单峰式函数关系能较好地表达不同海拔梯度植物群落多样性和均匀度与海拔之间的分布格局。
念青唐古拉山;植被类型;数量生态学;生物多样性;海拔梯度;双向指示种分析法;除趋势对应分析
植物与环境关系是植被生态学研究的重点之一,而分类和排序是研究植物与环境关系的两种重要数量方法,它为客观、准确地揭示植被、植物群落及植物与环境之间的生态关系提供了合理、有效的途径,是国际上植被生态学重要的研究内容之一(张金屯,1991),已被广泛应用于国内的植被生态学研究(张峰等,2000;贾晓妮等,2007;张新时,1991;彭少麟,1988;尚占环等,2005)。生物多样性作为全球变化的重要研究内容之一,是当前国际生态学研究的热点问题。目前,在生物多样性与生态系统功能研究中,草地生态系统备受关注(杨元合等,2004)。全球温度升高已经导致了高纬度及高海拔地区陆地生态系统的结构和功能发生了显著变化。在国际生物圈(IBP)研究计划中,青藏高原被列为全球气候变化的敏感区域,这种极端环境下发育的植被和土壤对气候变化极为敏感,是研究生态系统对气候变化响应与适应机制的天然实验室(王建林等,2009)。
念青唐古拉山脉位于我国西藏自治区,是青藏高原主要山脉之一,属断块山。西接冈底斯山脉,东南延伸与横断山脉伯舒拉岭相接,中部略为向北凸出,同时将西藏划分为藏北、藏南、藏东南三大区域。在海拔较低的暖热地区尚有常绿阔叶林及铁杉林分布,森林带以上则为高山灌丛草甸及高山草甸带,面积较广,是当地主要天然夏季牧场,适宜放养牦牛、绵羊等牲畜。近年来,许多学者就念青唐古拉山陆-气系统水热特征、气候特征、高寒草原生态系统表层土壤有机碳分布特征等方面的研究较多(谢健等,2009;王建林等,2009),而关于念青唐古拉山东南坡植被数量分析及群落多样性
的研究鲜有报道。研究沿海拔梯度的不同水热组合对植被群落的影响,有助于我们进一步理解未来气候变化背景下,高寒生态系统结构和功能的响应模式。基于此,选择念青唐古拉山东南坡植被进行研究,分析沿海拔梯度植被群落分布特征,为进一步了解念青唐古拉山植物群落发展趋势提供理论依据。
1 材料与方法
1.1 研究区域概况
念青唐古拉山脉横跨青藏高原腹地,总体上呈现东北走向,主峰海拔为 7162 m,山顶海拔一般为5600~6200 m;西北侧为纳木错盆地,东南侧为当雄羊八井地堑,构成青藏高原内部的典型盆-山构造地貌,山脉宽度在东北部约为10 km,西南部约35 km。念青唐古拉山脉的山顶面与两侧盆地相对高差大约800~1600 m,属青藏高原内部的高山-极高山地区。本研究选择位于藏北腹地的当雄县城(30°32′N,91°03′E)西北部的念青唐古拉山东南坡的高山植被群落为研究对象,海拔4775~5305 m,属于高原亚寒带季风半干旱气候。据当雄县气象站记载,近40余年(1963─2010年)以来,该地区气温升高明显,降水年纪间差异大。该区多年(1963─2010年)年均气温1.8 ℃,最冷月(1月)均温-10.4 ℃,最热月(7月)均温10.7 ℃。年均降水量479 mm,其中90%集中在5─9月(王忠,2011)。年均湿润度K值为0.7,植被为以寒旱生的高山嵩草为优势种的高山草甸(王建林等,2009)。本地区土壤为高山草甸土,土壤厚度在0.3~1.0 m之间(付刚,2013)。
1.2 植被调查与取样
选择念青唐古拉山东南坡能代表各个生境的典型地区,包括从海拔4775 m到5305 m全部范围内的植物群落,每隔约30 m海拔设置1条样带,共18条。在各个样带上随机设置5~8个样方,草本植被样方为0.25 m×0.25 m,灌丛样方为10 m×20 m,主要调查样方内物种数、物种高度、物种盖度、物种频度,共108个样方。盖度采用针刺法;频度为在每个样地内采用频度圈随机 20次,计数圈内物种的有无。同时记录样地海拔高度、坡向、坡度等地形特征。植被调查工作全部集中在 2010年 9月完成。
1.3 数据处理
数据分析根据张金屯(2004)论述,选择双向指示种分类法(TWINSPAN)和除趋势对应分析排序法(DCA)。采用植被数量分析程序WinTWINS和Canoco分别完成植物群落的TWINSPAN分类和DCA排序分析,计算时以植物重要值为属性进行分类和排序计算。进行排序和分类前先将调查的草本样方84个、灌丛样方24个以植物物种重要值大小进行初步分类,将样方合并,整理出 18个样地植物数据,对 18个样地植物群落进行分类、排序分析,研究18个样地植被的关系和分布特征。
物种多样性指数(Diversity index)、丰富度指数(Richness index)和均匀度指数(Evenness index)的计算公式如下(陈亚明等,2004;李新荣等,2000):
群落多样性指数的运算采用 Shannon-Wiener指数:

Simpson指数:

丰富度指数的运算采用Margalef指数:


重要值=(相对高度+相对盖度+相对频度)/3(5)
式中,S为每个样地的总物种数,N为每个样方中的全部物种的总个体数,Pi为种i的重要值。

图1 念青唐古拉山东南坡植物群落TWNSPAN分类结果树状示意图Fig. 1 Dendrogram of florae in the southeast slope of Mountain Nyenquentanglha by TWINSPAN
均匀度指数的运算采用Pielou指数:
2 结果与分析
2.1 TWINSPAN分类结果与群落类型概况
双向指示种分类法(TWINSPAN)的第1次划分将所有样地分为2类(图1),一类是流石滩冰
缘植被群落,包括4个样地;另一类为高寒草甸和高山灌丛植被群落包括 14个样地,逐步将样方继续分为6组,结合山地地形、植被分布的土壤生境的生态特征,在TWINSPAN基础上将该山地植被分为5组。各组物种优势度如表1。
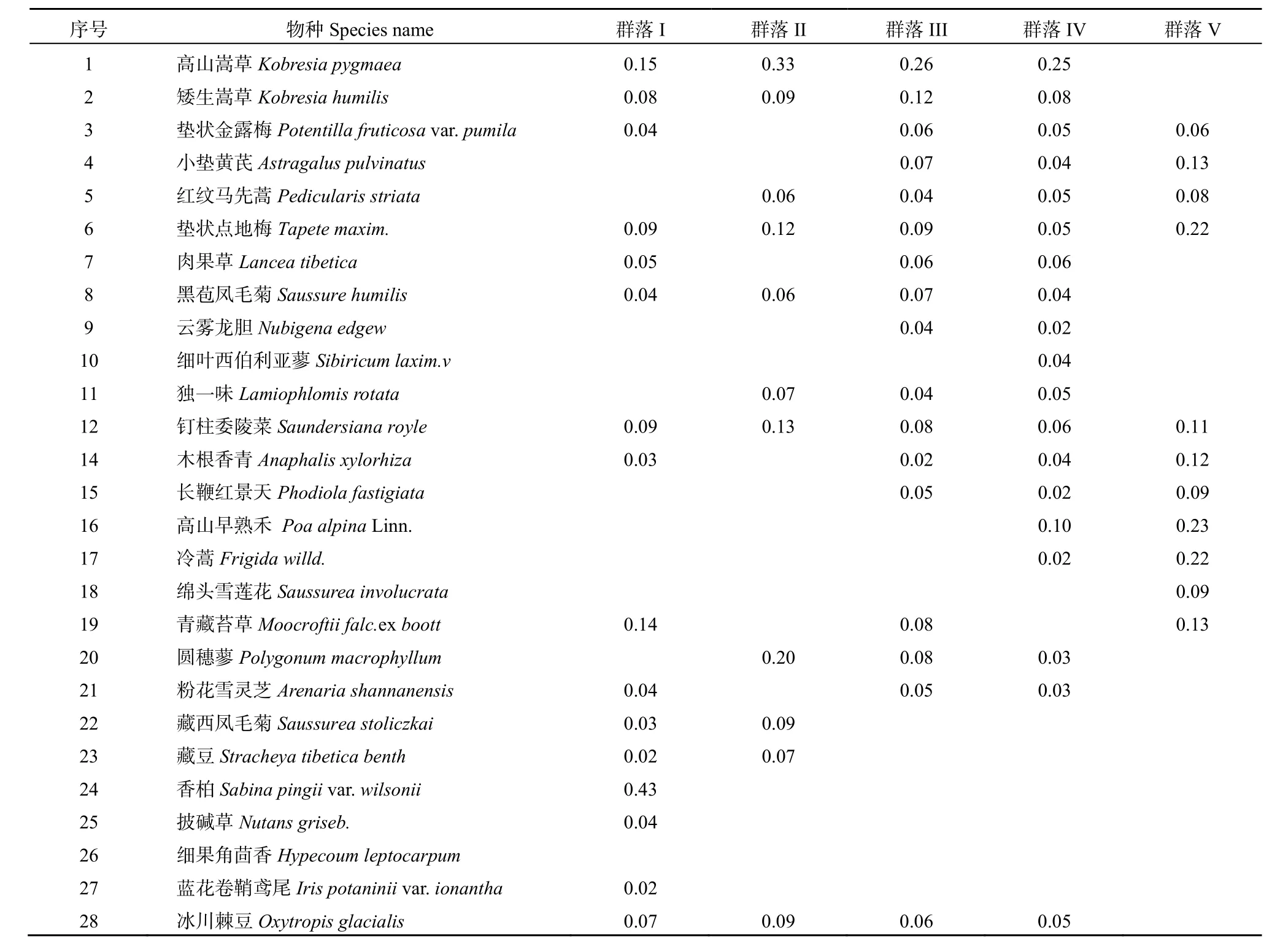
表1 念青唐古拉山东南坡主要植被种类重要值Table 1 Species importance value of major vegetation in the southeast slope of Mountain Nyenquentanglha
I香柏(Sabina pingii var.wilsonii)+高山嵩草(Kobresia pygmaea)灌丛草甸群落。该群落分布于海拔4775~4889 m,包含样地1~5。优势植物有香柏和高山嵩草,优势度分别为0.43和0.15。伴生植物主要有青藏苔草(Moocroftii falc.ex boott)、矮生嵩草(Kobresia humilis)、钉柱委陵菜(Saundersiana royle)冰川棘豆(Oxytropis glacialis)、木根香青(Anaphalis xylorhiza)和粉花雪灵芝(Arenaria shannanensis)等。
II 高山嵩草+圆穗蓼(Polygonum macrophyllum)草甸群落。该群落分布于海拔4996~5050 m,包含样地8、9和10。优势种为高山嵩草和圆穗蓼,优势度分别为 0.33和 0.20。伴生植物主要有矮生嵩草、藏西风毛菊(Saussurea stoliczkai)、独一味(Lamiophlomis rotata)、钉柱委陵菜、冰川棘豆等。
III高山嵩草+矮生嵩草草甸群落。该群落分布于海拔4945~5163 m,包含样地6、7和样地11、12。优势植被为高山嵩草和矮生嵩草,优势度分别为0.26和0.12。伴生植物主要有垫状点地梅、肉果草(Lancea tibetica)、钉柱委陵菜、圆穗蓼等。
IV高山嵩草草甸群落。该群落分布于海拔5163~5196 m,包含样地13和14。高山嵩草为该群落的单优势种,优势度为0.25。伴生植被有矮生嵩草、高山早熟禾(Poa alpina Linn.)、垫状金露梅(Potentilla fruticosa var. pumila)、红纹马先蒿(Pedicularis striata)等。
V流石滩冰缘植被群落。该群落分布于海拔5238~5305 m,包含样地15~18。高山早熟禾和垫状棱点地梅(Tapete maxim.)为该群落的优势种,优势度分别为0.23和0.22。伴生植被有冷蒿、雪莲、青藏苔草、木根香青等。
2.2 草甸群落的DCA排序
应用除趋势对应分析排序法(DCA)对念青唐古拉山东南坡高山草甸 18个样地进行分析,为了
便于和 TWINSPAN分类结果进行比较,将TWINSPAN的最终分组结果在DCA二维轴上标出(Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ)(图2)。从该图我们也可以看出TWINSPAN分类结果和DCA排序结果较吻合。DCA第一轴表现为海拔高度和土壤质地,第二轴表现为坡度和土壤水分。按第一轴群落顺序,从左到右,随着海拔增加,土质逐渐变差,植被群落逐渐从Ⅰ转变为Ⅴ。DCA第二轴群落顺序,从下到上,随着坡度逐渐增加,土壤水分降低,植被群落逐渐从Ⅰ转变为Ⅳ。从上述分析可见,DCA第一轴和第二轴有密切关系,草甸群落的分布明显受海拔、坡度、土壤水分、土壤质地等环境因子的制约,说明DCA排序较好地反映了念青唐古拉山东南坡高山草甸群落的生态关系。
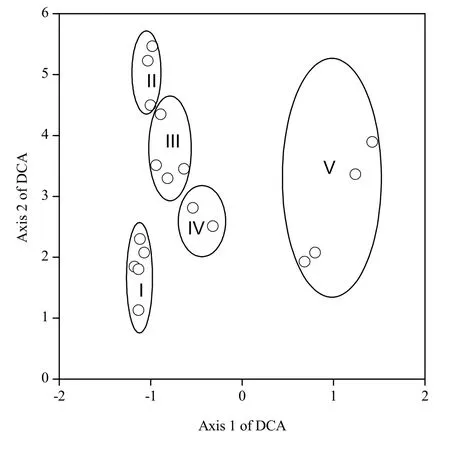
图2 研究区内植物群落的DCA二维排序Fig. 2 Two-dimensional ordiration of the florae in the region by the DCA
2.3 植被群落多样性分析
念青唐古拉山东南坡不同植物群落类型的多样性程度是综合生态因素作用的结果,其中海拔最具重要性。海拔以及由于海拔造成的水分、热量条件的差异是影响环境因子变化的主导因素,从而导致不同类型植物群落的物种多样性也发生了很大的变化(表2)。Shannon-Wiener指数,群落Ⅳ最高,群落Ⅱ、Ⅲ次之,群落Ⅰ较低,群落Ⅴ最低。其中,群落Ⅴ与其他群落差异均显著(P<0.05);群落Ⅳ与群落Ⅰ和Ⅱ差异显著(P<0.05),与群落Ⅲ差异不显著(P>0.05);群落Ⅰ、Ⅱ、Ⅲ之间差异不显著(P>0.05)。Simpson指数,群落Ⅳ最高,群落Ⅰ、Ⅱ、Ⅲ次之,群落Ⅴ最低。其中,群落Ⅴ与其他群落差异均显著(P<0.05);群落Ⅳ与群落Ⅰ差异显著(P<0.05),与群落Ⅱ、Ⅲ差异不显著(P>0.05);群落Ⅰ、Ⅱ、Ⅲ之间差异不显著(P>0.05)。Margalef指数,群落Ⅳ最高,群落Ⅰ、Ⅲ次之,群落Ⅴ较低,群落Ⅱ最低。其中,群落Ⅴ与群落Ⅲ之间差异显著(P<0.05),与其他群落差异不显著(P>0.05);群落Ⅳ与群落Ⅰ、Ⅱ、Ⅲ差异均显著(P<0.05);群落Ⅰ与群落Ⅱ、Ⅲ差异不显著(P>0.05);群落Ⅱ和群落Ⅲ之间差异显著(P<0.05)。Pielou指数,群落Ⅲ最高,群落Ⅲ次之,群落Ⅰ、Ⅳ较低,群落Ⅴ最低。其中,群落Ⅴ与群落Ⅰ、Ⅳ差异不显著(P>0.05),与群落Ⅱ、Ⅲ差异显著(P<0.05);群落Ⅳ与群落Ⅰ、Ⅱ、Ⅲ差异不显著(P>0.05);群落Ⅱ与群落Ⅰ、Ⅲ差异显著(P<0.05);群落Ⅰ、Ⅲ之间差异不显著(P>0.05)。

表2 念青唐古拉山东南坡植被群落多样性指数Table 2 Community diversity index of the vegetation in southeast slope of Mountain Nyenquentanglha
在每个海拔梯度,选择每个样带对Shannon-Wiener指数、Simpson指数、Margalef指数、Pielou指数与海拔进行回归分析(图 3)。结果表明:Shannon-Wiener指数、Simpson指数、Margalef指数、Pielou指数随海拔高度变化,在一定程度上呈现出高海拔和低海拔地区较低,中海拔高度地区较高的趋势。多样性指数与海拔高度之间的趋势模拟均呈负二次函数关系,除与Margalef指数拟合系数较小外,其他各多样性指数与海拔高度的拟合系数均能较好的解释二者之间的关系。说明单峰式函数关系能较好地表达不同海拔梯度植物群落多样性和均匀度与海拔之间的分布格局。植被多样性指数呈现出草本层>灌丛层>流石滩冰缘植被的特征(表2)。
3 讨论
我国学者从20世纪70年代后期引入数量分类和排序技术,开始了植被数量生态学的研究,但大多数研究集中在森林与草原植被中,而在高山-极高
山地区植被研究中应用很少。我们运用TWINSPAN和DCA方法对念青唐古拉山东南坡高山草甸进行数量分类和排序,取得到了较好的结果。

图3 念青唐古拉山东南坡植物群落物种多样性与海拔的关系Fig. 3 Relationship between community diversity and altitude of the southeast slope of Mountain Nyenquentanglha
TWINSPAN等级分类将念青唐古拉山东南坡高山草甸18个样地29个种划分为5个植物群落类型,其分类矩阵图反映出明显的综合环境梯度,即海拔高度、土壤水分、温度梯度,比较客观地反映了植物群落类型及优势种的分布与环境因子之间的关系,并在DCA二维排序图上得到了较好的验证。DCA排序表明群落生境所在地的海拔高度和水热条件是决定群落类型分布的主要因素(Wang et al.,2013;Li et al.,2013)。第一轴反映了群落海拔高度的梯度变化,第二轴反映了土壤水热组合的变化。一般来说,随着山地迎风坡海拔高度逐渐升高,温度逐渐降低,而降水逐渐增加(王忠,2011;李瑞成,2012)。因此,高海拔地区土壤湿度相对于低海拔地区要高。然而,随着海拔和湿度等的变化,念青唐古拉山东南坡高山草甸植物种类和植被高度发生明显变化。在海拔较低的山坡上,香柏等灌木为优势种。随着海拔增加,植被高度明显下降,高山嵩草生长茂盛,逐渐成为优势种。尤其在海拔较高的流石滩山坡上,垫状点地梅等一些垫状植被优势度显著增加,逐渐成为优势种。这是高寒植被对恶劣环境的一种适应机制,具有对恶劣环境的指示作用,也是高寒垫状植被生态系统工程师效应的体现(何永涛等,2010;李瑞成,2012)。同时,高低海拔优势群落的分布不同,也指示了高低海拔可能存在迥然不同的环境限制因子。低海拔较低的降水,更适宜根系粗大的灌木生长。然而,低温也是垫状植被普遍分布在于高海拔地区的原因。
念青唐古拉山坡降水量海拔梯度效应明显,随海拔梯度的升高而增加,高海拔降水过程和低海拔差异显著,最大降水高度位于山坡上部的 5100 m附近(谢健等,2009;王忠,2011;李瑞成,2012)。然而,念青唐古拉山坡草地群落,在降水量随海拔的升高而增加的同时,土壤中砂粒含量则逐渐增加,相应的粉粒和粘粒含量逐渐下降(Ohtsuka et al.,2008)。海拔以及由于海拔造成的水分、热量条件的差异是影响环境因子变化的主导因素,导致不同类型植物群落的物种多样性也发生了很大的变化(王忠,2011;李瑞成,2012)。关于高寒草甸群落植物多样性沿海拔梯度变化的研究表明,单峰式函数能较好地表达不同海拔梯度植物群落物种丰富度、多样性和均匀度与海拔间的分布格局,中海拔区植物群落多样性最大(王长庭等,2004;赵新全,2009)。Wang et al.(2013)研究发现,生产力在海拔梯度上的单峰分布是由于在不同海拔收到的限制因子不同而造成的。在念青唐古拉山
东南坡,水分导致的干旱胁迫是低海拔的主要限制因子,低温导致的水分和养份胁迫则是高海拔的主要限制因子。当研究区域处在干旱半干旱地区,海拔高度达到植物群落分布的上限时,上述因子的转变造成青藏高原地区高山植被群落分布的单峰分布格局应该是普遍存在的(王忠,2011;李瑞成,2012)。此外,海拔高度的变化也会改变了放牧强度,从而也会影响群落多样性的海拔分布(候扶江等,2006)。然而,随着海拔增加,放牧强度逐渐减弱(Hirota et al.,2009;Moriyama et al.,2013)。这也是中高海拔群落多样性相对较高的可能原因。本文研究发现,念青唐古拉山东南坡高山植被群落Shannon-Wiener指数、Simpson指数、Margalef指数、Pielou指数随海拔高度变化,在一定程度上也呈现出高海拔和低海拔地区较低,中海拔高度地区较高的趋势。多样性指数与海拔高度之间的趋势模拟均呈负二次函数关系,除与Margalef指数拟合系数较小外,其他各多样性指数与海拔高度的拟合系数均能较好的解释两者之间的关系。说明单峰式函数关系能较好地表达不同海拔梯度植物群落多样性和均匀度与海拔之间的分布格局。
4 结果
我们采用群落多样性指数、应用双向指示种分析法(TWINSPAN)和除趋势对应分析(DCA)等方法,对西藏念青唐古拉山东南坡高山草甸植物群落进行了分析。(1)TWINSPAN将所有植物群落划分为 5种类型,分别为:香柏+高山嵩草灌丛草甸群落、高山嵩草+圆穗蓼草甸群落、高山嵩草+圆穗蓼草甸群落、高山嵩草+矮生嵩草草甸群落、高山嵩草草甸群落和流石滩冰缘植被群落。这些群落分别位于不同的海拔高度。分类结果很好地反映了植物群落类型分布与地形、海拔的关系,并在DCA二维排序图上得到了较好的验证。(2)念青唐古拉山东南坡草地植物群落多样性在高海拔、低海拔地区较低,中间海拔高度地区较高。植物群落多样性指数呈现出草本层>灌丛层>流石滩冰缘的特征。(3)多样性指数与海拔高度之间的趋势模拟均呈负二次函数关系,单峰式函数关系能较好地表达不同海拔梯度植物群落多样性和均匀度与海拔之间的分布格局。
HIROTA M, ZHANG P, GU S, et al. 2009. Altitudinal variation of ecosystem CO2fluxes in an alpine grassland from 3600 to 4200 m. Journal of Plant Ecology, 2(4): 197-205.
LI R, LUO T, TANG Y, et al. 2013. The altitudinal distribution center of a widespread cushion species is related to an aptimum combination of temperature and precipitation in the central Tibetan Plateau [J]. Journal of Arid Environments, 88(1): 70-77.
MORIYAMA A, YONEMURA S, KAWASHIMA S, et al. 2013. Environmental indicators for estimating the potential soil respiration rate in alpine zone [J]. Ecological Indicators, 32(9): 245-252.
OHTSUKAT, HIROTA M, ZHANG X, et al. 2008. Soil organi carbon pools in alpine to nival zones along an altitudinal gradient (4400~5300 m) on the Tibetan Plateau [J]. Polar Sciene, 2(4): 277-285.
WANG Z , LUO T, LI R, et al. 2013. Causes for the unimodal pattern of biomass and productivity in alpine grasslands along a large altitudinal gradient in semi-arid regions [J]. Journal of Vegetation Science, 24(1): 189-201.
陈亚明, 李自珍, 杜国祯. 2004. 施肥对高寒草甸植物多样性和经济类群的影响[J]. 西北植物学报, 24(3): 424-429.
付刚. 2013. 不同海拔高寒草甸生态系统呼吸对实验增温和刈割的响应[D]. 北京: 中国科学院研究生院: 10-20.
何永涛, 石培礼, 闫巍. 2010. 高山垫状植物的生态系统工程师效应研究进展[J]. 生态学杂志, 29(6): 1221-1227.
候扶江, 杨中艺. 2006. 放牧对草地的作用[J]. 生态学报, 26(1): 244-264.
贾晓妮, 程积民, 万惠娥. 2007. DCA、CCA和DCCA三种排序方法在中国草地植被群落中的应用现状[J]. 中国农学通报, 23(12): 391-395.
李瑞成. 2012. 念青唐古拉山垫状点地梅沿海拔梯度的分布、物候和生存率格局及其成因[D]. 北京: 中国科学院研究生院: 14-20.
李新荣, 张景光, 刘立超, 等. 2000. 我国干旱沙漠地区人工植被与环境演变过程中植物多样性的研究[J]. 植物生态学报, 24(3): 257-261.
彭少麟. 1988. 广东亚热带部分森林群落排序分析[J]. 武汉植物学研究, 6(1): 37-44.
尚占环, 姚爱兴, 龙瑞军, 等. 2005. 中卫山羊核心产地植物群落的数量分类与排序[J]. 西北植物学报, 25(5): 985-990.
王建林, 欧阳华, 王忠红, 等. 2009. 念青唐古拉山东南坡高寒草原生态系统表层土壤有机碳分布特征及影响因素[J]. 地理科学, 29(3): 385-390.
王建林, 欧阳华, 王忠红, 等. 2010. 念青唐古拉山东南坡高寒草原生态系统表层土壤活性有机碳的影响因素研究[J]. 中国生态农业学报, 18(2): 235-240.
王长庭, 王启基, 龙瑞军, 等. 2004. 高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究[J]. 植物生态学报, 28(2): 240-245.
王忠. 2011. 念青唐古拉山南坡高寒草甸生产力沿海拔梯度的变化机理研究[D]. 北京: 中国科学院研究生院: 12-24.
谢健, 刘景时, 杜明远, 等. 2009. 念青唐古拉山南坡夏季降水梯度观测[J]. 水资源与水工程学报, 20(5): 14-18.
谢健, 刘景时, 杜明远, 等. 2010. 念青唐古拉山南坡气温分布及其垂直梯度[J]. 地理科学, 30(2): 113-118.
谢健, 刘景时, 杜明远, 等. 2010. 念青唐古拉山西段高海拔陆-气系统水热特征[J]. 地理科学进展, 29(2): 151-158.
杨元合, 饶胜, 胡会峰, 等. 2004. 青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系[J]. 生物多性, 12(1): 200-205.
张峰, 张金屯. 2000. 我国植被数量分类和排序研究进展[J]. 山西大学学报(自然科学版), 23(3): 278-282.
张金屯. 1991. 植被数量分类和排序的发展[J]. 山西大学学报(自然科学版), 14(2): 215-224.
张金屯.2004. 数量生态学[M]. 北京: 科学出版社: 1-70.
张新时. 1991. 西藏阿里植物群落的间接梯度分析、数量分类与环境解释[J]. 植物生态学与地植物学学报, 15(2): 101-113.
赵新全. 2004. 高寒草甸生态系统与全球变化[M]. 科学出版社: 106-111.
Quantitative Ecology and Species Diversity of the Vegetation in Southeast Slope of the Nyenchentanglha Mountain
QU Guangpeng1, CAN Muyou1, ZHAO Jingxue2, CHEN Xiaopeng3, ZHANG Tao4, BAI Yanfu4
1. Grassland Institute of Tibet Academy of Agricultural and Animal Husbandry Sciences, Lahsa 850000 China; 2. Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing 100101 China; 3. Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China; 4. College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou 730020 China.
Study on the response of vegetation to altitude gradient is helpful for us to understand how alpine ecosystems will response to future climate change. We chose to study the vegetation along the south slope of the Nyenchentanglha Mountain, with the altitude gradient from 4 775 m to 5 305 m. Along the altitude gradient, samples were set every 30 meters and species, with plant height, coverage and species frequency investigated in each samples. Double gaps indicator species analysis (TWINSPAN), and Detrended correspondence analysis (DCA) were applied to analysis the community types and species diversity along the altitude gradient of the southeast slope of the Nyenchentanglha Mountain. TWINSPAN divided the plant communication into 5 types: Sabina pingii var. wilsonii + Kobresia pygmaea meadow community, Kobresia pygmaea + Polygonum macrophyllum meadow community, Kobresia pygmaea + Kobresia humilis meadow community, Kobresia pygmaea community and stone beach periglacial vegetation communities. The result of vegetation’s classification by TWINSPAN reflected the relation between community’s location, topography and altitude, which were verified in pattern map of DCA very well. The species was lower in high and low altitude area compared with the middle of slope of mountain, which formed a hump cure. The species diversity index from high to low of community was herb layer >shrub layer > periglacial stone-community. The model of diversity index and altitude was negatively quadratic function.
Nyenchentanglha Mountain; vegetation types; Quantitative ecology; biodiversity; altitude variation; TWINSPAN, DCA
10.16258/j.cnki.1674-5906.2015.10.005
Q948;X176
A
1674-5906(2015)10-1618-07
曲广鹏,参木友,赵景学,陈晓鹏,张涛,白彦福. 念青唐古拉山东南坡植被群落数量生态分析及群落多样性[J].生态环境学报, 2015, 24(10): 1618-1624.
QU Guangpeng, CAN Muyou, ZHAO Jingxue, CHEN Xiaopeng, ZHANG Tao, BAI Yanfu. Quantitative Ecology and Species Diversity of the Vegetation in Southeast Slope of the Nyenchentanglha Mountain [J]. Ecology and Environmental Sciences, 2015, 24(10): 1618-1624.
“十二五”国家科技支撑项目;西藏牧区“生产生态生活”保障技术集成与示范(2012BAD13B04)
曲广鹏(1981年生),男,助理研究员,硕士,主要从事牧草栽培与草地生态工作。*通信作者。
2014-08-28
