水稻品种对CH4产生、排放及δ13CH4的影响
2015-12-06张文宣于海洋张广斌马静徐华
张文宣,于海洋,张广斌,马静,徐华*
1. 土壤与农业可持续发展国家重点实验室,中国科学院南京土壤研究所,江苏 南京210008;2. 中国科学院大学,北京100049
水稻品种对CH4产生、排放及δ13CH4的影响
张文宣1,2,于海洋1,2,张广斌1,马静1,徐华1*
1. 土壤与农业可持续发展国家重点实验室,中国科学院南京土壤研究所,江苏 南京210008;2. 中国科学院大学,北京100049
水稻品种是调控CH4产生和排放的关键因素。关于水稻品种对稻田产生和排放CH4的稳定性碳同位素组成(δ13CH4)的影响研究鲜见报道。通过温室盆栽和室内培养试验并结合稳定性碳同位素方法,研究了持续淹水条件下4个水稻生育期镇稻624、农香98和中早33的土壤CH4产生潜力、土壤溶液CH4浓度、CH4排放通量及产生、排放CH4的δ13C值,为最终筛选优质高产且低CH4排放的水稻品种提供CH4排放相关过程及其稳定性碳同位素方面的参考数据。结果表明:在分蘖期和拔节期,镇稻624和农香98的土壤CH4产生潜力显著高于中早33,在灌浆期和成熟期显著小于后者(P<0.05)。三者土壤CH4产生潜力、土壤溶液CH4浓度最高值和土壤Eh的最低值依次出现在拔节期(2.6 μg·g-1·d-1,346.9 μmol·L-1,-296 mV)、拔节期(3.2 μg·g-1·d-1,425.9 μmol·L-1,-316 mV)和灌浆期(2.4 μg·g-1·d-1、435.2 μmol·L-1,-308 mV)。各品种土壤CH4产生潜力均与相应土壤溶液中CH4浓度显著正相关(P<0.01),且与土壤Eh显著负相关(P<0.01)。镇稻624和农香98在分蘖盛期CH4排放通量最大(67.1和68.7 mg·m-2·h-1),中早33则在拔节期(58.5 mg·m-2·h-1)。各品种CH4季节排放总量依次为55.29、55.74和40.82 g·m-2,前二者无显著差异,显著高于中早33,这可能是镇稻624和农香98的土壤CH4产生潜力在分蘖期和拔节期显著大于中早33,而各品种CH4排放又相对集中在分蘖期和拔节期的缘故。相关分析表明,各生育期CH4排放通量与相应的土壤CH4产生潜力显著正相关(P<0.01)。可见水稻品种通过影响土壤的CH4产生,进而影响稻田CH4的排放。镇稻624和中早33土壤产生CH4的δ13C值从约-67.0‰增至-55.5‰,农香98则先减后增,范围为-64.2‰~-52.9‰,这说明镇稻624和中早33的土壤CH4产生途径差异较小,而二者与农香98差异较大。各品种排放CH4的δ13C值均先减后增,分别为-67.6‰~-48.5‰、-73.0‰~-47.3‰和-60.9‰~-46.7‰,季节平均值依次为-52.7‰、-52.5‰和-54.8‰。总体上,水稻品种影响排放δ13CH4值的季节变化。
水稻品种;CH4产生;CH4排放;δ13C;土壤Eh
全球气候变暖问题是人类社会当前所面临的最严峻挑战之一,最主要原因是化石燃料燃烧和土地利用变化所导致的大气温室气体浓度的增加(IPCC,2014;IPCC,2013;沈永平和王国亚,2013;IPCC,2007)。甲烷(CH4)是仅次于二氧化碳(CO2)的重要温室气体,其对全球温室效应的贡献为18%(IPCC,2007)。2012年大气 CH4浓度达到 1.819 μL·L-1,比工业化前高出160%,为近80万年来最高(IPCC,2013;沈永平和王国亚,2013)。稻田是大气 CH4的重要排放源,约占全球总排放量的5%~19%(IPCC,2007)。如何有效地进一步减小甚至阻止稻田向大气排放包括CH4在内的温室气体,业已成为全球亟需解决的科学问题。
稻田CH4排放是产生、氧化和传输综合作用的结果,水稻植株强烈影响稻田CH4排放。首先,水稻根系分泌物和脱落物为产甲烷菌提供丰富的碳源和能源,促进 CH4的产生(Vandergon和Vanbreemen,1993;Kludze等,1993;Jia等,2001;Wang等,1997);其次,水稻植株将大气中的 O2传输到植株根系以维持水稻生长,形成根际氧化区,有利于稻田CH4氧化(Frenzel等,1992;Yagi和Minami;1991);最后,水稻植株的通气组织是稻田 CH4排放的最重要通道(Schutz等,1989;Vandergon和Neue,1995),加速稻田土壤中产生的CH4迅速释放到大气。因此,深入研究水稻植株对稻田CH4排放过程,特别是对CH4产生的影响有助于为寻求减缓稻田CH4排放的对策提供科学依据和理论参考。
水稻品种是调控CH4产生和排放的关键因素。不同品种随着水稻生长期的变化,生理和形态特征都会发生明显的改变,从而对稻田CH4排放的影响不同(Ma等,2010),种植和选育合适的水稻品种可减少CH4排放量达20%~30%(陶战等,1994)。Jia等(2006)通过一年的温室盆栽实验研究不同水稻品种(盐选,72031,9516)对稻田 CH4排放的影响,发现整个稻季,盐选水稻品种甲烷排放量最高(5.88 μg·m-2·h-1),其次是 72301品种(4.48 μg·m-2·h-1)和9516品种(3.41 μg·m-2·h-1)。目前有关水稻品种对稻田CH4排放通量影响的研究已有大量报道(Schutz等,1989;Vandergon和Neue,1995;Sass和Fisher,1995),但关于它们对稻田CH4产生的影响的报道还很少(Conrad等,2002;Jia等,2006;Zhang等,2013)。更重要的是,不同水稻品种的分泌物和凋落物种类和含量可能存在差异,从而导致土壤产CH4的主要途径(乙酸发酵和H2/CO2还原)可能并不一样,进而使得产生的CH4的稳定性碳同位素组成(δ13CH4)会有所区别。此外,不同水稻品种的 CH4氧化和传输能力也存在很大差异,最终导致排放CH4的δ13CH4值也有所不同。然而关于这些方面的研究尚鲜见报道(张晓艳等,2010)。
我国是水稻生产大国,水稻种植面积大、品种多。本试验选用镇稻624、农香98和中早33为研究对象,通过盆栽实验和室内培养实验观测土壤的CH4产生潜力、土壤溶液CH4浓度、CH4排放通量以及它们的 δ13CH4值,旨在研究水稻品种对稻田CH4排放过程,特别是对产生和排放CH4的δ13CH4值的影响,为最终筛选优质高产且低CH4排放的水稻品种提供CH4排放相关过程及其稳定性碳同位素方面的参考数据。
1 材料与方法
1.1 试验设计
供试土壤采自江苏省句容市白兔镇,母质为发育于下蜀黄土的爽水性水稻土,土壤(0~15 cm土层)有机碳质量分数为18.9 g·kg-1,全氮质量分数为1.2 g·kg-1,pH值为6.91,土壤有机质的稳定性碳同位素组成为-26.8‰。试验为盆栽试验,盆钵为圆柱体,直径25 cm,高35 cm,每盆装16 kg风干土。设置3个处理:中早33、镇稻624和农香98,每处理3次重复。各盆钵于2012年6月29日淹水,5月16日育苗,6月30日移栽。每个盆钵3穴,每穴1株秧苗。中早33、镇稻624和农香98分别于9月17日、10月21日和11月6日收获。整个水稻生长期,各盆钵均维持水层在2 cm以上,直至水稻收获。
施用尿素共3.74 g,按m(基肥)∶m(分蘖肥)∶m(穗肥)=2∶1∶1施用,磷、钾肥分别为0.84 g的过磷酸钙和0.24 g的氯化钾,作为基肥一次性施入。中早33的基肥、分蘖肥、穗肥分别于6月29日、7月13日、8月1日施用。镇稻624的基肥、分蘖肥、穗肥分别于6月29日、7月13日、8月17日施用。农香98的基肥、分蘖肥、穗肥分别于6月29日、7月13日、8月23日施用。
1.2 样品采集和测定
CH4气样用静态箱法采集。水稻生长期每隔3~ 4 d采样1次,采样时间为09:00—11:00,每15 min采样1次,共采4次。为测定排放δ13CH4值,观测到CH4排放后每隔10~15 d采样1次,开始和结束时所采样品瓶内气体测定完 CH4浓度后测定其δ13CH4值。采样结束后移走采样箱。在采气的同时,用数字温度计(Model 2455, Yokogawa, Japan)记录箱温及10 cm处土温,通过便携式土壤Eh计测定10 cm处土壤Eh。气样CH4浓度用带有氢离子火焰检测器的气相色谱(岛津 GC-12A)分析。根据瓶内CH4浓度与时间的关系曲线计算CH4排放通量。
CH4的稳定性碳同位素组成用带有全自动预气相色谱(GC)浓缩接口(PreCon)的 Thermo FinniganMAT-253同位素质谱仪测定(曹亚澄等,2008)。PreCon接口是一种全自动预GC浓缩接口,包括样品瓶、化学阱、冷阱、燃烧反应器、六通阀5个部分。向抽真空的玻璃瓶样品瓶中注入100 mL气体样品,如果气样量少于 100 mL,须充入不含CH4的惰性气体,使瓶内呈常压。CH4被氧化成CO2和H2O,随后由CH4产生的CO2流入GC进行进一步地分离。所用 CO2参比气体经过标定(代号为Lab.Gas,纯度为99.999%,δ13CPDB值为-23.73‰)。根据标准CO2峰和样品峰的m/z44、m/z45和m/z46 3种离子流强度的比值,即可得出由样品中CH4转化成的 CO2相对于国际碳同位素 PDB的值,即δ13CPDB‰。
土壤溶液采用根际土壤溶液采集器(Rhizon soil moisture sampler,简称Rhizon SMS)采集(Zhang等,2011)。盆钵淹水前将采集器平放在土壤25 cm处直至水稻收获。每次取样之前,先用18 mL真空瓶抽取约5 mL的土壤溶液以冲洗取样器,然后再用另一只18 mL真空瓶采取约10 mL样品。随后将样品瓶内充入适量纯 N2,使之维持在 1 atm(1 atm=101.325 kPa)。震荡、取气,待测 CH4浓度(Zhang等,2013)。
水稻主要生育期(分蘖期、拔节期、灌浆期和成熟期),将盆钵中水稻植株拔出,尽量全部取下根系上附着的土壤,仔细挑选并去掉残留在土壤中的水稻根系和其他外来物质,混合均匀。取出约50 g干质量的土壤,加入无氧去离子水,搅拌成泥浆状,使瓶内土水比为1∶1。将泥浆样品迅速转移到体积为250 mL的培养瓶中。培养瓶充N2后用硅橡胶塞密封,进行厌氧培养试验。各生育期培养试验时间如表1。
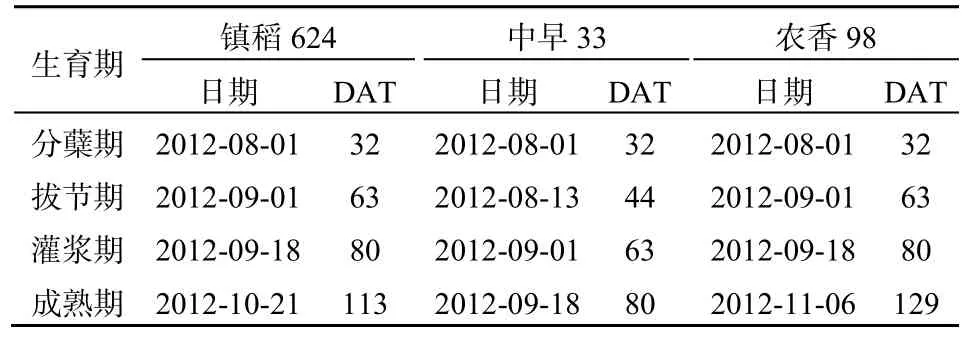
表1 培养实验时间安排Table 1 Cultivation experiments schedule at different growth stages
1.3 CH4产生潜力的厌氧培养试验
培养瓶经反复6次抽真空、充N2(最后充N2至 1 atm)后,参照实测土温置于暗箱培养。CH4产生潜力通过测定密闭培养 1 h(作为初始值)和50 h(作为最终值)后瓶内 CH4浓度的变化获得(Wang等,1997)。培养实验结束后,测定培养瓶内CH4气样的δ13CH4值。
1.4 土壤溶液中CH4浓度的计算
土壤溶液中 CH4浓度用下式计算获得:C=(m×Gv)/(GL×Mv),式中:C为土壤溶液中 CH4浓度(µmol·L-1);m为瓶中气相CH4的浓度(µL·L-1);Gv为瓶中气相的体积(L);GL为瓶中水样的体积(L);Mv为25 ℃下理想气体的摩尔体积,即24.47 L·mol-1。
1.5 排放δ13CH4值的计算
排放的 δ13CH4值由以下公式计算:F=[(B×b)-(A×a)]/(B-A),式中:F为排放的δ13CH4值;A和B分别为开始和结束采气箱内气样中CH4浓度(µL·L-1);a和b分别为开始和结束采气箱内气样中相应的δ13CH4值(‰)。
2 结果与分析
2.1 水稻品种对CH4产生的影响
各品种的土壤在分蘖期均有明显的 CH4产生(图1a),镇稻624和农香98的土壤CH4产生潜力增加较快,在拔节期出现峰值2.6和3.2 μg·g-1·d-1,而中早33在灌浆期才达到最大值2.4 μg·g-1·d-1。总体而言,农香98在各生育期土壤的CH4产生潜力均大于镇稻624,但仅在拔节期和灌浆期达到显著差异水平(P<0.05)。在分蘖期和拔节期,镇稻624和农香98土壤的CH4产生潜力显著高于中早33,但在随后的灌浆期和成熟期急剧减小,明显小于中早33(P<0.05)。尤其在灌浆期,中早33显著大于农香 98,农香 98显著大于镇稻 624(P<0.05)。
随着水稻的生长,各品种的土壤Eh先减再增(图1f)。镇稻624和农香98的土壤Eh在拔节期出现最低值,依次为-296和-316 mV,而中早33在灌浆期降至最低值-308 mV。各生育期,镇稻 624和农香98的土壤Eh均无显著差异。仅在灌浆期,中早33的土壤Eh显著小于镇稻624和农香98。各品种的土壤Eh与相应土壤CH4产生潜力的时间变化趋势一致(图1a和图1f),即分别在拔节期、拔节期和灌浆期出现最低值。统计分析表明,土壤的CH4产生潜力与土壤Eh显著负相关(镇稻624:r=-0.889,P<0.01;中早33:r=-0.787,P<0.01;农香98:r=-0.990,P<0.01)。
镇稻624、农香98和中早33的土壤溶液CH4浓度与对应的土壤 CH4产生潜力的变化趋势一致(图1a和图1c),分别在拔节期和灌浆期达到最大值(346.9、425.9和435.2 μmol·L-1)。在分蘖期和拔节期,各品种的土壤溶液CH4浓度差异不明显,在灌浆期,中早33的CH4浓度显著大于农香98,农香98显著大于镇稻624(P<0.05)。统计分析表明:土壤溶液CH4浓度与土壤CH4产生潜力显著正相关(镇稻624:r=0.743,P<0.01;中早33:r=0.665,P<0.05;农香98:r=0.832,P<0.01)。
CH4是严格厌氧条件下产甲烷菌作用于产甲烷底物的结果(Conrad,2007)。水稻生长前期,持续淹水使土壤Eh均处于较适宜产CH4的水平。随移栽时间延长,可能由于水稻植株的通气组织逐渐发育,大气中的氧气进入根部区域形成一个很强的氧化微域,进而氧化土壤复杂有机物形成大量的简单有机物质,各品种水稻土壤的CH4产生潜力逐渐增强,均在水稻前、中期较大。由于生长前期产甲烷菌消耗了大量底物,且土壤Eh不断增大,各品种土壤CH4产生潜力逐渐减小,在成熟期均降至最小。镇稻624和农香98土壤CH4产生潜力和土壤Eh表现出基本相同的规律(图1a和图1f),二者的土壤Eh均在拔节期出现最小值-296和-316 mV(图1f),此时它们土壤的CH4产生潜力达到峰值(图1a)。中早33的土壤Eh在灌浆期才降至最低值-308 mV,此时其土壤CH4产生潜力亦最大。土壤Eh越低,相应生育期的土壤CH4产生潜力就越高,且统计分析表明,各品种土壤的CH4产生潜力与土壤 Eh显著负相关,表明土壤 Eh是影响稻田CH4产生的重要因素。对于中早33而言,可能是由于该品种水稻植株发育较快,尽管其土壤CH4产生潜力和土壤 Eh最值出现的生育期均晚于镇稻 624和农香98,但达到峰值的时间均在水稻移栽后63 d(表1)。
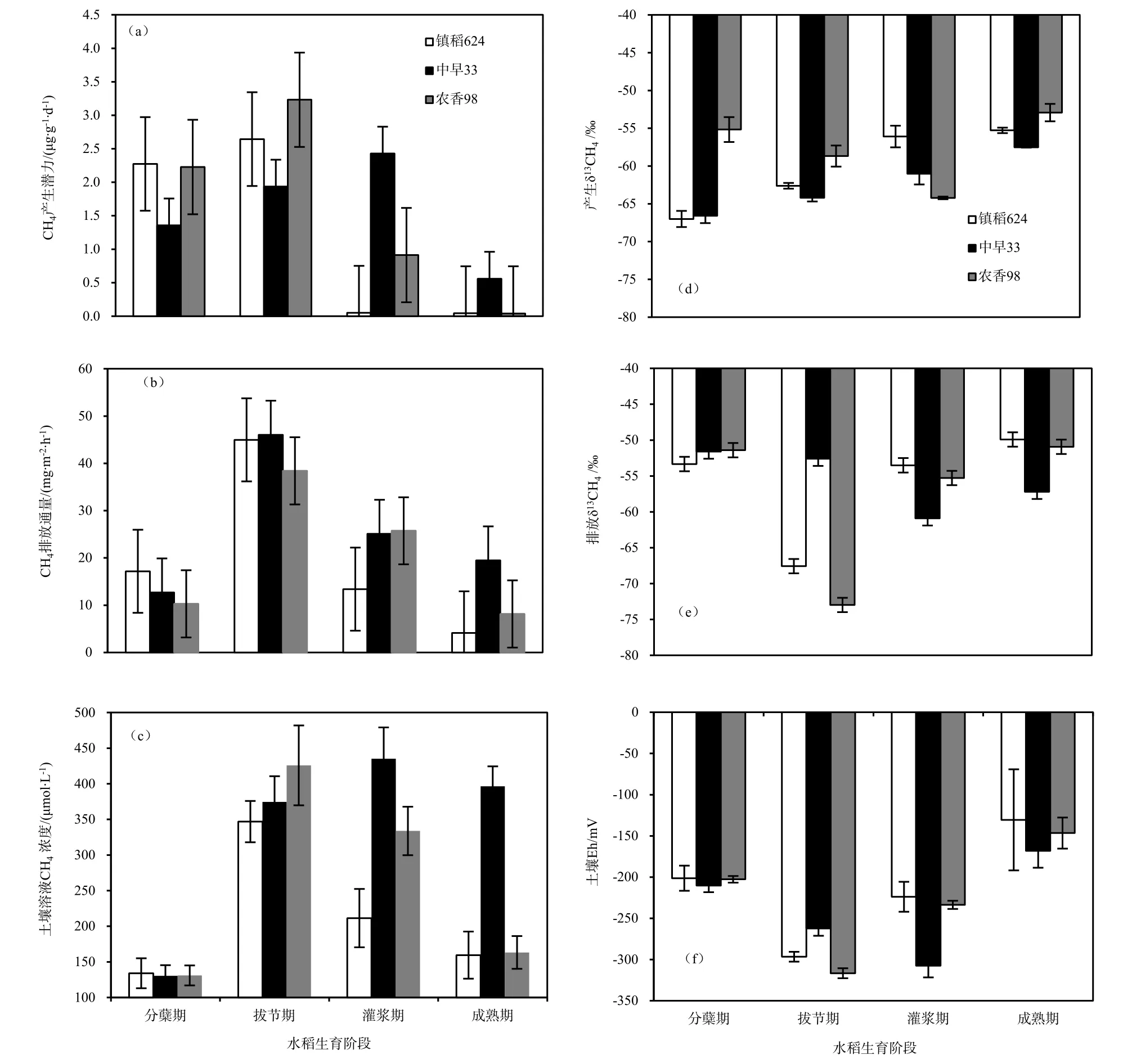
图1 各生育期CH4产生潜力、土壤溶液CH4浓度、CH4排放通量及δ13CH4和土壤EhFig. 1 CH4production potential, CH4conecentration in soil solution, CH4flux , δ13CH4and soil Eh at four stages
未被氧化且未立即释放到大气中的CH4以闭蓄和溶解的形式保留在土壤中。在拔节期镇稻624和农香98的土壤CH4产生潜力最大,其土壤溶液CH4浓度最大(347、426 μmol·L-1),在灌浆期中早33土壤CH4产生潜力最大,其土壤溶液CH4浓度亦最大(435 μmol·L-1)(图 1c),这至少可以表明土壤的CH4产生潜力是影响土壤溶液CH4浓度的重要因素,土壤CH4产生潜力越大,土壤溶液CH4浓度相应越高,这与张广斌等(2013)大田实验结果一致。统计分析发现,土壤的CH4产生潜力与土壤溶液中 CH4浓度显著正相关(镇稻 624:r=0.743,P<0.01;中早33:r=0.665,P<0.05;农香98:r=0.789,P<0.01),这充分表明,土壤溶液CH4浓度是土壤CH4产生潜力重要的指示因子。
2.2 水稻品种对产生δ13CH4的影响
镇稻624和中早33土壤产生的CH4逐渐变重,逐渐相对富集13C,它们的δ13CH4值总体上不断增大,范围为-67.0‰~-55.5‰左右(图1d),各生育期二者无显著差异(P>0.05)。农香98土壤产生CH4的 δ13CH4值则先减小后增大,在灌浆期降至最低(-64‰),范围为-64.2‰~-52.9‰左右。在分蘖期,农香98土壤产生CH4的δ13CH4值显著大于镇稻624和农香98。这说明镇稻624和中早33的土壤CH4产生途径差异较小,而它们与农香98的土壤CH4产生途径差异较大。
产生的 δ13CH4主要取决于产甲烷底物的类型以及 CH4产生途径(乙酸和 H2/CO2)的相对贡献率(Conrad等,2002;Krüger等,2002)。本试验不添加任何外源有机质,且土壤有机碳本身的13C相对稳定(Zhang等,2013),因此,土壤产生CH4的 δ13CH4值大小存在差异,且随水稻生长不断变化,主要是由于它们的CH4产生途径不同且受水稻植株生长的影响(Zhang等,2013;Krüger等,2002)。与H2/CO2还原产生的CH4相比,乙酸形成的CH4相对富集13C,其δ13CH4值较大(Sugimoto和Wada,1993;Whiticar,1999)。张广斌等(2012)田间试验研究发现,在持续淹水条件下乙酸产CH4途径贡献率存在明显的季节变化,由分蘖期时的70%以上逐渐下降到成熟期时的50%左右,恰好相反,Krüger等(2002;2001)的研究结果表明,乙酸产CH4仅在水稻后期处于主导地位,并认为这可能与实验的水稻品种及其生长状况有关。随着淹水时间延长,镇稻624和中早33品种土壤产生的CH4逐渐富集13C。这一方面表明,土壤本底有机质在持续淹水条件下产生的产甲烷底物—乙酸可能逐渐增多,另一方面也预示了随着水稻植株的生长,根系分泌物乙酸的含量也可能逐渐增加,导致产生CH4的δ13CH4值不断增大。以往研究发现,不同水稻品种水稻根系分泌物的种类和含量确实并不一致(林敏和尤崇杓,1989)。遗憾的是,本实验并未同时观测土壤和根系分泌的乙酸含量,相关研究尚有待进一步完善与探讨。尽管镇稻624和中早33品种土壤的CH4产生潜力在4个主要生育期都明显不同(图1d),但相较于土壤中大量的产甲烷底物,少量的根系分泌物和脱落物使得它们的CH4产生途径差异可能都很小,使得它们产生的δ13CH4值比较接近。在分蘖期,农香98土壤产生CH4的δ13CH4值显著大于镇稻624和中早33,这可能是由于此时植株生长发育稍迟缓的农香98营养生长消耗较少的土壤有机质,与之竞争的产甲烷菌对乙酸的利用率因此更高。随着水稻不断生长,植株营养生长和生殖生长加强,同时根系分泌物和脱落物含量不断增加,乙酸产CH4途径的贡献率降低,产生CH4的δ13CH4值不断减小。在成熟期,植株的生殖生长逐渐减弱,根系分泌物和脱落物中乙酸含量增加。土壤产生CH4又逐渐富集13C,其δ13CH4值有所上升。这总体可以说明镇稻624和中早33土壤CH4产生途径差异较小,而它们与农香98的土壤CH4产生途径差异较大。
2.3 水稻品种对CH4排放的影响
3个品种的CH4排放通量的季节变化趋势比较相似,但镇稻624和农香98均在分蘖盛期达到最大,为67.1和68.7 mg·m-2·h-1(图2a和图2c),而中早 33的峰值出现在拔节期,为 58.5 mg·m-2·h-1(图 2b),随后均逐渐减小直至水稻收获。供试品种的CH4排放均集中在分蘖期和拔节期,镇稻624在这两个生育期的CH4排放量依次占总量的43.2%和42.8%,农香98占总量的30.6%和36.4%,中早33则占22.0%和42.9%(表2)。全观测期内,镇稻624、农香98和中早33的CH4季节排放总量分别为55.29、55.74和40.82 g·m-2,中早33显著低于镇稻624和农香98(P<0.05)。相关分析表明,各生育期CH4排放通量与相应的土壤CH4产生潜力显著正相关(镇稻 624:r=0.652,P<0.01;中早 33:r=0.561,P<0.01;农香33:r=0.678,P<0.01)。
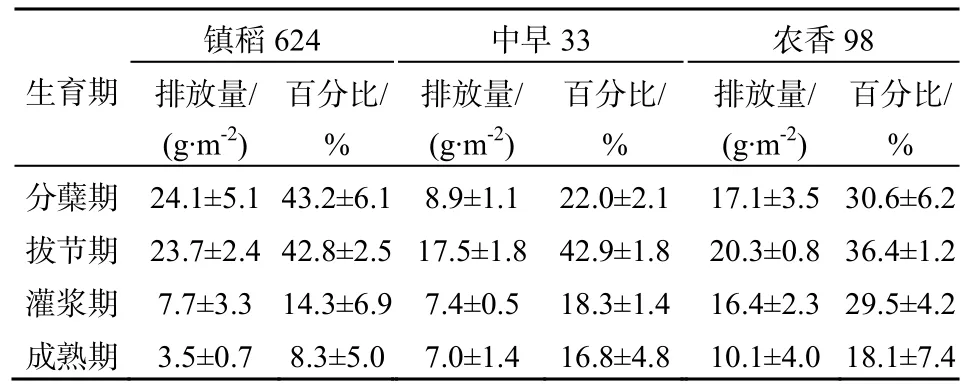
表2 不同水稻生育期累积排放量Table 2 Accumulative CH4emissions at different growth stages
现有研究表明,水稻品种显著影响稻田CH4排放通量(Jia等,2006;林而达,1993)。CH4的产生是CH4排放的基础,本实验统计发现,4个生育期,各品种土壤的CH4产生潜力与CH4排放通量均显著正相关。就季节 CH4排放总量来看,中早 33显著低于镇稻624和农香98,一方面可能与其较短的生育期有关;另一方面可能是由于在分蘖期和拔节期,镇稻624和农香98的土壤CH4产生潜力显著大于中早33(图1a),此时三者CH4排放所占整个生育期的比例较大(表 2),而在灌浆期和成熟期,尽管镇稻624和农香98的土壤CH4产生潜力显著小于中早33,但此时CH4排放所占比例均已较小。Jia等(2002)研究也发现,稻田CH4排放通量与土壤CH4产生速率及水稻植株传输速率成正比,而与根际的CH4氧化成反比。这充分表明水稻品种通过影响土壤的CH4产生,进而影响稻田CH4的排放。
2.4 水稻品种对排放δ13CH4的影响
随着水稻生长,各品种排放的CH4先贫13C后富13C,其δ13CH4值总体上均先减小后增大。镇稻624和农香98排放CH4的δ13CH4值变化趋势相似,均在拔节期较其他生育期相对偏负,而中早 33在灌浆期相对偏负(图1e,图2d、图2e、图2f)。整个观测期各品种排放 δ13CH4值分别为-67.6‰~-48.5‰(镇稻 624)、-73.0‰~-47.3‰(农香98)和-60.9‰~-46.7‰(中早33),季节平均值依次为-52.7‰、-52.5‰和-54.8‰。在拔节期,中早33排放CH4的δ13CH4值显著大于镇稻624和农香98,而在成熟期显著小于后两者(P<0.05)。总体而言,水稻品种影响排放δ13CH4值的季节变化。

图2 CH4排放通量、排放δ13CH4、土壤Eh的季节变化Fig. 2 Seasonal variations of CH4flux, δ13C-value of emitted CH4, soil Eh
稻田CH4排放是土壤中CH4的产生、氧化和向大气传输3个过程共同作用的结果。稻田CH4的产生、氧化和传输这3个过程均存在同位素分馏,即产甲烷菌优先利用12C、甲烷氧化菌优先氧化12CH4、12CH4能更快地被传输进入大气(Tyler等,1997)。因此,排放的δ13CH4值受产CH4底物的组成、CH4产生途径、CH4氧化程度和传输等综合过程的影响(Uzaki等,1991;Marik等,2002)。由于 CH4氧化细菌不仅消耗内源产生的 CH4以减少CH4排放,还能使得剩余下来未被氧化的CH4富集13C,即排放δ13CH4值增大(Chanton等,1997;Bilek等,1999)。与氧化作用相反,植株传输则使排放δ13CH4值减小(Chanton,2005)。因此,CH4被氧化的越少且被传输的越快,则将导致CH4排放通量越大,排放的δ13CH4值越小。镇稻624和中早33品种排放的CH4分别在分蘖期、分蘖期和拔节期较产生的CH4明显富集13C,而农香98排放CH4的δ13CH4值与产生CH4的δ13CH4值相差较小(图1d、图1e),这可能是因为对于镇稻624和中早33而言,CH4在排放过程中可能受到了强烈的 CH4氧化作用,而对于农香 98品种而言,因生长发育相对迟缓,植株通气组织尚不发达,CH4在排放过程中受到的CH4氧化作用较弱。随后,镇稻624和农香98排放的CH4分在拔节期最贫13C,中早33则在灌浆期最贫13C(图1d、图1e),这可能是由于它们的CH4氧化百分率在此时减小而植株传输能力增强,从而导致排放的δ13CH4值变小。在成熟期,3个品种排放的CH4均又逐渐富集13C(图1d、图1e),这可能与该生育期土壤产生 CH4的δ13CH4值逐步增大有关(图1d)。尽管供试水稻品种产生CH4的δ13CH4值存在较大差异,但排放CH4的 δ13CH4值的季节平均值总体上比较接近,这表明CH4氧化和传输产生的同位素分馏效应弥补了CH4产生对排放δ13CH4值的影响。
3 结论
在分蘖期和拔节期,镇稻624和农香98的土壤CH4产生潜力显著高于中早33, 在灌浆期和成熟期显著小于后者。镇稻624和农香98在分蘖盛期CH4排放通量最大(67.1和68.7 mg·m-2·h-1),中早33在拔节期(58.5 mg·m-2·h-1)。各品种CH4季节排放总量依次为55.29、55.74和40.82 g·m-2,前二者无显著差异,显著高于中早33。各生育期CH4排放通量与相应的土壤CH4产生潜力显著正相关。可见水稻品种通过影响土壤的 CH4产生,进而影响稻田CH4的排放。
镇稻624和中早33土壤产生CH4的δ13C值从约-67.0‰增至-55.5‰,农香98先减后增,范围为-64.2‰~-52.9‰,说明镇稻 624和中早 33的土壤CH4产生途径差异较小,而二者与农香 98差异较大。各品种排放CH4的δ13C值均先减后增,分别为-67.6‰~-48.5‰、-73.0‰~-47.3‰和-60.9‰~-46.7‰。总体上,水稻品种影响排放δ13CH4值的季节变化。
BILEK R S, TYLER S C, SASS R L, et al. 1999. Differences in CH4oxidation and pathways of production between rice cultivars deduced from measurements of CH4flux and δ13C of CH4and CO2[J]. Global Biogeochemical Cycles, 13(4): 1029-44.
CHANTON J P, WHITING G J, BLAIR N E, et al. 1997. Methane emission from rice: Stable isotopes, diurnal variations, and CO2exchange [J]. Global Biogeochemical Cycles, 11(1): 15-27.
CHANTON J P. 2005. The effect of gas transport on the isotope signature of methane in wetlands [J]. Organic Geochemistry, 36(5): 753-68.
CONRAD R, KLOSE M, CLAUS P. 2002. Pathway of CH4formation in anoxic rice field soil and rice roots determined by13C stable isotope fractionation [J]. Chemosphere, 47(8): 797-806.
CONRAD R. Microbial ecology of methanogens and methanotrophs [J]. Advances in Agronomy, 2007, 96:1-63.
FRENZEL P, ROTHFUSS F, CONRAD R. 1992. Oxygen Profiles and Methane Turnover in a Flooded Rice Microcosm [J].Biology and Fertility of Soils, 14(2): 84-9.
PCC. 2007. Climate change 2007: impacts, adaptation, and vulnerability [M].Cambridge: Cambridge University Press: 1-976.
IPCC. Climate change 2014: impact, adaptation, and vulnerability [M/OL]. Cambridge:Cambridge University Press, in press, 2014[2014-05-06]. http://www.ipcc.ch/report/ar5/wg2/.
IPCC.Climate change 2013: the physical science basis [M/OL].Cambridge: Cambridge University Press, in press, 2013 [2014-05-06].http://www. ipcc.ch/report/ar5/wg1/.
JIA Z J, CAI Z C, TSURUTA H R. 2006. Effect of rice cultivar on CH4production potential of rice soil and CH4emission in a pot experiment [J]. Soil Science and Plant Nutrition, 52(3): 341-8.
JIA Z J, CAI Z C, XU H, et al. 2001. Effect of rice plants on CH4production, transport, oxidation and emission in rice paddy soil [J]. Plant Soil, 230(2): 211-21.
细胞培养:PC3 细胞株用含10%新生牛血清、青霉素(100 U/L)及链霉素(100 U/L) 的 DMEM培养基中,放置在室温37 ℃、湿度为5%CO2的孵箱中培养,3~4天传代1次。
JIA Z J, CAI Z C, XU H, et al. 2002. Effects of rice cultivars on methane fluxes in a paddy soil [J]. Nutrient Cycling in Agroecosystems, 64(1/2): 87-94.
KLUDZE H K, DELAUNE R D, PATRICK W H. 1993. Aerenchyma Formation and Methane and Oxygen-Exchange in Rice [J]. Soil Science Society of America Journal, 57(2): 386-91.
KRÜGER M, ELLER G, CONRAD R, et al. 2002. Seasonal variation in pathways of CH4production and in CH4oxidation in rice fields determined by stable carbon isotopes and specific inhibitors [J]. Global Change Biology, 8(3): 265-80.
KRÜGER M, FRENZEL P, CONRAD R. 2001. Microbial processes influencing methane emission from rice fields [J]. Global Change Biology, 7(1): 49-63.
MA K, QIU Q F, LU Y H. 2010. Microbial mechanism for rice variety control on methane emission from rice field soil [J]. Global Change Biology, 16(11): 3085-95.
MARIK T, FISCHER H, CONEN F, et al. 2002. Seasonal variations in stable carbon and hydrogen isotope ratios in methane from rice fields [J]. Global Biogeochemical Cycles, 16(4): 1094.
SASS R L, FISHER F M. 1995. Methane emissions from Texas rice fields: A five-year study [M]. Berlin 33: Springer-Verlag Berlin.
SUGIMOTO A, WADA E. 1993. Carbon Isotopic Composition of Bacterial Methane in a Soil Incubation Experiment - Contributions of Acetate and CO2/H2[J]. Geochimica et Cosmochimca Acta, 57(16): 4015-27.
TYLER S C, BILEK R S, SASS R L, et al. 1997. Methane oxidation and pathways of production in a Texas paddy field deduced from measurements of flux, δ13C, and δ13D of CH4[J]. Global Biogeochemical Cycles, 11(3): 323-48.
UZAKI M, MIZUTANI H, WADA E. 1991. Carbon Isotope Composition of CH4from Rice Paddies in Japan [J]. Biogeochemistry, 13(2): 159-75.
VANDERGON H, NEUE H U. 1995. Influence Of Organic-Matter Incorporation On The Methane Emission From A Wetland Rice Field [J]. Global Biogeochemical Cycles, 9(1): 11-22.
VANDERGON H, VANBREEMEN N. 1993. Diffusion-controlled transport of methane from soil to atmosphere as mediated by rice plants. Biogeochemistry, 21: 177-90.
WANG B, NEUE H U, SAMONTE H P. 1997. Role of rice in mediating methane emission [J]. Plant Soil, 189(1): 107-15.
WHITICAR M J. 1999. Carbon and hydrogen isotope systematics of bacterial formation and oxidation of methane [J]. Chemical Geology, 161(1/3): 291-314.
YAGI K, MINAMI K. 1991. Emission and Production of Methane in the Paddy Fields of Japan [J]. Jarq-Japan Agricultural Research Quarterly, 25(3): 165-71.
ZHANG G B, JI Y, MA J, et al. 2012. Intermittent irrigation changes production, oxidation, and emission of CH4in paddy fields determined with stable carbon isotope technique [J]. Soil Biology and Biochemistry: 52(108-16).
ZHANG G B, JI Y, MA J, et al. 2013. Pathway of CH4production, fraction of CH4oxidized, and13C isotope fractionation in a straw-incorporated rice field [J]. Biogeosciences, 10(5): 3375-89.
ZHANG G B, ZHANG X Y, MA J, et al. 2011. Effect of drainage in the fallow season on reduction of CH4production and emission from permanently flooded rice fields [J].Nutrient Cycling in Agroecosystems, 89(1): 81-91.
曹亚澄, 孙国庆, 韩勇, 等. 2008. 大气浓度下N2O、CH4和CO2中氮、碳和氧稳定同位素比值的质谱测定[J]. 土壤学报,45(2): 249-258.
林而达. 1993. 减少农业甲烷排放的技术选择[J]. 农村生态环境, (S1): 9-12, 58.
林敏, 尤崇杓. 1989. 水稻根分泌物及其与粪产碱菌的相互作用[J]. 中国农业科学, (6): 6-12.
沈永平, 王国亚. 2013. IPCC第一工作组第五次评估报告对全球气候变化认知的最新科学要点[J]. 冰川冻土, 35(5): 1068-1076.
陶战, 杜道灯, 周毅, 等. 1994. 稻田施用沼渣对甲烷排放通量的影响[J].农村生态环境, (3): 1-5.
张晓艳, 张广斌, 纪洋, 等. 2010. 冬季淹水稻田CH4产生、氧化和排放规律及其影响因素研究[J]. 生态环境学报, 19(11): 2540-2545.
Effect of Rice Variety on Production and Emission of CH4and δ13CH4
ZHANG Wenxuan1,2, YU Haiyang1,2, ZHANG Guangbin1, MA Jing1, XU Hua1*
1. State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China; 2. Graduate University of Chinese Academy of Sciences, Beijing 100049, China
Rice variety is a key factor regulating CH4production and emission. However, little has so far been reported on effect of rice varieties on δ13C-value of CH4produced and emitted. Pot and indoor incubation experiments were carried out and with the aid of the stable carbon isotope method, CH4production potential of the paddy soil, CH4concentration in pore water, CH4production and emission flux as well as δ13C-value of the CH4emitted from the paddy soils cultivated with different varieties of rice (Zhendao 624, Nongxiang 98 and Zhongzao 33) under continuous flooding at four rice growing stages were studied and measured, with a view to providing some data in respect to CH4emission processes and δ13C-values of CH4emitted for reference in screening rice varieties for quality ones both high in yield and low in CH4emission. Results show that the paddy soils under Zhendao 624 and Nongxiang 98 were significantly higher than that under Zhongzao 33 at the tillering and booting stages in CH4production potential but significantly lower at the grain-filling and ripening stages. CH4production potentials, the highest CH4concentration in pore water and the lowest soil Eh of the three paddy soils under Zhendao 624, Nongxiang 98 and Zhongzao 33 appeared at the booting stage (2.6 μg·g-1·d-1, 346.9 μmol·L-1, -296 mV), booting stage (3.2 μg·g-1·d-1, 425.9 μmol·L-1, -316 mV) and grain-filling stage (2.4 μg·g-1·d-1, 435.2 μmol·L-1, -308 mV), respectively. Correlation analysis shows that CH4production potential of the paddy soil was significantly positively related to CH4concentration in pore water, but significantly negatively to soil Eh (P<0.01), regardless of rice varieties planted. CH4fluxes from the plots under Zhendao 624 and Nongxiang 98 peaked at the booting stage (67.1, 68.7 mg·m-2·h-1), while that from the plot under Zhongzao 33 did at the tillering stage (58.5 mg·m-2·h-1). The total CH4emission from the plots under Zhendao 624 and Nongxiang was 55.29 and 55.74 g·m-2, respectively, posing no obvious difference between the two, while that from the plot under Zhongzao 33 was 40.82 g·m-2(P=0.05), much lower than the former two, partly by reason that CH4production potentials of the former two were significantly higher than that of the latter at the tillering stage and booting stage, when CH4emissions occurred concentratedly in all the three. Correlation analysis also shows that CH4flux was significantly positively related to CH4production potential of the paddy soil at all the four stages (P<0.01), which fully demonstrates that rice plants different in variety affect CH4production and then CH4emission through regulating soil CH4production potential. δ13C-values of the CH4produced in the plots under Zhendao 624 and Zhongzao 33 gradually increased within the range of -67.0‰~-55.5‰, making no big difference between the two soils during the rice growing season, while that in the plot under Nongxiang 98 declined first and rose again within the range of -64.2‰~-52.9‰, which means that the soils in the plots under Zhendao 624 and Zhongzao 33 are quite similar in path way of CH4production, but sharply different from that under Nongxiang 98. The CH4emitted from the three plots all displayed a general trend in δ13C-value, declining first and rising again and varying in the range of -67.6‰~-48.5‰, -73.0‰~-47.3‰and -60.9‰~-46.7‰, respectively. To sum up, the effect of rice variety on δ13CH4varies with the season.
rice variety; CH4production; CH4emission; δ13C; soil Eh
10.16258/j.cnki.1674-5906.2015.02.003
X511
A
1674-5906(2015)02-0196-08
张文宣,于海洋,张广斌,马静,徐华. 水稻品种对CH4产生、排放及δ13CH4的影响[J]. 生态环境学报, 2015, 24(2): 196-203.
ZHANG Wenxuan, YU Haiyang, ZHANG Guangbin, MA Jing, XU Hua. Effect of Rice Variety on Production and Emission of CH4and δ13CH4[J]. Ecology and Environmental Sciences, 2015, 24(2): 196-203.
中国科学院战略性先导科技专项(XDB15020103);国家科技支撑计划项目(2013BAD11B02);国家自然科学基金项目(41271259;41201243);江苏省自然科学青年基金项目(BK2012497)
张文宣(1989年),女,硕士研究生,主要从事农田温室气体排放规律及其机理研究。E-mail: wxzhang@issas.ac.cn *通信作者:E-mail: hxu@issas.ac.cn
2014-11-23
