拟南芥和小菜蛾相互作用的差异蛋白分析
2015-12-08张静范娟
张静,范娟
(黑龙江省蚕业研究所,哈尔滨 150086)
拟南芥和小菜蛾相互作用的差异蛋白分析
张静,范娟
(黑龙江省蚕业研究所,哈尔滨 150086)
以模式植物拟南芥(Arabidopsis thaliana)和专食性害虫小菜蛾(Plutella xylostella)为研究系统,运用蛋白质组学方法分析了拟南芥在小菜蛾取食诱导条件下叶片蛋白的差异表达情况。结果表明:有52个蛋白点在小菜蛾取食诱导条件下发生显著变化,其中28个蛋白点表达量上调,24个蛋白点表达量下调。运用质谱技术对其中的部分差异蛋白进行了鉴定,发现富含甘氨酸的RNA结合蛋白,3-磷酸甘油醛脱氢酶,ATP合酶的α亚基和β亚基在植物诱导防御中具有重要的作用,为揭示植物在昆虫取食条件下的化学防御机制提供蛋白质水平方面的理论基础。
小菜蛾;拟南芥;双向电泳;质谱;诱导防御
植物在漫长的进化过程中形成了重要的防御机制,而昆虫取食诱导的防御机制是植物防御植食者保护自身的主要机制[1]。在昆虫取食诱导条件下的植物机体自身可以合成次生代谢物质来达到防御目的。诱导产生的物质有萜类、烟碱和酚类等次生化学物质,使植物增强抗性[2]。昆虫取食也可诱导相关抗虫基因的表达,引起一系列防御蛋白发生变化。鉴于此,研究昆虫取食诱导植物的抗虫性,既要研究相关抗虫基因,还要对虫害应答蛋白的表达变化进行探究。
小菜蛾(Plutella xylostella)是专食性昆虫(specialist),咀嚼式口器,以植物的叶片为食,是十字花科植物最重要的害虫之一[3]。小菜蛾是世界性破坏性害虫,其寄主多达40种以上,年发生的世代多,繁殖系数较高,且世代重叠的现象严重,最早发生于地中海地区,后来在我国热带地区的危害尤为猖獗,北方地区也呈加重趋势。以幼虫取食叶片,严重时叶片被食成网状,降低食用价值,影响经济效益。拟南芥是一种十字花科植物,广泛用于植物遗传学、发育生物学和分子生物学的研究,已成为一种典型的模式植物,该植物具有个体小、生长周期快、形态特征简单、生命力强、基因组小等优点。研究表明拟南芥是
植物诱导防御机制研究的模式材料,其与小菜蛾互作系统被证明是研究植物和昆虫相互作用的可靠模型[4]。
研究以模式植物拟南芥(Arabidopsis thaliana)和专食性害虫小菜蛾(Plutella xylostella)为植物与昆虫互作体系,运用差异蛋白质组学的方法,解析相关虫害应答蛋白的功能,为利用植物自身防御体系实现有效控制农业害虫提供理论方面的科学依据。
1 材料和方法
1.1 试验材料与处理
拟南芥(Arabidopsis thanliana,Col-0)养植方法:在MS培养基中对种子进行培养,发芽后一周将幼苗转移至营养土和蛭石(比例约为2∶1)中,培养3周,温度为白天22℃,夜晚20℃,光强度80 μE·s-1·m-2,相对湿度为75%,光暗比为16 L∶8 D。
小菜蛾的饲养是以白菜幼苗喂食,3龄幼虫作为供试昆虫。一头幼虫放在一株拟南芥叶片上进行取食,8 h后,移除幼虫收集拟南芥植株的样品[5]。
空白对照组:用MilliQ级纯水喷洒拟南芥植株,8 h后收集植株。
样品收集是幼虫取食后,移除幼虫剩下的全部植株叶片作为一个样品,三次重复。
1.2 蛋白质样品的制备
样品蛋白质的提取运用饱和酚抽提法[6]。蛋白质裂解液为(7 M尿素,2 M硫脲,2%IPG Buffer,2% CHAPS,2%SB3-10,20 mM DTT,5 mM TCEP,0.002%溴酚蓝)。采用Bradford法测定蛋白浓度[7]。
1.3 双向电泳
1.3.1 等电聚焦(IEF)
试验所用仪器为Ettan DALT双向电泳仪。蛋白质的上样量为300 μg,将蛋白质样液连续的加入在水化槽中。剥开固相pH梯度(IPG)胶条(pH 4-7)保护膜后,胶面朝下放入水化槽中(矿物油覆盖),室温水化16 h,然后进行等电聚焦[8]。
1.3.2 胶条的平衡
第一向等电聚焦结束后进行IPG胶条的平衡,IPG胶条分别在平衡液Ⅰ和Ⅱ中平衡15 min[9],然后进行垂直电泳。
1.3.3 聚丙烯酰胺凝胶电泳
胶条平衡后,进行聚丙烯酰胺凝胶电泳,凝胶浓度为12.5%,恒流电泳到凝胶前沿约0.5 cm时终止。试验重复6次。
1.3.4 银染及凝胶图像分析
蛋白凝胶染色方法根据Blum等[10]的报道,利用ImageMasterTM2D Platinum 6.0软件分析,步骤按照差异蛋白点检测、凝胶图谱标准化处理、蛋白质点匹配和生物统计进行。制备胶用考马斯亮蓝方法进行染色。1.3.5差异蛋白质点的质谱鉴定
从制备胶上切取差异蛋白质点,酶解采用Hughes等[11]报道的方法进行,利用胰蛋白酶进行酶解。利用4700型MALDI-TOF-TOF质谱仪进行蛋白鉴定,具体参照Zhang等[12]报道的方法。
2 结果与分析
2.1 拟南芥蛋白(小菜蛾取食诱导条件下)双向电泳
通过ImageMasterTM2D Platinum 6.0软件分析,在蛋白凝胶上,重复性较好,平均有652个蛋白点(图1)。
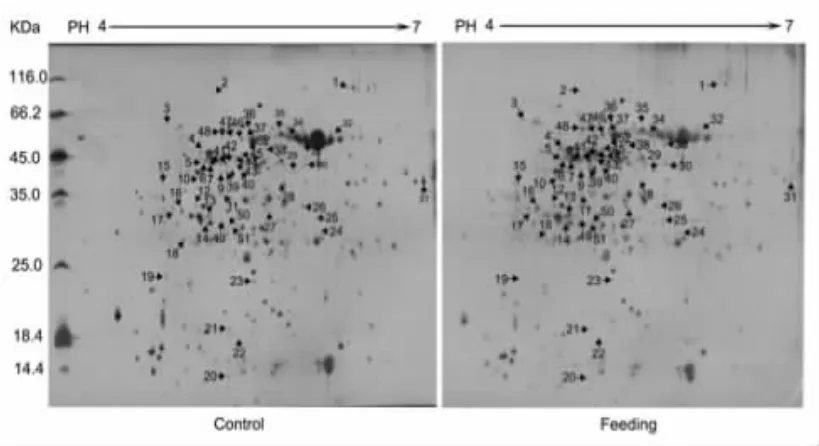
图1 小菜蛾取食诱导条件下拟南芥叶片蛋白质2-DE图谱Fig.1Proteomic 2-DE maps of Arabidopsis thaliana under the induced leaf of Plutella xylostella
通过软件对凝胶图谱进行分析,发现在小菜蛾取食拟南芥的处理组中蛋白与对照相比具有显著性的差异,以蛋白质表达丰度变化在3倍以上为检测阈值,结果有52个蛋白质点发生了变化,其中有28个蛋白质点表达量上调,24个蛋白质点表达量下调,如图1所示,代表性差异蛋白点见图2。
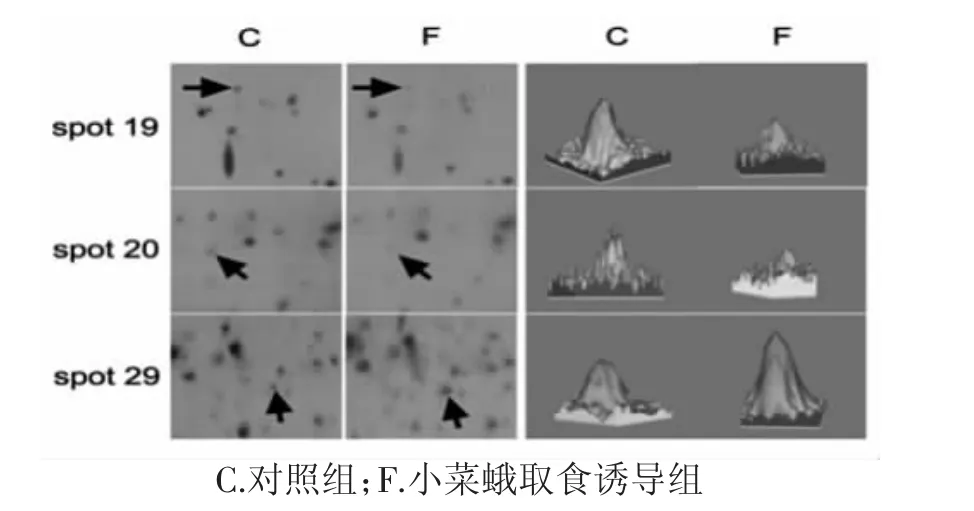
图2 不同处理条件下拟南芥蛋白点的丰度变化Fig.2Abundance changes of Arabidopsis thanliana protein spots under different stress
2.2 差异蛋白质点的质谱鉴定
通过软件分析不同处理条件下双向电泳图谱的匹配情况,运用MALDI-TOF/TOF MS技术对部分差异蛋白点进行了检测,采用Matrix science网站提供的Mascot检索程序进行检索,分析数据结果见表1。
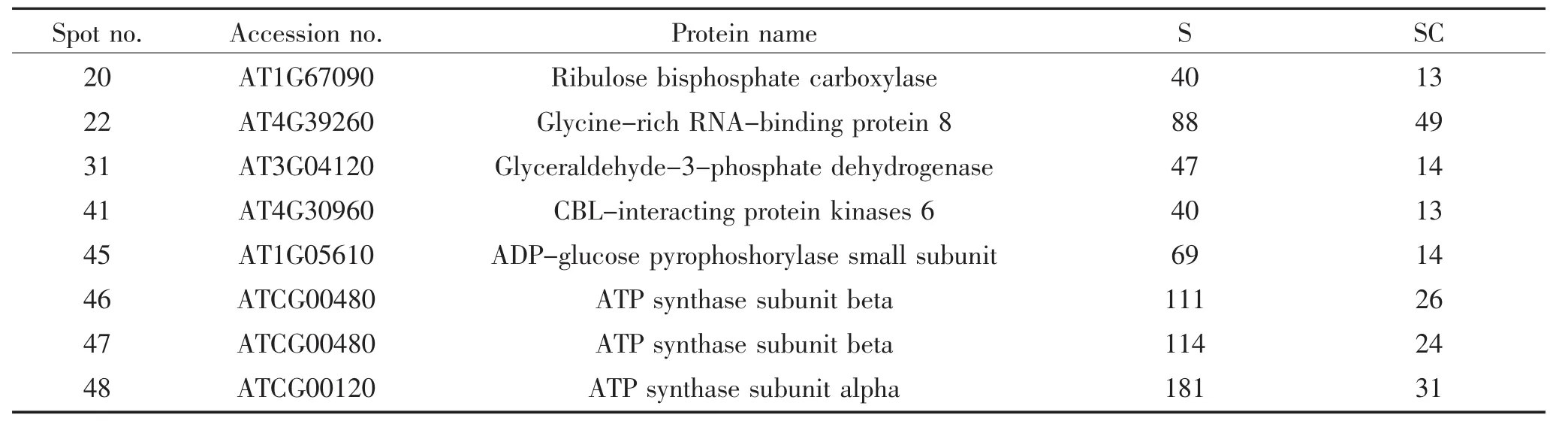
表1 拟南芥在小菜蛾取食诱导条件下差异蛋白的质谱鉴定Table 1Identification of the differentially expressed proteins in Arabidopsis under the induced leaf of Plutella xylostella
通过质谱鉴定得到的虫害应答蛋白包括:核酮糖二磷酸羧化酶(Ribulose bisphosphate carboxylase,Spot 20)、富含甘氨酸的RNA结合蛋白8(Glycinerich RNA-binding protein 8,Spot 22)、3-磷酸甘油醛脱氢酶(Glyceraldehyde-3-phosphate dehydrogenase,Spot 31)、CBL互作蛋白激酶6(CBL-interacting protein kinases 6,Spot 41)、ADP-葡萄糖焦磷酸化酶小亚基(ADP-Glucose pyrophosphorylase small subunit,Spot 45)、ATP合酶beta亚基(ATP synthase subunit beta,Spot 46 and Spot 47)和ATP合酶alpha亚基(ATP synthase subunit alpha,Spot 48)。
3 讨论与结论
随着拟南芥等模式植物基因组序列测定工作的完成,为蛋白功能的探究创造了条件。植物对昆虫取食应答不仅在基因水平上发生变化,在蛋白质表达方面也有一定的影响变化。差异蛋白质组一直是蛋白质组学研究的热点,其核心在于找到引起不同样本之间蛋白质组差异的影响因子,揭开和证明蛋白质组的变化[13]。试验中,在小菜蛾取食诱导条件下有52个表达发生明显变化的蛋白点,实现了我们预期的结果。在昆虫取食条件下,拟南芥的蛋白质表达情况不仅是数量和种类的变化,其基因表达的动态性和网络性存在着一定的复杂性。原因是不同的环境条件下,蛋白质的存在状态和所具有的特性都有很大的差别,这种差别体现在量的变化、合成和降解的速度以及翻译后的修饰等[14]。
富含甘氨酸RNA结合蛋白(表达量下调)是植物中重要的转录调控蛋白,具有C末端的RNA识别基序(RNA-recognition motifs,RRM),以及N末端的甘氨酸富集区域。研究表明RNA结合活性所必需的为C末端的甘氨酸富集区域[15]。有报道机械损伤等处理条件下的植物,体内编码GRP基因的mRNA含量增加,胁迫的条件下GRP的含量变化尤其显著,会迅速积累到很高的水平,提高植物应对胁迫的能力[16]。GRP基因的表达有利于木质素的形成,使植物细胞壁相关结构增厚,植物木质素的形成有利于对逆境的适应,增强植物的抗虫性[17]。
3-磷酸甘油醛脱氢酶(表达量上调)是高等植物卡尔文循环的关键酶,在糖酵解过程中发挥的作用尤其重要[18]。GAPDH基因是一种重要的抗逆胁迫基因,在机体应答伤害信号时发挥显著作用[19],植物在面对害虫取食诱导后,体内的GAPDH基因通过翻译调控来改变机体的代谢平衡进行防御[20]。在小菜蛾取食拟南芥后,植物GAPDH蛋白表达量发生了显著的变化,可以推断3-磷酸甘油醛脱氢酶在拟南芥的诱导防御反应中起着重要的作用,具体的调控机制有待于探究。
ATP合酶(表达量上调)又称H+-ATP酶或F1F0-ATP酶,是参与能量代谢的关键酶,广泛存在于叶绿体和线粒体中,是光合作用不可缺少的重要酶类。ATP合酶的β亚基为其催化亚基,参与呼吸链的电子传递。研究报道红树在盐胁迫的情况下ATP合酶β亚基的表达量增加,光合作用的增强,促进红树耐盐的能力提高[21]。在小菜蛾取食胁迫下,体内的ATP合酶β亚基的表达量增加,我们推测植物光合作用
的强度也在不同程度上发生改变,在受到外界伤害后需要启动各种防御反应来保护机体,而这些反应的过程需要消耗额外的能量来应对昆虫的取食。
在昆虫取食诱导条件下,与三羧酸循环相关的核酮糖二磷酸羧化酶(Spot 20)和3-磷酸甘油醛脱氢酶(Spot 31)、ADP-葡萄糖焦磷酸化酶小亚基(Spot 45)发生了显著的变化。研究表明,烟草在专食性与多食性害虫取食24 h后,植株的光和速率显著下降[22]。可以推断,光合作用与植物防御之间这种负相关可能与植物能量分配有关,即植物机体合成的能量有一部分应用于防御反应的能量消耗[23],当植物遭受昆虫为害时,体内进行能量资源的再分配以防御昆虫的取食。植物对昆虫的取食响应是一个复杂的过程,通过植物和昆虫互作模型的建立,利用差异蛋白质组学技术能够发现一些昆虫取食应答蛋白,为解析昆虫取食条件下植物诱导防御机制,开拓害虫防治的新思路具有重要的指导意义。
[1]Agrawal A A,Karban R.Domatia mediate plant-arthropod mutualism[J].Nature,1997,387:562-563.
[2]Wang Y,Ge F,Li Z Y.Spatial-tempetialtrends of induced chemical change in pine Pinus massoniana[J].Acta Ecologica Sinica,2001,21(4):1256-1261.
[3]Talekar N S,Shelton A M.Biology,ecology and management of the diamondbackmoth[J].Ann.Rev.Entomo,1993,38:275-301.
[4]BarkerJE,PoppyGM,PayneCC.Suitabilityof Arabidopsis thaliana as a model for host plant-Plutella xylostella-Cotesia plutellae interactions[J].Entomologia Experimentaliset Applicata,2007,122,17-26.
[5]Liu L L,Zhang J,Zhang Y F,et al.Proteomic analysis of differentially expressed proteins of Arabidopsisthaliana response to specialist herbivore Plutella xylostella[J]. Chemical Research Chinese Universities,2010,26(6):958-963.
[6]Wang W,Scali M,Vignani R,et al.Protein Extraction for Two-dimensional Electrophoresis from Olive Leaf,a Plant Tissue Containing High Levels of Interfering Compounds[J].Electrophoresis,2003,24:2369-2375.
[7]BradfordM.ARapidandSensitiveMethodforthe Quantitation of Microgram Quantities of Protein Using the PrincipleofProteinDyeBinding[J].Analytical Biochemistry,1976,72:248-254.
[8]潘怡欧,刘琳琳,张炬红,等.茉莉酸诱导的拟南芥叶片蛋白质组分析[J].华北农学报,2010,25(2):35-39.
[9]Finnie C,Melchior S,RoepstorffP,et al.Proteome Analysis of Grain Filling and Seed Maturation in Barley[J].Plant Physiology,2002,129:1308-1319.
[10]Blum H,Beier H,Gross H J.Improved Silver Staining of Plant Proteins,RNA and DNA in Polyacrylamide Gels[J].Electrophoresis,1987,8:93-99.
[11]Hughes S M,Moroni-Rawson P,Jolly RD,et al.Submitochondrial distribution and delayed proteolysis of subunit C of the H+-transporting ATP-synthase in ovine ceroid-lipofuscinosis[J].Electrophoresis,2001,22:1785-1794.
[12]Zhang J H,Sun L W,Xi J H,et al.Proteomic Analysis of Interactions Between the Generalist Herbivore Spodoptera exigua(Lepidoptera:Noctuidae)and Arabidopsis thaliana[J].Plant Mol Biol Rep,2010,28:324-333.
[13]金晓芬,杨肖娥,冯英.蛋白质组学及其在植物营养学研究中的应用[J].植物生理学通讯,2005,41(6):705-709.
[14]Riccardi F,Gazeau P,De Vienne D.Protein changes in response to progressive water deficit in maize Quantitative variation and polypeptide identification[J].Plant Physiol,1998,117,1253-1263.
[15]Hanano S,Sugita M,Sugiura M.Isolation of a novel RNA-bindingproteinanditsassociationwithalarge ribonucleoprotein particle present in the nucleoplasm of tobacco cells[J].Plant Mol Biol,1996,31:57-68.
[16]Kim J S,Park S J,Kwak K J,et al.Cold shock domain proteins and glycine-rich RNA-binding proteins from Arabidopsis thaliana can promote the cold adaptation process in Escherichia coli.[J].Nucleic Acids Research,2007,35(2):506-516.
[17]潘妍,王玉成,张大伟,等.二色补血草LbGRP基因的克隆及抗逆能力分析[J].遗传,2010,32(3):278-286.
[18]Martin W,Brinkmarm H,Savonna C,et al.Evidence for a chimeric nature of nuclear genomes:eubacterial origin of eukaryotic glyceraldehydes-3-phosphate dehydrogenase genes[J].Proceedings of the National Academy of Sciences(PNAS),1993,90:8682-8696.
[19]王晓阳,孙立荣,李珊珊,等.拟南芥NAD激酶3(NADK3)参与调节镉胁迫响应[J].河南科技学院学报:自然科学版,2013,41(5):13-17.
[20]Hajirezaeii M R,Biemel T S,Peisker M,et al.The influence of cytosolic phosphorylating glyceraldehydes-3-phosphate dehydrogenase on potato tuber metabolism[J]. Journal of Experimental Botany,2006,57(10):2363-2377.
[21]叶文景.NaCl处理下红树植物白骨壤和木揽的蛋白质差异表达与质谱鉴定[D].厦门:厦门大学,2007.[22]Voelckel C,Baldwin I T.Herbivore-induced plant vaccination:array-studiesrevealthetransienceof herbivore-specific transcriptional imprints and a distinct imprint from stress combinations[J].Plant J,2004,38: 650-663.
[23]John Browse,Gregg A,Howe.New Weapons and a Rapid Response against Insect Attack[J].Plant Physiology,2008,146:832-838.
Analysis of Differential Proteins on Interaction between Arabidopsis thaliana and Plutella xylostella
Zhang Jing,Fan Juan
(Sericulture Institute of Heilongjiang Province,Harbin 150086)
The differential expressed proteins of Arabidopsis thaliana that fed by the special diets insect Plutella xylostella compared with the normal growth were analyzed by use of the two dimensional electrophoresis.The results of the intensities of 52 protein spots in more than 650 spots were detected on each 2D gel changed significantly by using Image MasterTM2D platinum 6.0 software. Among them,28 protein spots were up-regulated,and 24 spots were down-regulated under the insect feeding for 8 h.Some proteins were identified by MALDI-TOF-TOF MS successfully,such as the Glycine-rich RNA-binding protein,Glyceraldehyde-3-phosphate dehydrogenase,ATP synthase α and β subunits.These proteins might have played an important role in the process,and the further analysis study of the different proteins could offere the theoretical foundation to reveal the plant defense mechanism.
Plutella xylostella;Arabidopsis thaliana;2-DE;mass spectrometry;induced-defense
S433
A
1002-2090(2015)02-0001-04
10.3969/j.issn.1002-2090.2015.02.001
2014-05-26
张静(1984-),女,吉林大学毕业,现主要从事蜂业研究方面的工作。
