宁夏盐池封育草地植被群落多样性及其与环境关系的典范对应分析
2015-12-07苗静张克斌刘小丹李海涛
苗静,张克斌,刘小丹,李海涛
1. 北京林业大学水土保持与荒漠化防治教育部重点实验室,北京 100083
宁夏盐池封育草地植被群落多样性及其与环境关系的典范对应分析
苗静1,张克斌1,刘小丹1,李海涛1
1. 北京林业大学水土保持与荒漠化防治教育部重点实验室,北京 100083
为探究干旱区封育草地植被群落多样性以及影响植被群落分布的主要环境因子,结合国家荒漠化定位监测项目,于2014年7月对宁夏盐池封育草地进行植被调查。将研究区分为核心区(E)、边缘区(E1)、外围区(E2),采用样线与样方结合法调查植物名称、植物种类、株数、高度、盖度、生物量、土壤容重、土壤含水量、土壤紧实度、地表生物结皮盖度。在植被调查的同时,对每个样方中的土壤进行取样,分别测定土壤速N、速P、速K、土壤有机质含量。采用Shannon-Wiener指数、Simpson指数、Pielou均匀度指数测得植被群落多样性,运用典范对应分析方法(CCA)分析植被与环境因子的相关关系。结果表明,边缘区共出现了22种植物,主要物种有:黑沙蒿(Artemisia ordosica)、刺沙蓬(Salsola ruthenica)、地锦草(Euphorbia humifusa)、赖草(Leymus secalinus)、蒙古虫实(Corispermum mongolicum)。其SW、SP指数高于其他2个区域,数值分别为2.548、0.893,植被多样性最为丰富,群落稳定性最高。外围区共出现了16种植物,主要物种有:苦豆子(Sophora alopecuroides)、阿尔泰狗娃花(Heteropappus altaicus)、蒙古虫实其JSW指数最高,为0.842,植被多样性较丰富,群落稳定性较高。核心区出现了19种植物,猪毛蒿(Artemisia scoparia)、刺沙蓬、苦豆子,其SW、SP、JSW指数均小于其它2个区域,数值分别为2.163、0.828、0.734,植被多样性较差,群落稳定性较低。植被与环境的典范对应分析表明,土壤容重和结皮盖度是影响干旱区封育草地植被群落的主要环境因子。
半干旱区;封育草地;植被多样性;典范对应分析
植物群落是植物与环境相互作用的产物,相互作用过程伴随物种多样性的变化(王世雄等,2010)。前人在对植被群落多样性及其与环境关系做了大量研究。张玲和王承义(2014)对大兴安岭过渡区阔叶红松林研究发现:原始阔叶红松林物种多样性指数最高,物种最为丰富,其次是天然次生阔叶林,再次是次生裸地;纬度、土壤含水率、土壤全氮含量是影响物种多样性的主要因子。许传阳等(2014)对宝天曼自然保护区研究表明:研究区植物物种多样性与土壤性状相关性最强,与地形地貌关系密切。郝占庆和郭水良(2003)通过典范对应分析得出影响长白山北坡草本植物主导因子是海拔。孔令伟等(2014)对华北落叶松人工林研究发现:在海拔1400~2000 m的范围,随海拔升高,Shannon-Wiener多样性指数和Pielou均匀性指数变化较为平缓,而Simpson多样性指数和Menhinick丰富度指数则表现出上下波动的变化趋势,坡位对草本植物多样性影响程度较大。苏日古嘎等(2010)对松山自然保护区研究发现森林群落的决定性环境因子是海拔。
本文研究宁夏盐池封育草地植被群落多样性以及探究影响植被群落分布的主要环境因子,采用样线与样方结合的调查方法,通过典范对应分析探究影响草地植被群落的主要环境因子,为研究草场植被的变化规律、恢复和重建退化的草场提供科学依据和理论基础。
1 研究方法
1.1 研究区概况
盐池县位于宁夏回族自治区东部,北纬37°04′~38°10′,东经106°30′~107°41′。盐池县地貌以干旱剥蚀或风蚀为主,海拔在1290~1950 m之间。土壤以灰钙土为主,其次是黑垆土和风沙土,另有少量黄土、盐土、白浆土等。盐池县北与毛乌素沙地相连,南靠黄土高原,自南向北地形上是从黄土高原向鄂尔多斯台地(沙地)过渡地带,在气候上是从半干旱区向干旱区的过渡地带,在植被上是从干草原向荒漠的过渡地带,在资源利用上是从农区向牧区过渡地带,这种地理上的过渡性造成了盐池县生态环境的多样性和脆弱性。
1.2 外业调查
结合国家荒漠化定位监测项目,选择具有代表性的地段采用GPS定位设置固定样地。本实验地位于盐池县柳杨堡乡人工封育区。草原植被是封育区主要植被类型,主要包括以下群从:(Ⅰ)黑沙蒿+刺沙蓬群从:群落高度一般大于30 cm,盖度大于40%,群落有分层现象,第一层为黑沙蒿(Artemisia ordosica),第二层为刺沙蓬(Salsola ruthenica);(Ⅱ)猪毛蒿+刺沙蓬群从:群落高度较低,20 cm左右,但盖度较高,达60%。该群落分层不明显。调查时将样地分为核心区(E)、外围区(E1)以及边缘区(E2)。核心区是采用铁丝网围栏,完全排除人为放牧和其他干扰,是 1991年建立的第一批全国防沙治沙试验示范区,封育时间超过20年。边缘区从 2002年以来采取完全封育措施,封育时间10余年。外围区从2002年以来也采用了封育,但仍然受到一些人为影响,时有放牧活动。调查时间为2014年7月。调查方法为从封育区沿着样线方向随机布设1 m×1 m的样方,核心区、外围区和边缘区各布设10样方,共计30个样方。每个样方调查植物名称、植物种类、株数、高度、盖度、生物量、土壤容重、土壤含水量、土壤紧实度、地表生物结皮盖度,在植被调查的同时,对每个样方中的土壤进行取样,土壤样品室内采用国家标准方法分别测定土壤速N、速P、速K、土壤有机质含量(中国科学院南京土壤研究所,1978)。
1.3 数据处理
通过 EXCEL2010建立研究区草地植被数据库、环境因子数据库,计算植物物种重要值矩阵,并计算植物物种多样性指数。使用R语言对环境数据矩阵进行标准化处理,采用Canoco for windows 4.5进行典范对应分析。
1.3.1 重要值
重要值计算公式(郑翠玲等,2005)如下:


1.3.2 多样性指数
植被群落多样性指数采用 Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数进行分析,其公式(李瑞等,2009)如下:
Shannon-Wiener多样性指数:

式中:Ni其中为样方中第i种植物的重要值;N为样方植物重要值总和。
Simpson优势度指数:

Pielou均匀度指数:

式中:H为Shannon-Wiener多样性指数;S为植物物种数。
2 结果与分析
2.1 柳杨堡人工封育区植被群落多样性
柳杨堡不同封育区域的SW指数、SP指数、JSW均匀度指数见表1。
经调查,核心区(E)出现了19种物种,主要物种有:猪毛蒿(Artemisia scoparia)、刺沙蓬、苦豆子(Sophora alopecuroides)。边缘区(E1)出现了 22种植物,主要物种有:黑沙蒿、刺沙蓬、地锦草(Euphorbia humifusa)、赖草(Leymus secalinus)、蒙古虫实(Corispermum mongolicum)。外围区(E2)出现了16种植物,主要物种有:苦豆子、阿尔泰狗娃花(Heteropappus altaicus)、蒙古虫实。

表1 不同封育区域植被多样性指数Table 1 Species diversity in different fencing areas
Shannon-Wiener指数综合了物种群落的均匀度和丰富度,是表述物种多样性最好的指数。由表 1可知,封育区域 Shannon-Wiener指数的排列顺序为:边缘区>外围区>核心区。边缘区 Shannon-Wiener指数是2.548,说明边缘区物种多样性最高,群落组成复杂,群落稳定性最高。核心区的Shannon-Wiener指数为2.163,Shannon-Wiener指数最低,说明核心区群落组成简单,物种多样性较低。
生态优势度是群落水平上的综合数值,它是把群落作为一个整体而把各个种的重要性总结为一个合适的度量值,以表征群落的组成结构特征。由表1可知,不同封育区域Simpson指数的变化为:边缘区>外围区>核心区。变化趋势与 Shannon-Wiener指数一致。边缘区的优势度最高,表明该区域优势种明显。该区域黑沙蒿具有明显优势。
群落均匀度反映群落种群空间分布的均匀程度。群落均匀度与群落优势度是2个相反的概念,该指标常作为群落物种多样性指数的辅助指标,在反映群落特征结构方面有重要意义。核心区、边缘区、外围区的Pielou均匀度指数分别为0.734、0.824、 0.842,排列顺序为外围区>边缘区>核心区。说明外围区的群落种群分布最均匀,核心区群落种群分布最集中。
综上所述,核心区3个多样性指数均为最低值,说明核心区群落结构单一、不稳定,生物多样性低。外围区物种分布均匀,结构稳定,群落物种多样性高。边缘区是3个封育区域中多样性指数最高的区域,其物种多样性最高,群落结构复杂且稳定。
2.2 柳杨堡人工封育区植被典范对应分析
为了探究影响柳杨堡人工封育区植被的主要环境因子,本文对封育区域的植被和环境进行典范对应分析。分析结果见图1和表3。
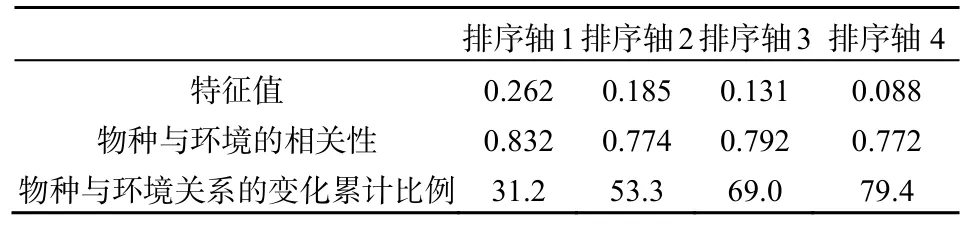
表2 CCA排序图特征值及物种与环境的相关性Table 2 Eigenvalues and correlations between special axes and environmental axes of CCA ordination diagram
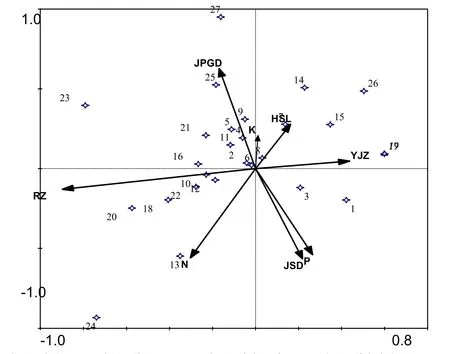
图1 柳杨堡人工封育区植被与环境因子的CCA排序图Fig. 1 CCA ordination diagram of plants and environmental factors in artificial fencing area in Liu Yang Bao
CCA排序图中箭头表示环境因子,箭头连线的长短表示群落的分布于环境因子相关性的大小,箭头所处的象限表示环境因子与排序轴之间相关性的正负(熊先华等,2014)。由表2可知,物种CCA排序轴与环境因子相关。在前2个排序轴中,物种与环境的相关性分别为 0.832、0.774,物种与环境关系的变化累计比例分别为31.2%、53.3%。研究表明,如果前3个排序轴的物种与环境关系方差累计贡献率大于 40%,则排序结果可行有效(Gauch,1982)。故本文排序结果能较好的解释物种与环境之间的相关关系。
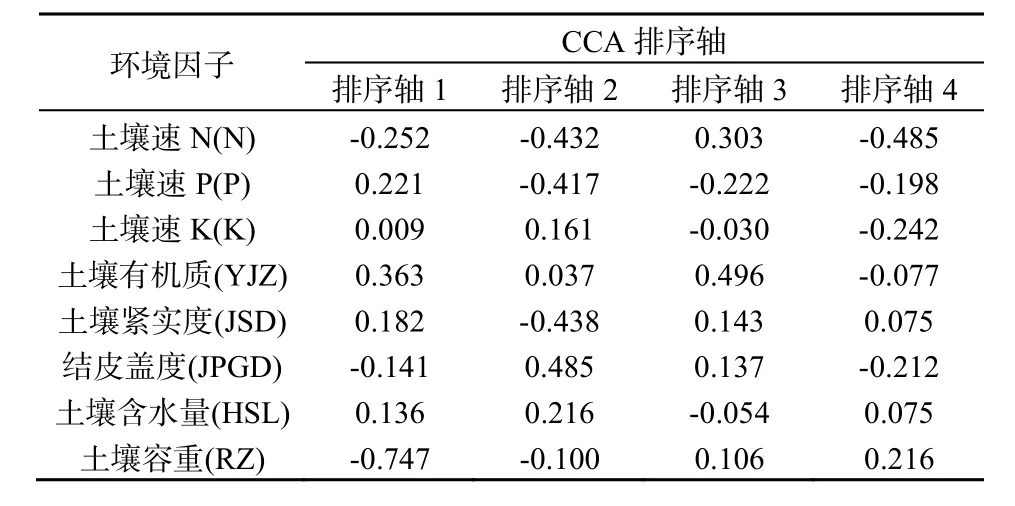
表3 环境因子与CCA排序轴的相关系数Table 3 The correlation coefficient of environmental factors andCCA axes

表4 CCA分析植物Table 4 Lists of Plants in the CCA diagram
图1可知,选取的8个环境因子对植被都有不同程度的影响。在第一排序轴中,环境因子的影响程度分别是:土壤容重>土壤有机质>土壤速 N>土壤速 P>土壤紧实度>结皮盖度>土壤含水量>土壤速K。第一排序轴与土壤容重呈极显著负相关,相关系数是-0.7472(表 3);与土壤有机质呈显著正相关,相关系数为0.3633。这也说明沿着第一排序轴从左到右,土壤容重越来越小,土壤有机质含量越来越高。在第二排序轴中,物种与环境的影响程度顺序为:结皮盖度>土壤紧实度>土壤速 N>土壤速 P>土壤含水量>土壤速 K>土壤容重>土壤有机质。第二排序轴与结皮盖度呈极显著正相关,相关系数为0.4853;与土壤紧实度、土壤速N呈显著负相关,相关系数分别为-0.4379、-0.4321。说明沿着第二排序轴从下至上,结皮盖度越来越大,土壤紧实度和土壤速N含量逐渐降低。
根据以上分析,可以得出:沿第一排序轴CCA能清晰区分柳杨堡人工封育区物种与各个环境因子之间的相关关系。其次,影响柳杨堡人工封育区的主要环境因子是土壤容重和结皮盖度。
3 结论与讨论
3.1 结论
柳杨堡人工封育区由于所处的地理位置特殊,其生态环境较为脆弱。不同封育年限的区域物种多样性存在差异。边缘区物种多样性最为丰富,外围区其次,核心区最差。边缘区群落优势度指数最高,其次是外围区,最差是核心区。而外围区物种分布最为均匀,边缘区物种分布较为均匀,核心区物种分布较集中。通过典范对应分析分析植被分布的主要环境因子,发现土壤速N、土壤速P、土壤速K、土壤有机质、土壤紧实度、结皮盖度、土壤含水量、土壤容重对柳杨堡人工封育区植被群落存在不同程度的影响。在第一排序轴中,物种与土壤容重和土壤有机质含量之间相关性显著(P=0.05)。在第二排序轴中,物种与结皮盖度、土壤紧实度、土壤速N含量之间相关性显著(P=0.05)。由此得出,土壤容重和结皮盖度是影响柳杨堡人工封育区植被群落的主要环境因子。
3.2 讨论
自然界的植被群落是植物与环境相互作用的产物,其空间分布特征受环境因子、生态、非生态因子,生境梯度等多种因素的影响。容重作为土壤的一项物理性质,影响着土壤水肥气热条件的协调(胡伟,2013)。容重大的土壤,其物理结构紧实、土内透气少、土块大而板结;加之土壤水分少,水分扩散快,往往导致土壤活力不高、养分易流失,进而限制植物根系下扎,使其难从土中摄取养分,植物生长受到抑制甚至死亡。不同封育区域的容重不同,植物生长、发育不同,进而群落结构特征、植物物种多样性存在差异。生物结皮是由藻类(包括蓝藻)、地衣、苔藓、真菌和细菌等与其下层很薄的土壤共同形成的一个复合的生物土壤层。土壤结皮通过改变土壤结构、土壤稳定性、有机质含量、土壤粗糙度等进而影响土壤上层的水分循环(吴玉环,2002)。在宁夏盐池县,长期封育形成的结皮改变了土壤水分的传导,有限的降水在沙层土壤中的时空分布发生了改变,降水后水分的渗透率大大降低。本研究中核心区由于长时间的封育形成了高覆盖度的土壤结皮,影响水分入渗,降低了植被利用的有效性,植物生长受到影响,群落多样性降低。相比之下,边缘区还未完全形成结皮,围栏封育也避免人畜破坏,植被生长良好,植被盖度、生物量等大大增加,群落多样性增加。同时,封育区供植物生长的资源有限,随封育时间的延长生物量不会无限制的增加(毛绍娟等,2015)。所以针对本地,建议以2~3年为周期进行轮牧(郝智如和张克斌,2014)。
由于实验条件的限制,本研究没有对影响人工封育区的气象因子、人为因子等进行研究讨论,分析结果具有一定的局限性,在以后的工作中需要加强这方面的研究。
GAUCH H G. 1982. Multivariate Analysis in Community Ecology [M]. Cambridge: Cambridge University Press.
郝占庆, 郭水良. 2003. 长白山北坡草本植物分布与环境关系的典范对应分析[J]. 生态学报, 23(10): 2003-2007.
郝智如, 张克斌. 2014.宁夏盐池人工封育区植物群落结构及多样性[J].生态环境学报, 23(3): 377-384.
胡伟. 2013. 土壤容重对烟草生长及植烟土化学性质影响的研究[D]. 云南: 昆明理工大学: 13-17.
孔令伟, 陈祥伟, 鲁绍伟, 等. 2014. 华北落叶松林木生长草本植物多样性及地形因子之间的关系[J]. 水土保持通报, 34(5): 60-66.
李瑞, 张克斌, 边振, 等. 2009. 半干旱地区湿地生态系统植物α及β多样性分析[J]. 干旱区资源与环境, 23(9): 139-145.
毛绍娟, 吴启华, 祝景彬, 等. 2015. 藏北高寒草原群落维持性能对封育年限的响应[J]. 草业科学, 24(1): 21-30.
苏日古嘎, 张金屯, 张斌, 等. 2010. 松山自然保护区森林群落的数量分类和排序[J]. 生态学报, 30(10): 2621-2629.
王世雄, 王孝安, 李国庆, 等. 2010. 陕西子午岭植物群落演替过程中物种多样性变化与环境解释[J]. 生态学报, 30(6): 1638-1647.
吴玉环. 2002. 生物土壤结皮的生态功能[J]. 生态学杂志, 21(4): 41-45.
熊先华, 高末, 吴庆玲, 等, 2014. 江西新岗山亚热带森林生物多样性监测样地 A草本植物分布与环境关系的典范对应分析[J]. 生态科学, 33(3): 474-479.
许传阳, 陈志超, 郝成元. 2014. 宝天曼自然保护区植物物种多样性与其主要生境因子相关性[J]. 草业科学, 31(9): 1623-1627.
张玲, 王承义. 2014. 大小兴安岭过渡区阔叶红松林次生演替阶段群落多样性指数与环境因子关系[J]. 森林工程, 30(5): 1-5.
郑翠玲, 曹子龙, 赵廷宁, 等. 2005. 浑善达克沙地南缘农牧交错带弃耕地植被的演替规律[J]. 中国水土保持科学, 3(1): 72-76.
中国科学院南京土壤研究所. 1978. 土壤理化分析[M]. 上海: 上海科学技术出版社: 62-141.
Diversity of Plant Community in Relation to Environment Factors Based on CCA in Fencing Region in Yanchi, Ningxia
MIAO Jing1, ZHANG Kebin1, LIU Xiaodan1, LI Haitao1
1. College of Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China
To explore the diversity of plant community and the main environmental factors affecting the distribution in fencing region in arid area, combined with the national desertification monitoring and positioning project, vegetation investigation was carried out in July, 2014 in Yanchi, Ningxia. The studied area was divided into three areas:core area (E), edge area (E1) and peripheral area (E2). Samples of the plant population were recorded according to name, species, number, height, coverage, height, biomass, soil bulk density, soil moisture, soil compaction, the coverage of biological crust. Meantime, soil available N, fast P, fast K, the content of soil organic matter were respectively measured. They were analyzed in respect to diversity indices for community diversity (species diversity index, community dominant concentration index, community evenness index), and canonical correspondence analysis in order to analysis of the relationship between vegetation and environmental factors. The results showed 22 kinds of plants were found in edge area, the main species include: Artemisia ordosica, Salsola ruthenica, Euphorbia humifusa, Leymus secalinus, Corispermum mongolicum and SW (2.548), SP (0.893) index is higher than the other two areas. Its vegetation diversity is richest and the community stability is the highest. There are 16 kinds of plants were appeared in peripheral area, the main species include: Sophora alopecuroides, Heteropappus altaicus, Corispermum mongolicum and JSWindex (0.842) is highest which means vegetation diversity rich and community high. There are 19 kinds of plants were found in core area, the main species include: Artemisia scoparia, Salsola ruthenica, Sophora alopecuroides. SW (2.163), SP (0.828), JSW(0.734) index is the lowest than the other two areas, its vegetation diversity is poor and community stability is low. Soil bulk density and soil crust coverage are the main environmental factors affecting plant community in fencing grassland in arid area based on CCA.
arid area; fencing grassland; vegetation diversity; canonical correspondence analysis
10.16258/j.cnki.1674-5906.2015.05.006
Q948;X176
A
1674-5906(2015)05-0762-05
苗静,张克斌,刘小丹,李海涛. 宁夏盐池封育草地植被群落多样性及其与环境关系的典范对应分析[J]. 生态环境学报, 2015, 24(5): 762-766.
MIAO Jing, ZHANG Kebin, LIU Xiaodan, LI Haitao. Diversity of Plant Community in Relation to Environment Factors Based on CCA in Fencing Region in Yanchi, Ningxia [J]. Ecology and Environmental Sciences, 2015, 24(5): 762-766.
国家自然科学基金项目(30771764):国家林业局宁夏盐池荒漠化定位监测项目
苗静(1991年生),女,硕士研究生。主要研究方向为荒漠化防治与监测。Email:miaojing824@163.com *通信作者:张克斌(1957年生),男,教授,博士生导师。主要研究方向为荒漠化防治与监测及干旱区环境管理。Email:ctccd@126.com
2015-02-02
