酸雨胁迫对毛竹叶片光合速率和叶绿素荧光参数的影响
2015-12-07王玉魁郭慧媛阎艳霞张汝民高岩温国胜
王玉魁,郭慧媛,阎艳霞,,张汝民,高岩,温国胜
1. 国家林业局竹子研究开发中心//浙江省竹子高效加工重点实验室,浙江 杭州 310012;2. 浙江农林大学//亚热带森林培育国家重点实验室培育基地,浙江 临安 311300;3. 国家林业局泡桐研究开发中心,河南 郑州 450003
酸雨胁迫对毛竹叶片光合速率和叶绿素荧光参数的影响
王玉魁1,郭慧媛1,阎艳霞1,3*,张汝民2,高岩2,温国胜2
1. 国家林业局竹子研究开发中心//浙江省竹子高效加工重点实验室,浙江 杭州 310012;2. 浙江农林大学//亚热带森林培育国家重点实验室培育基地,浙江 临安 311300;3. 国家林业局泡桐研究开发中心,河南 郑州 450003
为探讨酸雨胁迫下毛竹(Phyllostachys pubescens)色素含量变化及其对光合速率和叶绿素荧光动力学参数的影响,试验模拟酸雨(SO42-∶NO3-=4∶1摩尔比),设定pH 2.5、4.0、5.6和6.9(CK),对毛竹3年生盆栽实苗进行处理,测定了叶片色素含量、净光合速率(Pn)、叶绿素荧光动力学参数并进行分析。结果表明:pH 5.6酸雨处理增加了毛竹叶片中叶绿素a(Chl a)和类胡萝卜素(Car)含量,使得光合速率(Pn)升高;pH 4.0和pH 2.5处理使Chl a、Car、Chl a+b、Chl a/b降低,Pn下降;随着处理时间延长影响效果越明显,75 d时Pn与对照(CK)相比差异极显著。pH 4.0和pH 2.5处理使荧光诱导动力学曲线的IPJ点荧光强度出现不同程度降低,处理75 d时初始荧光(F0)降低11.73%和17.59%,峰值P点荧光(Fp)降低25.46%和31.41%,反应中心密度(RC/CSo)降低22.94%和26.22%,实际量子产额(Yield)下降20.41%和35.31%、光化学淬灭系数(qP)下降15.48%和19.42%,表观光合电子传递速率(ETR)下降22.02%和25.14%,非光化学淬灭系数(qN)上升23.16%和33.46%。pH 5.6处理使荧光强度升高,F0和Fp分别比CK增加30%和18.57%,qP和ETR上升8.38%和13.53%,qN下降17.65%。酸雨胁迫使毛竹叶片天线色素吸收能量(ABS/RC)、捕获能量(TRo/RC)和热耗散能量(DIo/RC)升高;以吸收光能为基础的性能指数(PIABS)、最大光化学效率(φPo)、电子传递量子产额(φEo)、电子传递效率(ψo)和电子传递能量(ETo/RC)降低。轻度酸雨处理促进毛竹叶片光合色素合成并提高 Pn,重度酸雨胁迫下光合色素含量降低,Pn下降。荧光诱导动力学曲线OJIP曲线及JIP-test分析可作为快速检测毛竹在酸雨胁迫条件下生理响应机制的方法。
酸雨胁迫;光合速率;叶绿素;荧光参数;毛竹
酸雨对植物的直接伤害主要发生在叶片上,一定强度的酸雨会导致植物叶片表层结构和膜系统损伤,引起植物细胞超微结构的变化(殷秀敏,2010),影响植物细胞对物质的选择性吸收(Back et al.,1992;李志国等,2007),使植物生理代谢出现紊乱(赵则海,2014)。酸雨胁迫会影响植物的叶绿素组成和含量(曹洪法等,1983;严重玲等,1995;张耀明等,1996;蒋馥蔚等,2009),干扰植物正常的光合生理功能(谢寅峰等,2008;Magel et al.,1990;鲁美娟等,2009),影响植物叶片气孔正常开放,导致光合速率的显著下降(邱栋梁等,2002)。但不同树种光合作用对酸雨胁迫的响应有所区别。齐泽民等(2006)研究发现,低浓度酸雨(pH 4.0~5.6)处理对杜仲(Eucommia ulmoides)最大净光合速率无显著影响。李万超等(2008)研究认为,一定程度的酸雨胁迫才能显著降低木荷(Schima superba)和青冈(Quercus glauca)的最大净光合速率。单运峰等(1988)认为pH 2.0酸雨处理使杉木(Cunninghamia lanceolata)和马尾松(Pinus massoniana)的净光合速率显著降低,而pH>2.0酸雨处理下两种植物的净光合速率均无显著变化。Lee et al.(1990)的试验结果是pH 3.0酸雨处理下,火炬松(Pinus taeda)幼苗的蒸腾速率和净光合速率均显著增大。金清等(2010)研究认为,酸雨处理前期,高浓度的酸雨胁迫对苦槠(Castanopsis sclerophylla)幼苗叶绿素含量、光合生理参数有显著影响,但随着酸雨处理时间的延长,对苦槠幼苗的影响逐渐减小。也有研究发现,酸雨中的 NO3-会引起植物叶片叶绿素含量上升(Shan,1998),在一定程度上提高净光合速率(Hogan,1998)。可见,酸雨对植物的影响除了酸度以外还取决于酸雨的硫氮比值,较低的硫氮比可能会减缓酸雨对植物的负面作用(Tamm et al.,1977)。
目前,毛竹(phyllostachys pubescens)在酸雨胁迫条件下光合色素含量变化及其对毛竹光合生理功能的影响研究鲜有报道。我国酸雨发生面积约150万 km2,集中分布在长江以南地区,这也是我国竹子资源主要分布区。毛竹是我国分布面积最广,社会、经济和生态效益较高的竹种,在我国竹产业中具有极其重要的地位(许改平等,2014)。本文综合分析了酸雨胁迫对毛竹叶片光合色素、净光合速率和叶绿素荧光参数的影响,旨在探究毛竹对酸雨胁迫的光合生理机制。
1 材料与方法
1.1 试验材料
试验苗木为3年生毛竹实生苗,苗高约1 m,于2013年4月移栽到高26 cm、直径35 cm的花盆中,每盆1株,红壤土(pH 6.0,有机质含量4.12 g·kg-1)栽培。盆栽苗置于温室中常规管理,缓苗期间用自来水浇灌。
1.2 试验方法
2013年6月选取长势相近的盆栽苗24盆,随机分为4组,每处理组6株重复,进行人工酸雨喷施。胁迫处理第15 d开始对毛竹叶片中光合色素指标进行测定,每15 d测定一次。模拟酸雨的配制根据浙江省临安市酸雨监测分析资料,用浓硫酸和浓硝酸配制酸雨母液,按照酸雨中SO42-∶NO3-=4∶1(摩尔比)的比例配制,每次喷施前用蒸馏水分别稀释成pH 2.5、4.0和5.6的酸液,对照(CK)用pH 6.9的自来水喷施(郭慧媛等,2014)。根据临安市常年月均降雨量确定酸雨喷施量,每周喷施 2次,每次每盆喷施100 mL。
1.3 测定指标和方法
1.3.1 色素含量测定
将新鲜的毛竹叶片洗净擦干,除去叶主脉剪成0.2 cm的碎片,称取0.2 g置于具塞试管中,加入10 mL 95%乙醇溶液,置于黑暗处室温下萃取48 h,每隔12 h摇晃一次,取上清液,使用UV-2550型紫外-可见分光光度计分别测定664、649和470 nm的光密度(optical density,OD)值;按Lichtenthaler(1987)方法计算Chl a、Chl b和Car含量。每个样品重复3次。1.3.2 净光合速率的测定
用LI-6400(LI-COR,USA)便携式光合仪及LI-6400-02B人工光源测定毛竹叶片的净光合速率。每次测定时自上向下选取第3片成熟叶片,测定时间在9:00─11:00时进行,每处理测试6株。在毛竹叶片净光合速率的测定过程中选取开放式气路,设置气流速度为0.5 L·min-1,叶温25 ℃,相对湿度60%,外界CO2浓度为400 μmol·mol-1,用以下公式计算酸雨胁迫下毛竹叶片净光合速率(Pn),横坐标为酸雨处理时间,纵坐标为净光合速率的值绘图。

1.3.3 叶绿素荧光参数测定
采用非调制式叶绿素荧光仪(PAM-2100,Germany)测定叶绿素荧光动力学参数。测定前将毛竹叶片暗适应15 min,用强度为3000 μmol·m-2·s-1的发光二极管光源,闪光照射1 s,以10 μs(2 ms之前)和1 ms(2 ms之后)的间隔记录叶绿素荧光信号,测得叶绿素荧光动力学曲线。每个处理6株,每株选取毛竹叶片5片,即每个处理重复30次,取平均值作为该处理叶片叶绿素荧光动力学曲线测定值。本文所用快速叶绿素荧光诱导动力学曲线(O-J-I-P)的JIP-test参数见表1。
2 结果与分析
2.1 酸雨胁迫对毛竹叶片色素含量的影响
酸雨胁迫下毛竹叶片中色素含量发生了变化(见表2)。酸雨胁迫75 d时,pH 5.6处理使Chl a、Chl b、Car、Chl(a+b)和 Chla/b分别比 CK增加21.86%、0%、26.67%、15.77%和24.25%,pH 4.0处理使之分别减少39.53%、19.28%、6.67%、33.89%和 27.24%,pH 2.5处理使之分别减少 53.49%、22.89%、13.33%、44.97%和 41.04%。从变化过程看,酸雨胁迫15 d,毛竹叶片中Chl a、Car、Chl(a+b)和Chla/b与CK比较均下降,Chl b在pH 4.0和pH 2.5处理下上升,在pH 5.6处理下下降。轻度酸雨(pH 5.6)处理使色素含量升高,可能是由于酸雨中NO3-的施氮效应在一定程度上促进了色素的合成(Shan,1998),重度酸雨(pH 4.0和pH 2.5)处理使色素含量下降,其原因可能是强酸雨进入植物体内,导致超氧阴离子自由基大量产生,诱发膜脂过氧化,使细胞膜功能受损,进而引发光合色素结构破坏所致(余春珠等,2005;李志国等,2007)。

表1 快速叶绿素荧光诱导动力学曲线(O-J-I-P)及JIP-test参数Table 1 The technical data of the O-J-I-P curves and the selected JIP-test parameters

表2 酸雨胁迫对毛竹叶片色素含量的影响Table 2 Effects of acid rainstress on leaves pigmentscontent of P. pubescens
2.2 酸雨胁迫对毛竹叶片净光合速率的影响
如图1所示,酸雨胁迫影响毛竹叶片的净光合速率(Pn)。在酸雨胁迫初期(15 d),各处理的净光合速率(Pn)与CK相比差异不显著(P>0.05),随着处理时间的延长,pH 5.6处理的毛竹叶片 Pn呈上升的趋势,在处理 75 d时与 CK相比增加20.6%,差异极显著(P<0.01);与之相反,随着酸雨胁迫时间的延长,pH 4.0和pH 2.5处理毛竹叶片Pn呈逐渐下降趋势;在处理45 d时,pH 2.5处理与CK的差异显著(P<0.05),处理60 d时,pH4.0与CK的差异显著(P<0.05),pH 2.5处理与CK差异极显著(P<0.01);在处理75 d时,pH 4.0和pH 2.5处理与CK差异极显著(P<0.01),分别比CK降低28%和31%。轻度酸雨处理促进了毛竹光合作用,而重度酸雨胁迫则使毛竹光合作用效率降低。
2.3 酸雨胁迫对毛竹叶片叶绿素荧光参数的影响
2.3.1 酸雨胁迫下毛竹叶片快速叶绿素荧光动力学曲线特征
植物发出的荧光强度随时间而变化,在从暗适应到暴露在光照条件下时,荧光强度先上升,然后下降。通常将刚暴露在光照下时的最低荧光定义为O点,荧光的最高峰定义为P点,快速叶绿素荧光诱导动力学曲线是指从O点到P点的荧光变化过程(李鹏民等,2005),其主要反映了光系统Ⅱ(PSⅡ)的原初光化学反应及光合机构的结构和状态的变化。图2是在不同酸雨胁迫下毛竹叶片快速叶绿素荧光动力学 OJIP曲线的变化情况。试验过程中毛竹叶片各处理在经过了暗适应进入饱和脉冲光照射后,叶绿素荧光迅速上升,在经过O、J、I和P各点后逐渐趋于平稳。初期各处理在 OIJP点增值缓慢,酸雨胁迫15 d时,各处理与CK的差异不显著(P>0.05);在胁迫30 d后,以pH 2.5处理在P点的荧光强度迅速减小,随后(45~60 d)pH 4.0和pH 2.5处理IPJ点的荧光强度均出现不同程度的降低,与CK相比差异显著(P<0.05),75 d时与CK差异极显著(P<0.01);pH 5.6处理60~75 d时IPJ点的荧光强度值则大幅度上升(P<0.01)。

图1 酸雨胁迫对毛竹叶片净光合速率的影响Fig. 1 Effects of acid rain stress on net photosynthetic rate in P. pubescens leaves
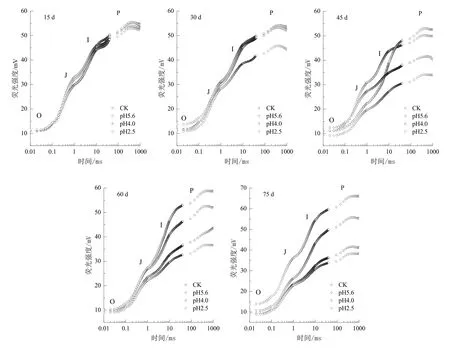
图2 酸雨胁迫对毛竹叶片快速叶绿素荧光诱导动力学曲线的影响Fig. 2 Effect of acid rain stress on the fast induction curves of chlorophyll fluorescence in P. pubescens leaves
JIP-测定(JIP-test)是以生物膜能量流动为基础建立的分析方法,利用该方法可以获得有关 PSⅡ的大量信息。酸雨胁迫下对毛竹叶片的 JIP-test分析见图3。酸雨胁迫15~30 d,毛竹叶片初始荧光(Fo)、P点荧光(Fp)、最大光化学效率(φPo)、电子传递量子产额(φEo)及电子传递效率(ψo)等参数变化均不显著(P>0.05)。随着胁迫时间延长,45 d时,pH 2.5处理使Fo、Fp、φPo、反应中心密度(RC/CSo)、天线色素吸收能量(ABS/RC)和捕获能量(TRo/RC)下降;pH 4.0处理使Fp、φPo、φEo、ψo、TRo/RC、电子传递能量(ETo/RC)和以吸收光能为基础的性能指数(PIABS)下降;pH 5.6处理则使Fo、Fp、φEo、ψo、TRo/RC、RC/CSo、PIABS上升。试验到75 d时,pH 4.0和pH 2.5处理使F0分别降低11.73%和17.59%,差异显著(P<0.05);使Fp降低25.46%和31.41%,RC/CSo降低22.94%和26.22%,PIABS降低49.14%和56.14%,差异极显著(P<0.01);ETo/RC降低 5.47%(P>0.05)和15.20%(P<0.05);使φPo、ψo和φEo不同程度降低(P>0.05);使ABS/RC上升14.55%和11.89%(P<0.05),用于热耗散的能量(Dio/RC)上升35.66%和34.19%(P<0.01)。以pH 5.6处理到75 d时,Fo和Dio/RC分别上升30.00%和26.05%,与对照相比差异极显著(P<0.01);Fp、RC/CSo、ABS/RC和 TRo/RC分别上升18.57%、13.18%、14.86%和12.20%,差异显著(P<0.05);PIABS上升41.54%(P<0.01),ψo和φEo分别上升11.83%和13.88%(P<0.05),φPo和ETo/RC上升但差异不显著(P>0.05)。

图3 酸雨胁迫下毛竹叶片JIP-test参数雷达图Fig. 3 The JIP-test parameters radar chart of P. pubescens leaves under acid rain stress
2.3.2 酸雨胁迫对毛竹叶片叶绿素荧光参数的影响
酸雨胁迫下毛竹叶片叶绿素荧光参数变化见图4。在酸雨处理15 d时,毛竹叶片光化学淬灭系数(qP)、非光化学淬灭系数(qN)、表观电子传递效率(ETR)和PSⅡ的实际量子产额(Yield)各参数变化不显著(P>0.05)。随着胁迫时间延长,pH 5.6处理的毛竹叶片qP、ETR和Yield逐渐上升,qP在45~75 d时比CK上升5.98%~8.38%,差异显著(P<0.05),ETR在75 d比CK上升13.53%(P<0.05),Yield上升(P>0.05),qN则在处理到75 d时比CK下降17.65(P<0.05);pH 4.0和pH 2.5处理使毛竹叶片qP、ETR和Yield随时间延长逐渐下降,pH 4.0处理在45~75 d时,使qP比CK下降8.27%~15.48%(P<0.05),ETR在60 d时下降17.23%(P<0.05),75 d时下降22.02%(P<0.01),Yield在75 d时下降20.41%(P<0.05);pH 2.5处理在45和60 d时使qP比CK分别下降12.34%和17.99%(P<0.05),在 75 d时 qP比 CK下降 19.42%(P<0.01);ETR在45 d时下降17.23%(P<0.05),在 60和 75 d时分别下降 23.59%和 25.14%(P<0.01);Yield在45和60 d时比CK分别下降16.69%和21.51%(P<0.05),在75 d时下降35.31%(P<0.01)。pH 4.0和pH 2.5处理使毛竹叶片qN上升,pH 4.0处理60 d和75 d时,qN分别上升22.83%和23.16%(P<0.05);pH 2.5处理45 d和60 d时,qN分别上升18.56%和29.92%(P<0.05),处理75 d时上升33.46%(P<0.01)。

图4 酸雨胁迫对毛竹叶片调制叶绿素荧光特征参数的影响Fig. 4 Effect of acid rain stress on the chlorophyll fluorescence characteristics of modulation parameters in P. pubescens leaves
3 讨论
植物叶片是捕获转换光能和制造有机物的最重要器官,植物光合能力可以用光合速率(Pn)、PSII实际光化学量子产量(Yield)等指标表示(Funk et al.,2013)。植物叶绿素含量变化直接影响其的光合作用过程(尹华军,2007),叶绿素含量与植物光合作用的捕捉光能力、过剩光能耗散、胁迫氧化产物失活等过程直接相关。本试验在酸雨胁迫初期(15 d),毛竹叶片中Chl a、Car含量和Chl a+b、Chl a/b均显著下降,此时,Pn与对照(CK)的差异不显著。随着胁迫时间延长,胁迫30~75 d时,以pH 5.6处理的毛竹叶片中Chl a、Chl b、Car含量和Chl a+b升高,Pn呈上升趋势;而pH 4.0和pH 2.5处理则使Chl a、Chl b、Car含量和Chl a+b及Chl a/b均下降,Pn呈逐渐下降趋势。试验表明:轻度酸雨(pH 5.6)处理促进了毛竹光合作用,而重度酸雨(pH 2.5和pH 4.0)胁迫则使毛竹光合作用效率降低。低硫氮比的酸雨胁迫中NO3-的施氮作用可以在一定程度上缓解轻度酸雨对毛竹的负面影响,但是当pH值过低时,氮肥效应无法补偿酸雨中由于H+取代卟啉环中的Mg2+而造成的叶绿素降解对毛竹的伤害,使毛竹叶绿素光合活性下降,并最终抑制毛竹光合作用。此外,逆境胁迫下植物Pn下降受气孔限制和非气孔限制两方面的影(Bethke et al.,1992;刘会超等,2004)。轻度胁迫下以气孔限制为主,严重胁迫下以非气孔限制为主(关义新等,1995)。重度酸雨胁迫诱导植物气孔部分关闭,引起植物体内外气体交换参数变化,导致毛竹叶片Pn下降;同时,酸雨破坏叶片膜系统,降低叶绿素光合活性,并最终影响到叶片的Pn(严重玲等,1995;邱栋梁等,2002)。关于毛竹在酸雨胁迫条件下对其叶片膜系统的影响机制和气体交换参数变化需进一步研究。
试验结果表明:重度酸雨胁迫,使毛竹叶片在IPJ点的荧光强度出现不同程度降低,处理75 d时初始荧光(F0)降低11.73%和17.59%,P点荧光(Fp)降低25.46%和31.41%,反应中心密度(RC/CSo)降低22.94%和26.22%,实际量子产额(Yield)下降20.41%和35.31%、光化学淬灭系数(qP)下降15.48%和19.42%,表观光合电子传递速率(ETR)下降22.02%和25.14%,非光化学淬灭系数(qN)上升23.16%和33.46%。轻度酸雨处理使荧光强度升高,F0和Fp分别比CK增加30%和18.57%,qP和ETR上升8.38%和13.53%,qN下降17.65%。qP是指被PSⅡ天线色素吸收,并用以推动线性光合电子流的能量部分,反映的是PSⅡ原初电子受体QA的氧化还原状态、PSⅡ开放中心的数目或开放程度(Demmigadams et al.,1996)、比例和参与CO2固定电子的多少(孙骏威等,2007;柏祥等,2011),即反映了由光合作用引起的荧光淬灭和光合活性的高低。qP数值越大,说明PSⅡ的电子传递活性越高,qP越小,则PSⅡ的开放程度越低(王可玢等,1997)。本试验轻度酸雨处理时,毛竹通过自身调节保持了PSⅡ的电子传递活性,使qP数值上升,但重度酸雨胁迫时电子传递受到严重影响。ETR反映了实际光强条件下的表观电子传递效率(张守仁,1999;胡启鹏,2008),可作为植物光合电子传递效率快慢的指标,且与植物净光合速率相关(李志博等,2005)。PSⅡ是植物光合机构对环境胁迫最敏感的部位,叶绿素荧光动力学参数的变化在一定程度上反映了外界环境对植物的影响(许大权,2002;Force et al.,2003;董瑜等,2013),可作为检验植物受胁迫伤害程度的指标(Krause et al.,1991;Jiang et al.,2003;马清等,2010),快速叶绿素荧光诱导动力学曲线(OJIP)能够提供关于PSⅡ的光化学信息(朱新广等,1999)。
PSⅡ的电子受体减少,会引起PSⅡ功能下降,表现为ETR的降低(宋维民等,2008;李晓锋等,2009;齐红岩等,2011;莫亿伟等,2011)。轻度酸雨处理使毛竹叶片电子传递效率提高,ETR值上升,但重度酸雨胁迫时电子传递受到严重抑制,导致光合电子传递速率下降和Pn下降。qN反映了植物 PSⅡ反应中心非辐射能量耗散能力的大小,是PSⅡ天线色素吸收的光能不能用于光合电子传递,而以热的形式耗散掉的光能部分(张其德等,2000),其值越大说明植物将捕捉到光能用于光合作用的部分越少,反映了植物耗散过剩光能为热的能力。热耗散是一种自我保护机制,对光合机构起一定的保护作用(王学英等,2005;陈建明等,2006)。毛竹受到重度酸雨胁迫时,qN大幅度上升,表明叶片受到严重胁迫,为防止光合机构受到破坏,使吸收的多余光能以热的形式耗散掉;而轻度酸雨处理,qN下降,表明尚未对毛竹光合系统造成破坏性伤害。轻度酸雨处理有利于毛竹幼苗的生长,表明毛竹幼苗可能更适于在微酸性环境中生长,但是随着酸度加强,其光合作用会受到抑制。酸雨胁迫使毛竹叶片天线色素吸收能量(ABS/RC)、捕获能量(TRo/RC)和热耗散能量(DIo/RC)升高;以吸收光能为基础的性能指数(PIABS)、最大光化学效率(φPo)、电子传递量子产额(φEo)、电子传递效率(ψo)和电子传递能量(ETo/RC)降低。这些参数的变化,反映了毛竹受酸雨胁迫时其“内在性”特征,可作为研究毛竹光合作用与环境关系的内在探针。
4 结论
酸雨胁迫对毛竹叶片光合色素含量和光合作用过程产生了不同程度影响,且影响程度随着胁迫时间的延长和酸度降低(pH值减小)而增强。酸雨胁迫对毛竹叶片中Chl a含量影响显著,而Car和Chl b含量变化对酸雨胁迫的响应不敏感。重度酸雨胁迫使毛竹叶片在IPJ点荧光强度出现不同程度降低,PSⅡ电子传递受到抑制,光合电子传递速率下降,导致毛竹光合作用效率降低;轻度酸雨处理则使毛竹叶片在IPJ点荧光强度出现不同程度升高,毛竹通过自身调节保持了 PSⅡ的电子传递活性,促进了毛竹叶片的光合作用。
BACK J, HUTTUNEN S. 1992. Effects of long-term exposure to simulated acid rain on conifer needle uhrastructure and hardening status [J]. Forest Ecology and Management, 51(1-3): 95-103.
BETHKE P C, MALCOLM C D. 1992. Stomatal and nonstomatalcomponents to inhibition of photosynthesis in leaves of capsicum annuum during progressive exposure to NaClsalinity [J]. Plant Physiology, 99(1): 219-226.
DEMMIGADAMS B, ADAMS W W, BARKER D H, et al. 1996. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation [J]. Physiologia Plantarum, 98(2): 253-264.
FORCE L, CRITCHLEY C, RENSEN J J. 2003. New fluorescence parameters for monitoring photosynthesis in plants [J]. Photosynthesis Research, 78(1): 17-33.
FUNK J L, CORNWELL W K. 2013. Leaf traits within communities: context may affect the mapping of traits to function [J]. Ecology, 94(9): 1893-1897.
HOGAN G D. 1998. Effect of simulated acid rain on physiology,growth and foliar nutrient concentrations of Sugar maple [J]. Chemosphere,36(4): 633-638.
JIANG C D, GAO H Y, ZOU Q. 2003. Changes of donor and acceptor side in photosystem2 complex induced by iron deficiency in attached soybean andmaize leaves [J]. Photosynthetic, 41(2): 267-271.
KRAUSE G H, WEIS E. 1991. Chlorophyll fluorescence and photosynthesis: the basics [J]. Annual Review of Plant BiologyAnd Plant Molecular Biology, 42: 313-349.
LEE W S, CHEVONE B I, SEILER J R. 1990. Growth and gas exchange of loblolly pine seedlings as influenced by drought and air pollutants [J]. Water, Air, and Soil Pollution, 51(1-2): 105-116.
LICHTENTHALER H K. 1987. Chlorophylls and carotenoids:Pigments of photosynthetic biomembranes [J]. Methods in Enzymology, 148(1): 350-382.
MAGEL E, HÖLLW, ZIEGLER H. 1990. Alteration of physiological parameters in needles of cloned spruce trees (Picea abies (L.) Karst.) by ozone and acid mist [J].Environmental Pollution, 64(3-4): 337-345.
SHAN Y. 1998. Effects of simulated acid rain onpinus densiflora:inhibition of net photosynthesis by the pheophytization of chlorophyll [J]. Water, Air, and Soil Pollution, 103(1-4): 121-127.
TAMM C O, WIKLANDER G, POPVI B. 1977. Effects of application of sulphuric acid to poor pine forests [J]. Water, Air, and Soil Pollution, 8(1): 75-87.
柏祥, 陈开宁, 黄蔚, 等. 2011. 黄菖蒲和美人蕉对水生梯度的响应差异[J]. 生态学杂志, 30(3): 464-470.
曹洪法, 高映新, 舒俭民. 1983. 模拟酸雨对农作物生长和产量影响的初步研究[J]. 植物生态学与植物学学报, 13(1): 58-65.
陈建明, 俞晓平, 程家安. 2006. 叶绿索荧光动力学及其在植物抗逆生理研究中的应用[J]. 浙江农业学报, 18(1): 51-55.
单运峰, 冯宗炜. 1988. 模拟酸雨对马尾松和杉木幼树的影响[J]. 环境科学学报, 8(3): 307-315.
董瑜, 田昆, 郭绪虎, 等. 2013. 不同区域气候条件影响下的纳帕海湿地植物叶绿素荧光特性[J]. 生态环境学报, 22(4): 588-594.
关义新, 戴俊英, 林燕. 1995. 水分胁迫下植物叶片光合的气孔和非气孔限制[J]. 植物生理学通讯, 31(4): 293-297.
郭慧媛, 马元丹, 王丹, 等. 2014. 模拟酸雨对毛竹叶片抗氧化酶活性及释放绿叶挥发物的影响[J]. 植物生态学报, 38(8): 896-903.
胡启鹏. 2008. 长白山岳桦叶性状和生长对海拔梯度响应研究[D]. 北京:北京林业大学: 14-15.
蒋馥蔚, 江洪, 李巍, 等. 2009. 不同起源时期的3种被子植物对酸雨胁迫响应的光合生理生态特征[J]. 植物生态学报, 33(1): 125-133.
金清, 江洪, 余树全, 等. 2010. 酸雨胁迫对苦槠幼苗气体交换与叶绿素荧光的影响[J]. 植物生态学报, 34(9): 1117-1124.
李鹏民, 高辉远, RETO J S. 2005. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J]. 植物生理学与分子生物学学报, 31(6): 559-566.
李万超, 江洪, 曾波, 等. 2008. 模拟酸雨对青冈和木荷幼苗光合响应特性的影响[J]. 西南大学学报: 自然科学版, 30(7): 98-103.
李晓锋, 侯瑞贤, 朱玉英, 等. 2009. 高温胁迫对大白菜叶绿素荧光特性的影响[J]. 农业工程学报, 25(1): 49-54.
李志博, 魏亦农, 张荣华, 等. 2005. 棉花不同叶位叶绿素荧光特性的初步研究[J]. 棉花学报, 17(3): 189-190.
李志国, 翁忙玲, 姜武, 等. 2007. 模拟酸雨对乐东拟单性木兰幼苗部分生理指标的影响[J]. 生态学杂志, 26(1): 31-34.
刘会超, 孙振元, 彭镇华. 2004. NaCl胁迫对五叶地锦生长及某些生理特性的影响[J]. 林业科学, 40(6): 63-67.
鲁美娟, 江洪, 余树全, 等. 2009. 模拟酸雨对山核桃和杨梅光合生理特征的影响[J]. 生态学杂志, 28(8): 1476-1481.
马清, 楼洁琼, 王锁民. 2010. Na+对渗透胁迫下霸王幼苗光合特性的影响[J]. 草业学报, 19(3): 198-203.
莫亿伟, 郭振飞, 谢江辉. 2011. 温度胁迫对柱花草叶绿素荧光参数和光合速率的影响[J]. 草业学报, 20(1): 96-101.
齐红岩, 华利静, 赵乐, 等. 2011. 夜间低温对不同基因型番茄叶绿素荧光参数的影响[J]. 华北农学报, 26(4): 222-227.
齐泽民, 钟章成. 2006. 模拟酸雨对杜仲光合生理及生长的影响[J]. 西南师范大学学报, 31(2): 151-156.
邱栋梁, 刘星辉, 郭素枝. 2002. 模拟酸雨对龙眼叶片气体交换和叶绿素a荧光参数的影响[J]. 植物生态学报, 26(4): 441-446.
宋维民, 周海燕, 贾荣亮, 等. 2008. 土壤逐渐干旱对4种荒漠植物光合作用和海藻糖含量的影响[J]. 中国沙漠, 28(3): 449-454.
孙骏威, 付贤树, 奚辉, 等. 2007. 水稻不同叶位气体交换和叶绿素荧光研究[J]. 浙江大学学报: 农业与生命科学版, 33(3): 277-283.
王可玢, 许春辉, 赵福洪, 等. 1997. 水分胁迫对小麦旗叶某些体内叶绿素a荧光参数的影响[J]. 生物物理学报, 13(2): 273-278.
王学英, 师生波, 吴兵. 2005. 西宁和海北麻花艽净光合速率和叶绿素荧光参数的日变化比较[J]. 西北植物学报, 25(12): 2514-2518.
谢寅峰, 杨万红, 陆美蓉, 等. 2008. 模拟酸雨胁迫下硅对髯毛箬竹光合特性的影响[J]. 应用生态学报, 19(6): l179-1l84.
许大权. 2002. 光合作用效率[M]. 上海: 上海科学技术出版社: 92-93.
许改平, 吴兴波, 刘芳, 等. 2014. 高温胁迫下毛竹叶片色素含量与反射光谱的相关性[J]. 林业科学, 50(5): 41-48.
严重玲, 李瑞智, 钟章成. 1995. 模拟酸雨对绿豆、玉米生理生态特性的影响[J]. 应用生态学报, (S1): 124-131.
严重玲, 钟章成, 李瑞智. 1995. 酸雨对农作物叶片活力、叶绿素及叶汁pH的影响[J]. 环保科技, 17(2): 27-30.
殷秀敏, 伊力塔, 余树全, 等. 2010. 酸雨胁迫对木荷叶片气体交换和叶绿素荧光参数的影响[J]. 生态环境学报, 19(7): 1556-1562
尹华军. 2007. 增温对川西亚高山针叶林不同光环境下几种幼苗生长的影响[D]. 成都: 中国科学院研究生院(成都生物研究所): 12-13.
余春珠, 温达志, 彭长连. 2005. 三种木本植物对酸雨的敏感性和抗性[J]. 生态环境学报, 14(1): 86-90.
张其德, 张建华, 刘合芹, 等. 2000. 限水灌溉和不同施肥方式对冬小麦旗叶某些光合功能的影响[J]. 植物营养与肥料学报, 6(1): 24-29.
张守仁. 1999. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报, 16(4): 444-448.
张耀明, 吴丽英, 王晓霞, 等. 1996. 酸雨对农作物的叶片伤害及生理特性的影响[J]. 农业环境保护, 15(5): 197-208.
赵则海. 2014. 模拟酸雨对五爪金龙幼苗光合生理特性的影响[J]. 生态环境学报, 23(9): 1498-1502.
朱新广, 张其德. 1999. NaCl对光合作用影响的研究进展[J]. 植物学通报, 16(4): 332-338.
Effect of Acid Rain Stress on Photosynthetic Rate and Chlorophyll Fluorescence Parameters in Leaves of Phyllostachys pubescens
WANG Yukui1, GUO Huiyuan1, YAN Yanxia1*, ZHANG Rumin2, GAO Yan2, WEN Guosheng2
1. China National Research Center of Bamboo, Key Laboratory of High Efficient Processing of Bamboo of Zhejiang Province, Hangzhou 310012, China; 2. The Nurturing Station for the State Key Laboratory of Subtropical Silviculture, Zhejiang Agriculture and Forestry University, Lin’an 311300, China; 3. Paulownia Research & Development Center of the State Forestry Administration, Zhengzhou 450003, China
In order to discuss the effect of acid rain stress on photosynthetic rate and chlorophyll fluorescence parameters in leaves of Phyllostachys pubescens, 3-year-old potted seedlings were tested for determination of leaves pigment content, net photosynthetic rate (Pn) and chlorophyll fluorescence dynamic parameters by experiment simulation of acid rain (SO42-∶NO3-=4∶1 mole ratio) with pH 2.5, 4.0, 5.6, and 6.9 (CK). Results showed that content Chl a, Car and Pnin leaves increased with the treatment of pH 5.6 acid rain, while Chl a, Car, Ch a+b, Ch a/b and Pndecreased with the treatment of pH 4.0 and pH 2.5. The effect got more apparent as time processing and it was highly significant compared with Pnand CK after 75 d treatment. IPJ fluorescence intensity reduced in diffenent degree with the treatment of pH 4.0 and pH 2.5. When handling 75d , F0reduced by 11.73% and 17.59%, Fpreduced by 25.46 and 31.41%, RC/CSo reduced by 22.94% and 26.22%, Yield reduced by 20.41% and 35.31%, qP reduced by 15.48% and 19.42%, ETR reduced by 22.02% and 25.14%,but qN increased 23.16% and 33.46%. pH 5.6 processing enabled fluorescence intensity rise, F0and Fpincreased by 30% and 18.57% than CK, qP and ETR increased by 8.38% and 13.53%, qN declined by 17.65%. The acid rain stress could rise ABS/RC, TRo/RC, TRo/RC and DIo/RC of bamboo leaf antenna pigment while drop PIABS, φPo, φEo, ψoand ETo/RC. Mild acid rain treatment promoted the synthesis of photosynthetic pigments and improved Pnwhile severe acid rain stress reduced the content of photosynthetic pigments and decreased Pn. OJIP and JIP-test analysis can be used as rapid methods to test the physiological response mechanism of bamboo under acid rain stress.
acid rain stress; photosynthetic rate; chlorophyll; fluorescence parameters; Phyllostachys pubescens
10.16258/j.cnki.1674-5906.2015.09.001
Q945.78;X171.5
A
1674-5906(2015)09-1425-09
王玉魁,郭慧媛,阎艳霞,张汝民,高岩,温国胜. 酸雨胁迫对毛竹叶片光合速率和叶绿素荧光参数的影响[J]. 生态环境学报, 2015, 24(9): 1425-1433.
WANG Yukui, GUO Huiyuan, YAN Yanxia, ZHANG Rumin, GAO Yan, WEN Guosheng. Effect of Acid Rain Stress on Photosynthetic Rate and Chlorophyll Fluorescence Parameters in Leaves of Phyllostachys pubescens [J]. Ecology and Environmental Sciences, 2015, 24(9): 1425-1433.
国家林业局948项目(2013-4-23);国家自然科学基金项目(30972397);浙江省科技计划项目(2012F20025)
王玉魁(1962年生),男,研究员,博士,研究方向为植物生理生态。E-mail: wyukui@126.com *通信作者。E-mail: yanyanxia202@gmail.com
2015-02-11
