宁夏河东沙区柠条植株叶片蒸腾对干旱胁迫的响应
2015-12-07韩磊孙兆军展秀丽焦炳忠董良
韩磊,孙兆军,展秀丽,焦炳忠,董良
1. 宁夏大学新技术应用研究开发中心,宁夏 银川 750021;2. 宁夏大学资源环境学院,宁夏 银川 750021;3. 宁夏大学土木与水利工程学院,宁夏 银川 750021
宁夏河东沙区柠条植株叶片蒸腾对干旱胁迫的响应
韩磊1,孙兆军1,展秀丽2,焦炳忠3,董良1
1. 宁夏大学新技术应用研究开发中心,宁夏 银川 750021;2. 宁夏大学资源环境学院,宁夏 银川 750021;3. 宁夏大学土木与水利工程学院,宁夏 银川 750021
以宁夏河东沙地沙生灌木柠条(Caragana korshinskii Kom.)为材料,采用采用盆栽方法分析不同土壤水分条件下的柠条植株叶片蒸腾日变化特征及其与环境因子的相关关系,研究干旱胁迫下柠条蒸腾对主要环境驱动因子的响应机制,为区域植被恢复与重建及林分结构的优化配置提供理论依据。结果表明,(1)充分供水条件下的柠条蒸腾速率是土壤水分严重胁迫下的3.43倍,随着水分胁迫的加剧,蒸腾速率峰值前移,且日蒸腾过程在较为干旱时表现为双峰型;各水分梯度下柠条蒸腾速率日过程与气孔导度具有相同的变化规律,呈极显著的线性相关。(2)随着土壤含水量的逐渐降低,柠条苗木叶片蒸腾速率先增加后缓慢下降,过高的土壤含水量同样会抑制植物的蒸腾,该临界值为 15.78%,即土壤质量含水量为田间持水量的74.1%。(3)低光强柠条叶片蒸腾作用的光响应曲线差别不大,都表现出随着光合有效辐射强度增加而缓慢上升的趋势,但启动速度随着土壤含水量的减少而降低;在土壤水分受到限制时(土壤质量含水量≤40%田间持水量),进一步增强光合有效辐射量(≥1 000 μmol·m-2·s-1)并没有使蒸腾速率提高,严重水分胁迫下,增加光辐射强度反而导致蒸腾速率降低。(4)柠条蒸腾速率的主要影响因子偏相关分析表明:充分供水下,光合有效辐射(PAR)是主要影响因子,贡献率为54.4%;随着土壤含水量的下降,气孔对PAR的敏感性下降,叶片内外水汽压饱和亏缺成为调节柠条蒸腾过程的主导因素。
柠条;蒸腾;水分胁迫;河东沙地
蒸腾是植物耗水特征的主要方式,在“土壤-植物-大气连续体”水热传输过程中占有极为重要的地位,蒸腾效率是一个涉及气孔导度、叶片水势和水力导度与环境因子相互作用的集合特征,有关调控森林蒸腾的多因子交互作用的研究已有报道(Wallace和McJannet,2010;Fernández等,2009;奚如春等,2006),但未取得一致结论。气孔导度决定植物的蒸腾效率,它对驱动水汽移动的水汽应力的响应受外界环境因子的影响,包括物理学机理和叶片生物学特性的过程,蒸腾强度主要取决于土壤的可利用水、液态水转化为水蒸汽所必需的能量以及叶片内部与外界之间的水汽压梯度(赵平,2011)。
不同地区不同树种的蒸腾耗水规律存在着明显的差异,当土壤水分供应明显下降时,植物蒸腾对环境因子响应的敏感性差异较大(常学向和赵文智,2009;Han等,2012),在极端干旱的沙漠地区,植物的蒸腾速率随着叶片气孔导度的变化呈现出周期性的波动(Whitley等,2009;司建华等,2008),对于降水量稀少且无效蒸发大的宁夏河东沙地,人工林的营造改变了沙地水分的再分配。柠条(Caragana korshinskii Kom.)作为治理水土流失和退化沙化草场的先锋植物,其对干旱的适应机制、光合和呼吸特征等研究已有不少积累(徐当会等,2012;余峰等,2011;李清河等,2006),但土壤水分胁迫严重时,苗木叶片气孔水汽交换及其对土壤水分利用策略可能会有差异,这种差异与哪些环境因子有关,这些问题有待于进一步研究。因此,研究柠条蒸腾速率在不同土壤水分条件下对环境主要驱动因子响应的敏感性,对分析干旱胁迫下环境因子对林木水分利用的影响机理,在全球变暖加速地球系统水循环的背景下显得尤为重要。
1 材料和方法
1.1 研究区概况
试验地点位于宁夏河东沙区南缘的同心县,地理坐标为北纬36°52′06″,东经105°59′7″,海拔1568 m,在气候区上属于温带大陆性气候,干旱少雨、风大沙多。年平均气温 8.7 ℃,极端最高气温38.5 ℃,极端最低气温为-27.3 ℃,≥10 ℃年平均积温为2963.1 ℃,无霜期165~183 d;多年平均降雨量251 mm,降水多集中在7─9月份,占年降水量的72.4%。多年平均蒸发量为2340 mm,年日照时数多年平均为2900~3055 h。试验区土壤类型为灰钙土区的沙化土壤,田间最大持水量21.3%,表层土壤平均容重1.47 g·cm-3。
1.2 研究方法
在试验区选取3年生柠条植株,于2014年4月植入试验花盆(高33 cm×上口径35 cm×下口径29 cm),试验盆土壤采自试验区大田,栽植后放置于试验地的温室内并充分补水,使之成活并正常生长。7月份人工控制土壤水分含量(SWC)分别为田间最大持水量(w)的80%~100%(T1:充分供水),60%~80%(T2:轻度水分胁迫),40%~60%(T3:中度水分胁迫)和20%~40%(T4:重度水分胁迫),即土壤含水量为 T1:17.0%~21.3%,T2:12.8%~17.0%,T3:8.5%~12.8%,T4:4.3%~8.5%,每处理3个重复,待长势稳定后,于2014年8月开始观测不同水分梯度下苗木蒸腾速率变化节律及其影响因子。
1.2.1 植物叶片蒸腾速率
采用Li-6400(Li-Cor Inc., Lincoln, USA)观测,选择苗木中上部向阳面生长良好的叶片,在生长季典型晴天,从7:00至19:00每隔2 h测定一次,每个叶片每次连续采取6个稳定的数据,取平均值。测定内容有蒸腾速率(Tr)、气孔导度(Gc)、叶温(Tl)等。
1.2.2 蒸腾-光响应曲线
利用 Li-6400-02B红蓝光源(Li-Cor Inc., Lincoln, USA)设定模拟光辐射强度,模拟光辐射强度的梯度设置为:0、50、100、200、400、600、800、1000、1200、1500、1800、2000和2200 μmol·m-2·s-1,观测苗木蒸腾速率对光辐射强度的响应。
1.2.3 蒸腾速率对土壤水分胁迫的响应
7月份选择长势良好的盆栽苗木,进行萎蔫试验。在干旱处理前充分浇水,使土壤水分达到田间持水量水平,从第6天开始不再浇水,使其自然干燥。干旱过程中每隔3 d对土壤含水量和叶片蒸腾速率进行观测,同时用 ML3X土壤水分速测仪(Delta-T Inc., UK)测定盆栽土壤含水量(体积),每次测定3个重复,直至苗木最后萎蔫枯死。土壤水分速测仪测定前用烘干法标定,搭建透光防雨棚减小降水对试验的影响。
1.2.4 气象因子
采用美国Vantage pro 2自动气象站(Davis Inc., USA),每10 min记录一次气温、相对湿度(2 m处)、光量子通量密度、降雨量等气象因子。
1.3 数据分析
采用SPSS 19.0统计分析软件(SPSS Inc, USA)对数据进行相关分析、单因素方差分析、多变量偏相关分析。
2 结果与分析
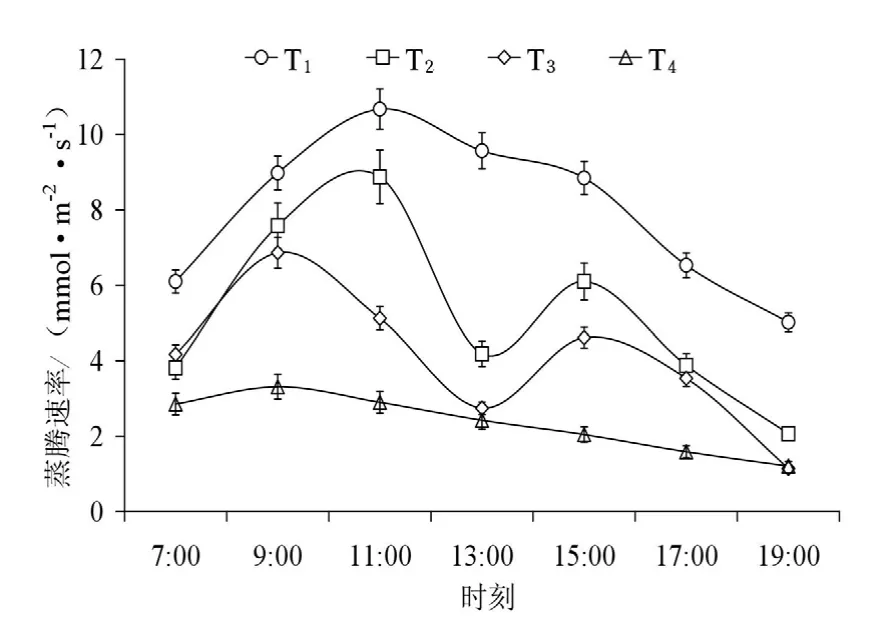
图1 不同水分条件下蒸腾速率日变化Fig. 1 Diurnal changes of leaf transpiration of Caragana korshinskii Kom. in different soil water content
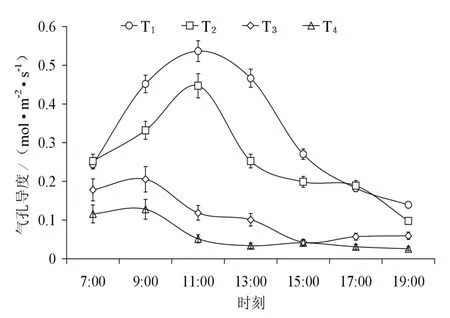
图2 不同水分条件下叶片气孔导度日变化Fig. 2 Diurnal changes of leaf stomatal conductance of Caragana korshinskii Kom. in different soil water content
2.1 不同土壤水分下柠条叶片蒸腾速率日变化节律
图1和图2为不同土壤水分条件下柠条叶片蒸腾速率和气孔导度变化的日过程,可看出各水分梯度下蒸腾速率日过程与气孔导度具有相同的变化规律,但峰值出现的时间存在差异。日出后,随着光照增强,气温升高,植物气孔开放,蒸腾作用逐渐增强;中午由于光照强度大、气温高、空气湿度低,植物为减少过度蒸腾失水,叶片气孔开度会减小或关闭,蒸腾强度随之大幅下降;蒸腾的减弱使叶细胞的含水量在午后(15:00)得到一定程度的恢复(T2,T3),蒸腾有所回升,此时柠条叶片蒸腾速率变化呈现双峰型,最后随着光照强度的减弱和日落前后气孔的逐步关闭,蒸腾作用逐渐下降。高土壤含水量下苗木具有较高的蒸腾速率,日均值分别为(T1:(7.959±2.080) mmol·m-2·s-1;T2:(5.214±2.398) mmol·m-2·s-1);在低水平的土壤含水量时,苗木叶片蒸腾速率的日变化相差不大,都维持在较低的水平,日均值分别为(T3:(4.027±1.817) mmol·m-2·s-1;T4:(2.332±0.761) mmol·m-2·s-1),充分供水条件下的柠条苗木蒸腾速率是土壤水分严重胁迫下的3.43倍,这是因为当根向叶片供水能力小于因各项生理活动的耗水速率时,柠条叶片气孔开度降低或部分关闭以防止水分向外散失;且随着水分胁迫的加剧(T3,T4),叶部水分亏缺提早出现,使得蒸腾速率峰值前移,反映了其控制失水和维持体内水分平衡能力较强,是适应干旱的一种方式(张卫强等,2007)。柠条叶片蒸腾和气孔导度的日变化规律表明,蒸腾速率的日变化和叶片气孔导度、土壤水分、环境因子的变化有密切联系。气孔的运动直接调控着蒸腾速率的变化(图3),柠条气孔导度与蒸腾速率具有相同的变化规律且二者呈极显著的线性相关(r2=0.628,P<0.001)。

图3 柠条叶片蒸腾速率与气孔导度的关系Fig. 3 Relationship between leaf transpiration and stomatal conductance of Caragana korshinskii Kom.
2.2 柠条叶片蒸腾速率对土壤水分胁迫的响应
在干旱条件下,水分是最重要的限制因子,当土壤水分出现胁迫时,将引起蒸腾速率的下降。如图4所示,较高的土壤含水量并未获得较高的蒸腾速率,随着土壤含水量的逐渐降低,柠条苗木叶片蒸腾速率先迅速增加后增幅变缓,随着土壤含水量的进一步减少,蒸腾速率则开始缓慢下降。二者之间的关系经回归分析可以用 3次曲线进行描述( r2=0.861, P=0.000, n=58): Tr=-0.001(SWC)3+0.0142(SWC)2+0.1779(SWC)+1.0357,蒸腾速率随土壤含水量的变率为:dTr/dSWC=3a(SWC)2+2b(SWC)+c。令dTr/dSWC=0,得出柠条叶片蒸腾速率最高时所对应的土壤含水量为15.78%,即当土壤质量含水量≤田间持水量的74.1%时,柠条蒸腾速率随土壤供水量的增大而增大;高于此值,蒸腾速率降低。

图4 柠条叶片蒸腾速率对土壤水分胁迫的响应Fig. 4 Leaf transpiration response to soil water stress of Caragana korshinskii Kom.

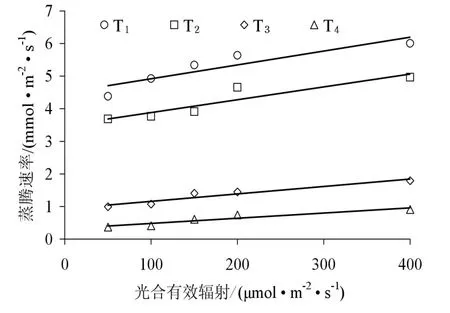
图5 不同SWC下柠条叶片蒸腾速率对光辐射强度的响应Fig. 5 Leaf transpiration response to solar radiation of Caragana korshinskii Kom. in different SWC
2.3 不同土壤水分条件下柠条叶片蒸腾速率对光合有效辐射的响应
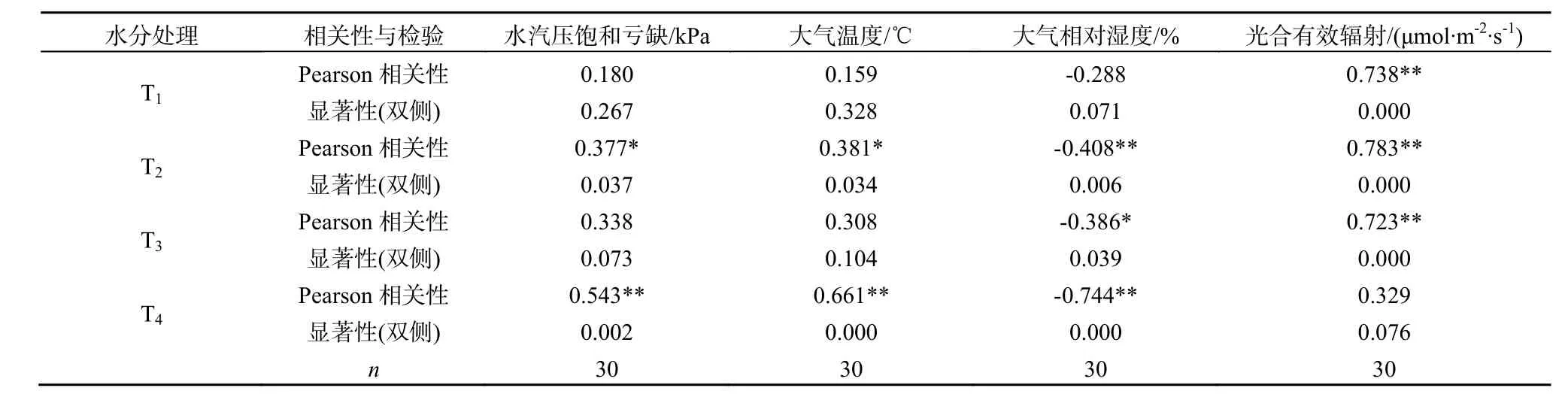
光合有效辐射为植物蒸腾提供了能量,它不仅影响树木自身的生理活动,同时也是大气温度、大气相对湿度、土壤温度等环境因子的直接控制因子(周海光等,2008)。通过设定模拟光辐射强度梯度,来观测苗木蒸腾速率对光辐射强度的响应,由图 5和图6可以看出,不同土壤水分条件下柠条叶片蒸腾速率(Tr)对光照强度的响应在启动阶段即0 表1 不同SWC下柠条叶片蒸腾速率光响应模型Table 1 Regression equation of leaf transpiration response to solar radiation in defiffent SWC 图6是低光照强度(PAR<400 μmol·m-2·s-1)下柠条叶片蒸腾作用的光响应过程。4种水分处理下,蒸腾速率随光辐射强度的增加而增加(y=kx+c),但启动速度不同,依次为 T1(k=0.0043, r2=0.772*, P=0.032)>T2(k=0.0039, r2=0.803*, P=0.025)>T3(k=0.0023, r2=0.892**, P=0.010)>T4(k=0.0016, r2=0.861*, P=0.015)。这说明在相同光照条件下,随着土壤含水量的降低,植物根系吸水速率越来越小,甚至满足不了叶片蒸腾失水的需要;当土壤水分受到限制时,植物保持较低的蒸腾速率以避免水汽过度散失导致的萎蔫。 图6 弱光下不同SWC柠条叶片蒸腾速率光响应Fig. 6 Light response of leaf transpiration under low PAR of Caragana korshinskii Kom. in different SWC 表2 不同水分条件下柠条叶片蒸腾与气象因子的相关分析Table 2 The correlation coefficients between leaf stomatal transpiration and meteorology conditions in different SWC 表3 不同水分条件下柠条叶片蒸腾与主要气象因子的回归模型Table 3 Regression models of leaf stomatal transpiration and main environmental factors 2.4 不同土壤水分条件下柠条叶片蒸腾速率与主要气象因子的相关性分析 柠条叶片蒸腾速率(Tr)与光辐射强度(PAR)、大气温度(Ta)、大气相对湿度(RH)及水汽压饱和亏缺(VPD)之间有显著的相关关系,表2和表3对各因素与叶片蒸腾速率的关系作了深入分析,不同土壤水分条件下,影响蒸腾速率的主要气象因子明显不同。Pearson相关分析(表2)得出,T1充分供水条件下,柠条蒸腾速率与 PAR极显著相关,PAR对蒸腾速率的贡献率(决定系数r2)为54.4%(P=0.000),其他气象因子的影响不显著;对于轻度水分胁迫(T2),除PAR外,其他气象因子对蒸腾速率的影响显著,但相关性不高,经偏相关分析得出(表4),柠条蒸腾速率的主要影响因子依次为:PAR(偏相关系数r=0.777,P=0.000)>VPD(偏相关系数r=0.527,P=0.004)>Ta(偏相关系数r=0.400,P=0.035)。对于中度水分胁迫(T3)下,影响柠条叶片蒸腾速率的主要气象因子为PAR和RH,偏相关系数分别为0.665(P=0.000)、-0.078(P=0.693),说明中度胁迫下,光合有效辐射强度是影响柠条叶片蒸腾速率的主要气象因子。严重水分胁迫下(T4),蒸腾速率主要影响因子为RH、Ta和VPD,根据偏相关系数绝对值的大小可知它们对柠条叶片蒸腾速率的作用从大到小依次为:VPD(r=-0.952,P=0.000)>Ta(r=0.929,P=0.000)>RH(r=0.557,P=0.000),其贡献率分别为90.6%,86.3%和 31.0%,而水汽压亏缺(VPD)是温度(Ta)和湿度(RH)综合作用的结果,说明重度胁迫下叶片内外水汽压亏缺成为调节柠条蒸腾的主导因素。由于水分胁迫明显降低了植物的蒸腾速率(图5),黄刚等人(2008)在对科尔沁沙地植物水分胁迫响应的研究表明,干旱会使植物体内水分含量降低,叶水势下降,气孔阻力加大,降低光合速率和蒸腾速率,因此研究发现干旱会引起植物出现光抑制现象,对PAR的敏感性显著降低。 表4 不同水分处理下柠条叶片蒸腾速率与主要环境影响因子的偏相关分析Table 4 Analysis of partial correlations between leaf stomatal transpiration and effective factors of C. korshinskii Kom. in different SWC 3.1 讨论 蒸腾作用是一个复杂的植物生理过程和水分运动的物理过程,不仅与植物本身的生理需水特性有关,而且与土壤有效水供给能力有关(王进鑫等,2005)。充分供水条件下的柠条苗木蒸腾速率是土壤水分严重胁迫下的 3.43倍,随着水分胁迫的加剧,叶部水分亏缺提早出现,使得蒸腾速率峰值前移,且日蒸腾过程在较为干旱时表现为双峰型,反映了其控制失水和维持体内水分平衡能力较强,是适应干旱的一种方式(张卫强等,2007)。 气孔的运动直接调控着蒸腾速率的变化,柠条气孔导度与蒸腾速率具有相同的变化规律且二者成正比关系,这与前人对沙地植物蒸腾耗水规律的研究一致(阮成江和李代琼,2007;赵明等,2003;邓继峰等,2014)。光合有效辐射是影响苗木蒸腾速率的主导因子,为植物的蒸腾提供了能量(李小磊等,2005)。不同土壤含水量下,低光强柠条叶片蒸腾作用的光响应曲线差别不大,都表现出随着光合有效辐射强度增加而缓慢上升的趋势,但启动速度随着土壤含水量的减少而降低。不同土壤水分条件下,柠条蒸腾速率的主要影响因子不尽相同。充分供水下,光合有效辐射(PAR)是主要影响因子,对柠条蒸腾速率的贡献率为54.4%;轻度水分胁迫,柠条蒸腾速率的主要影响因子依次为:PAR>VPD>Ta,其贡献率分别为 60.4%,27.8%和16.0%,PAR是影响柠条蒸腾作用的主导因子;对于中度水分胁迫,影响柠条叶片蒸腾速率的主要气象因子为PAR和RH,但光合有效辐射强度(贡献率为44.2%)仍是影响柠条叶片蒸腾速率的主导气象因子。严重水分胁迫下,蒸腾速率主要影响因子为VPD>Ta>RH,其贡献率分别为90.6%,86.3%和31.0%,而水汽压亏缺(VPD)是温度(Ta)和湿度(RH)综合作用的结果,说明柠条蒸腾速率对干旱胁迫的响应,通过光诱导气孔开放,光辐射是描述柠条蒸腾过程气孔行为的重要环境因子;随着土壤含水量的下降,气孔对PAR的敏感性降低,通过水汽压亏缺驱动气孔导度的变化,叶片内外水汽压亏缺成为调节柠条蒸腾的主导因素。 3.2 结论 (1)随着土壤水分胁迫的加剧,柠条苗木叶片蒸腾速率先增加后缓慢下降,该临界值为15.78%,即当土壤质量含水量≤田间持水量的74.1%时,柠条蒸腾速率随土壤供水量的降低而减小;高于此值,蒸腾速率同样会受到抑制。 (2)在土壤水分受到限制时(土壤质量含水量≤40%田间持水量),进一步增加光合有效辐射量(≥1000 μmol·m-2·s-1)并没有使蒸腾速率提高,严重水分胁迫下,增加光辐射强度反而导致蒸腾速率降低,此时植物保持较低的蒸腾速率以避免水汽过度散失。 (3)柠条蒸腾速率对干旱胁迫的响应,通过光诱导气孔开放,光辐射是描述柠条蒸腾过程气孔行为的重要环境因子;随着土壤含水量的下降,气孔对PAR的敏感性降低,通过水汽压亏缺驱动气孔导度的变化,叶片内外水汽压亏缺成为调节柠条蒸腾的主导因素。 FERNÁNDEZ M E, GYENGE J, SCHLICHTER T. 2009. Water flux and canopy conductance of natural versus planted forest in Patagonia, South America [J]. Trees, 23(2): 415-427. HAN L, HE K N, HU X B, et al. 2012. Canopy transpiration response to environmental variations in Platycladus orientalis: properties and modeling [J]. Pakistan Journal of Botany, 44(2): 541-545. WALLACE J, MCJANNET D. 2010. Processes controlling transpiration in the rainforests of north Queensland, Australia [J]. Journal of Hydrology, 384(1-2): 107-117. WHITLEY R, MEDLYN B, ZEPPEL M, et al. 2009. Comparing the Penman-Monteith equation and a modified Jarvis-Stewart model with an artificial neural network to estimate stand-scale transpiration and canopy conductance [J]. Journal of Hydrology, 373(1-2): 256-266. 常学向, 赵文智. 2009. 林木尺度水分利用测算方法与研究进展[J]. 冰川冻土, 31(6): 1201-1210. 邓继峰, 丁国栋, 赵媛媛, 等. 2014. 盐池地区三种典型树种蒸腾速率的研究[J]. 干旱区资源与环境, 28(7): 161-165. 黄刚, 赵学勇, 崔建垣, 等. 2008. 水分胁迫对2种科尔沁沙地植物光合和水分利用特性的影响[J]. 西北植物学报, 28(11): 2306-2313. 李清河, 刘建锋, 张景波, 等. 2006. 乌兰布和沙漠东北部8种沙生灌木生长季末期的光合生理特性[J]. 西北植物学报, 26(11): 2318-2323. 李小磊, 张光灿, 周泽福, 等. 2005. 黄土丘陵区不同土壤水分下核桃叶片水分利用效率的光响应[J]. 中国水土保持科学, 3(1): 43-47. 阮成江, 李代琼. 2007. 半干旱黄土丘陵区沙棘的光合特性及其影响因子[J]. 沙棘, 20(4): 6-9. 司建华, 常宗强, 苏永红, 等. 2008. 胡杨叶片气孔导度特征及其对环境因子的响应[J]. 西北植物学报, 28(1): 125-130. 王进鑫, 黄宝龙, 王明春, 等. 2005. 不同供水条件下侧柏和刺槐幼树的蒸腾耗水与土壤水分应力订正[J]. 应用生态学报, 16(3): 419-425. 奚如春, 马履一, 王瑞辉, 等. 2006. 林木耗水调控机理研究进展[J]. 生态学杂志, 25(6): 692-697. 徐当会, 方向文, 宾振钧, 等. 2012. 柠条适应极端干旱的生理生态机制——叶片脱落和枝条中叶绿体保持完整性[J]. 中国沙漠, 32(3): 691-697. 余峰, 陈林, 王峰, 等. 2011. 宁夏中部干旱带人工柠条茎流及蒸腾特征研究[J]. 水土保持研究, 18(2): 143-147. 张卫强,贺康宁, 朱艳艳, 等. 2007. 黄土半干旱区油松苗木蒸腾特性与影响因子的关系[J]. 中国水土保持科学, 5(1): 49-54. 赵明, 李爱德, 王耀琳, 等. 2003. 沙生植物的蒸腾耗水与气象因素的关系研究[J]. 干旱区资源与环境, 17(6): 131-137. 赵平. 2011. 整树水力导度协同冠层气孔导度调节森林蒸腾[J]. 生态学报, 31(4): 1164-1173. 周海光, 刘广全, 焦醒, 等. 2008 黄土高原水蚀风蚀复合区几种树木蒸腾耗水特性[J]. 生态学报, 28(9): 4568-4574. Research on Leaf Transpiration Response to Soil Water Stress of Caragana korshinskii Kom. in Hedong Sandy Land of Ningxia HAN Lei1*, SUN Zhaojun1, ZHAN Xiuli2, JIAO Bingzhong3, DONG Liang1 Caragana korshinskii Kom., a type of psammophilic shrub growing in Hedong sandy land of Ningxia was transplanted in pots in a greenhouse and treated with progressive water stress. The leaf transpiration characteristics under different soil water condition were studied to reveal the response mechanism of plant leaf transpiration to main environmental factors. It is expected that the study will provide the scientific basis for optimizing forest structure allocation of regional vegetation restoration and reconstruction in arid region. It was shown that: (1) The transpiration rate of C. korshinskii Kom. under the adequate soil water supply is exactly 3.43 times than that in severe water stress. The transpiration peak moved forward while increasing of soil water stress, and the daily transpiration rate varied in a double-peak curve under the severe drought stress. There was a linear correlation between leaf transpiration (Tr) and stomatal conductance, both were the same dynamics under different soil water condition. (2) As the soil water content reduction, the transpiration rate of C. korshinskii Kom., after reaching its peak figure, will slowly decrease. The critical value of soil water content (SWC) was 15.78% (that was 74.1% of field moisture capacity). (3) Light response curves of leaf transpiration under low photosynthetically available radiation (PAR) in different SWC made little different. The leaf transpirations under low solar radiation were up with linear while increasing of PAR. However, the start-up speeds of Trwere decrease with soil water content reduction. Under the soil water stress (SWC≤40% of field moisture capacity), any further increase in PAR (≥1 000 μmol·m-2·s-1), however, was not helpful to improve Tr. Meanwhile, it was arguable that increase the light radiation intensity led to decreased transpiration rate under the severe soil water stress. And (4) after analysis of partial correlations, the PAR was the major factor influencing changes of Trunder the adequate soil water supply, contributing 54.4% of the variation Tr. But with the decline in soil moisture, the sensitivity of stomata decreased to PAR. The vapour pressure deficit between air and leaf became the dominant factor in regulating transpiration process of C. korshinskii Kom.. Caragana korshinskii Kom.; leaf transpiration; soil water stress; sandy land 10.16258/j.cnki.1674-5906.2015.05.005 Q945.78 A 1674-5906(2015)05-0756-06 韩磊,孙兆军,展秀丽,焦炳忠,董良. 宁夏河东沙区柠条植株叶片蒸腾对干旱胁迫的响应[J]. 生态环境学报, 2015, 24(5): 756-761. HAN Lei, SUN Zhaojun, ZHAN Xiuli, JIAO Bingzhong, DONG Liang. Research on Leaf Transpiration Response to Soil Water Stress of Caragana korshinskii Kom. in Hedong Sandy Land of Ningxia [J]. Ecology and Environmental Sciences, 2015, 24(5): 756-761. 宁夏自然科学基金项目(NZ13025);国家自然科学基金项目(31460220;31340018;41201087) 韩磊(1985年生),男,博士,副研究员,主要从事水土保持与荒漠化防治、林业生态工程研究。E-mail:layhan@163.com 2015-02-06


3 结论与讨论
1. Research Center of New Technology Application and Development, Ningxia University, Yinchuan 750021, China; 2. School of Recourses and Environment, Ningxia University, Yinchuan 750021, China; 3. School of Civil engineering and water conservancy, Ningxia University, Yinchuan 750021, China
