人M2型丙酮酸激酶促进肾癌细胞生长
2015-11-29梅国徽麦海星梁迎春张柯郭靖李玲董倩洪甜徐小洁叶棋浓陈立军
梅国徽 ,麦海星,梁迎春,张柯,郭靖,李玲,董倩,洪甜,徐小洁,叶棋浓,陈立军
军事医学科学院a.附属医院泌尿外科,北京 100071;b.生物工程研究所,北京 100850;c.科技部,北京 100850
丙酮酸激酶(pyruvate kinase,PK)是糖酵解途径的关键酶之一,可以催化磷酸烯醇式丙酮酸产生丙酮酸。PK 有4 种同工酶,即PKL、PKR、PKM1、PKM2,其中PKM2 主要表达于胚胎及增殖活跃的细胞中,随着胚胎的形成,在红细胞和肝脏中分别由PKR 和PKL 取代,骨骼肌、心脏和脑组织中则表达PKM1[1-5]。1972 年,Imamura 发现在肝癌细胞中PKL异常转变为PKM2[3]。随后的研究表明,在乳腺癌[6]、膀胱癌[7]、结肠癌[8]、肺癌[9]、肾透明细胞癌[10]等肿瘤中都伴有PKM2 的高表达,且与肿瘤的发展与转移密切相关。
我们拟构建PKM2 的真核表达载体,并验证其对肾癌细胞系Caki-1 生长的影响,为进一步研究PKM2与肾癌发生发展的关系奠定基础。
1 材料和方法
1.1 材料
人胚肾293T 细胞由本室传代培养;人透明细胞癌细胞Caki-1 由解放军总医院赠送;人乳腺文库、pXJ-40-myc 载体为本室保存;限制性内切酶、PCR试剂、DNA 连接酶购自TaKaRa 公司;质粒提取、胶回收试剂盒购自Promega 公司;VigoFect 购自威格拉斯生物技术有限公司;DMEM 培养基购自Gibco 公司;胎牛血清购自浙江天杭生物科技有限公司;引物由北京赛百盛生物技术有限公司合成;基因测序由北京博迈德科技发展有限公司完成。
1.2 myc-PKM2重组质粒的构建与测序
以人乳腺文库为模板,根据文献报道的PKM2序列合成上游引物(5'-CGGGATCCATGCCCCGGA GGGCGGAGAA-3')和下游引物(5'-CCGCTCGAGT CAATCTCCCATCCGTTGATGTG-3'),PCR 扩增人的PKM2编码序列。扩增条件:95℃预变性5 min;95℃变性30 s、58℃退火30 s、72℃延伸2 min,31个循环后;72℃延长7 min;最后用胶回收PCR产物。
用BamHⅠ和XhoⅠ双酶切pXJ-40-myc 载体,10 g/L 琼脂糖凝胶电泳后,胶回收载体的大片段;将PCR 片段回收后再用BamHⅠ和XhoⅠ酶切,形成带有粘性末端的双链,用T4DNA 连接酶连接入pXJ-40-myc 载体;连接产物转化大肠杆菌DH5α感受态后,挑选克隆进行菌液PCR,选取阳性克隆培养并提取质粒,用BamHⅠ和XhoⅠ双酶切鉴定,酶切鉴定阳性的克隆送北京博迈德科技发展有限公司测序。
1.3 细胞转染和Western印迹检测
用含双抗及10%胎牛血清的DMEM培养基将细胞接种于2块6 cm皿中,接种量以转染时60%~70%的细胞密度为宜,常规培养24 h 后行转染,转染前1 h 换液;将4 μL VigoFect 与200 μL NaCl 混合,在其静置5 min 的过程中,将10 μg 重组质粒与200 μL NaCl 混合,然后将上述2 种溶液混匀,室温孵育15 min 后滴入6 cm 皿中,以同种方法转染空pXJ-40-myc 载体作为对照;37℃、5% CO2条件下常规培养,转染4 h 后换液,24 h 后收集细胞,用1×PBS 溶液清洗1 次,4℃、3000 r/min 离心3 min 后弃上清,加入2 倍细胞量的RIPA 于冰上裂解30 min,加入等量2×SDS 上样缓冲液,煮沸15 min,12 000 r/min 离心3 min 后取上清液进行SDS-PAGE,湿转法转至硝酸纤维素膜上;用5%脱脂奶粉于常温封闭1 h,加入用5%脱脂奶粉以1∶2000 稀释的myc-HRP 鼠单克隆抗体,室温下轻摇1 h,用1×TBST 洗膜3 次,每次5 min;ECL 显色反应3 min,暗室压片显影。
1.4 生长曲线实验
pmyc-PKM2 质粒转染Caki-1 细胞24 h 后,与转染pXJ-40-myc 空载体的Caki-1 细胞同时以每孔3000 个细胞的密度接种于5 块96 孔板,每组设3 个复孔,分别在第0、1、2、3、4 d 加入100 μL 10%CCK-8后37℃孵育1 h,测定D450nm值,以培养时间为横轴、D450nm平均值为纵轴绘制生长曲线。
2 结果
2.1 myc-PKM2重组质粒的构建与鉴定
以本室保存的人乳腺文库为模板,PCR 扩增人PKM2 的编码序列,获得长约1500 bp 的DNA 片段,与预期大小一致(图1)。将BamHⅠ和XhoⅠ双酶切后的PCR 产物和pXJ-40-myc 载体连接,转化大肠杆菌DH5α感受态,挑克隆后进行菌液PCR 鉴定,若获得与目的条带1500 bp大小接近(图2)的克隆,则初步认为是带有人PKM2 基因的阳性重组克隆。将所得阳性克隆提质粒,经酶切鉴定,可切出2 条长度分别约为5000 和1500 bp 的条带,且相应的载体经酶切只见大片段,符合预期结果(图3)。DNA 序列测序结果表明,插入的DNA 序列与人PKM2 基因的编码序列完全一致(数据略)。
2.2 Western 印迹检测myc-PKM2 在293T 细胞中的表达
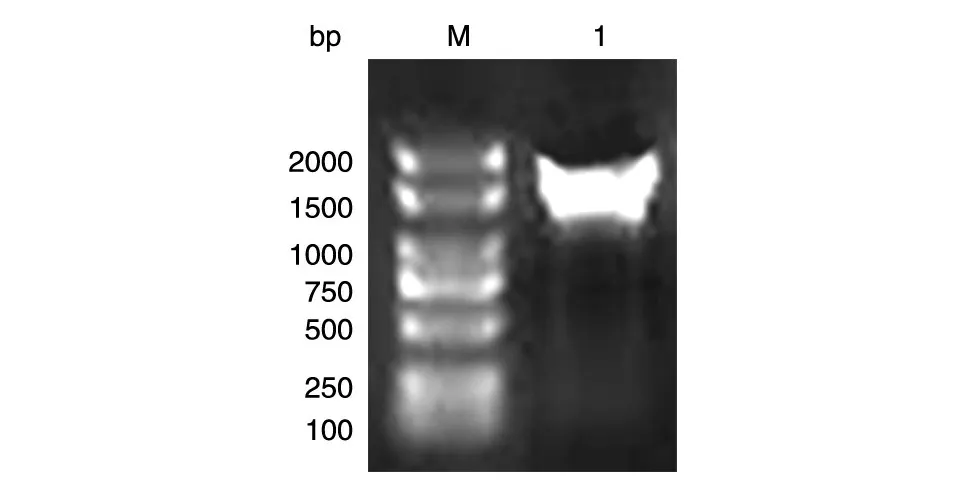
图1 PCR扩增人PKM2的编码序列
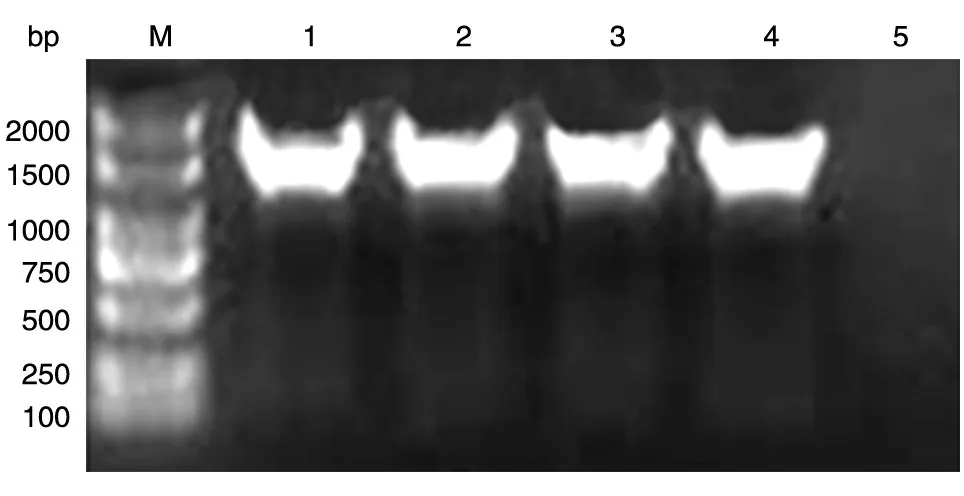
图2 重组质粒pmyc-PKM2的菌液PCR结果电泳图谱
将构建的pmyc-PKM2 重组质粒和空载体分别转染293T 细胞系,24 h 后提取蛋白进行SDSPAGE,Western 印迹检测myc-PKM2 的表达。结果显示,转染重组质粒后,用myc-HRP 抗体能在相对分子质量约60×103处检测到明显的特异性条带,而空载体无条带(图4)。
2.3 CCK8 法验证myc-PKM2 促进肿瘤细胞系的生长
将pmyc-PKM2真核表达载体及pXJ-40-myc 载体转染肾癌Caki-1 细胞,通过CCK8 法进行生长曲线实验。结果显示,myc-PKM2 能促进肾癌Caki-1细胞的生长(图5)。
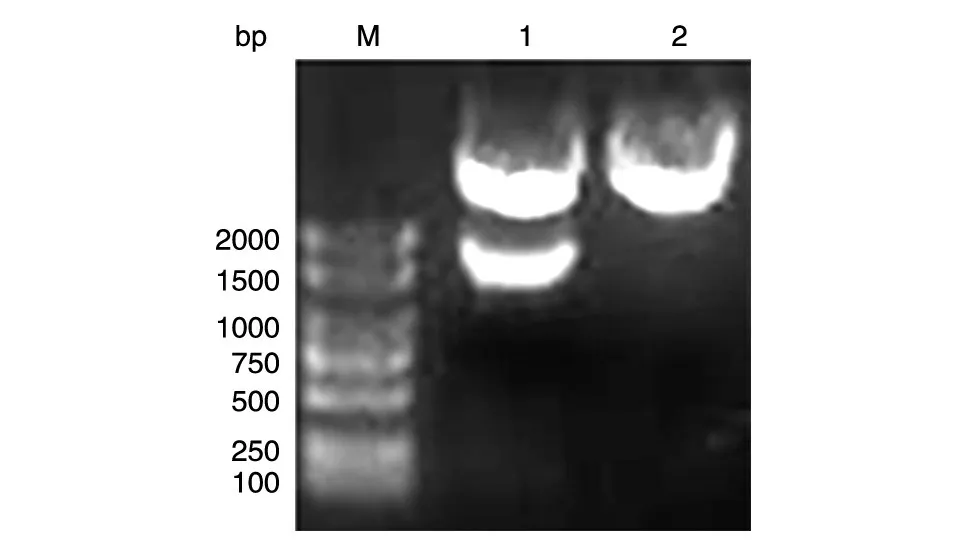
图3 重组质粒pmyc-PKM2的BamHⅠ/XhoⅠ双酶切电泳图谱
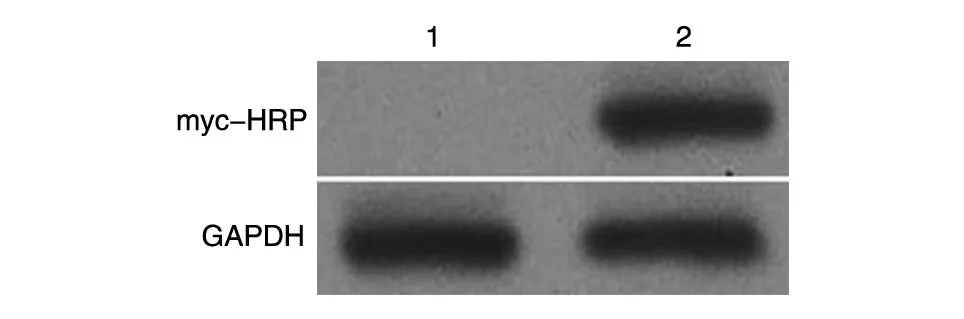
图4 Western印迹检测融合蛋白的表达
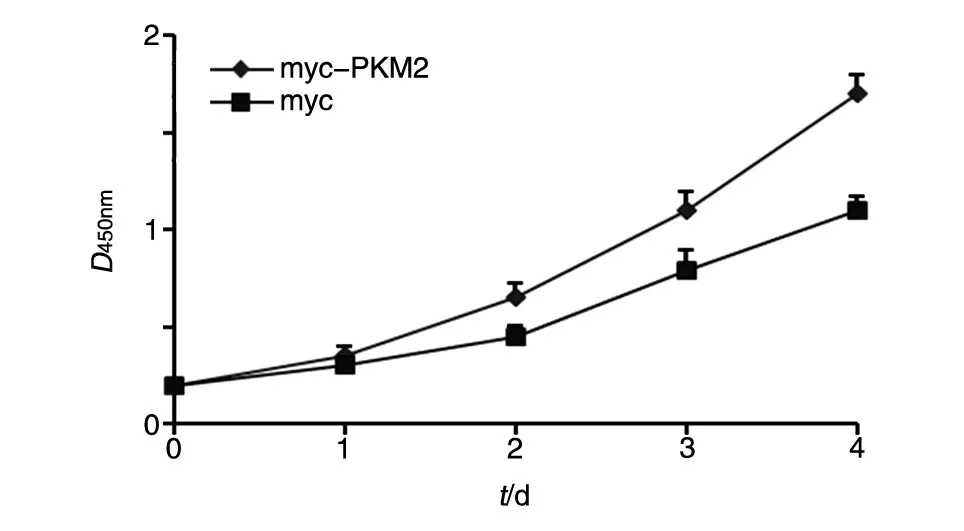
图5 PKM2促进Caki-1细胞生长
3 讨论
1924 年Warburg 观察到,即使在氧气充足的条件下,肿瘤细胞依然通过糖酵解途径获取能量,而非有氧呼吸,这一现象被称为Warburg 效应或有氧糖酵解,是肿瘤细胞区别于正常细胞的特征之一。近年来的研究表明,PKM2 可能通过激活Warburg 效应促进了肿瘤的发生[11-14]。
PKM2 是PK 的同工酶之一,由PKM 基因编码。PKM 基因前体mRNA 经由核不均一核糖核蛋白(hnRNP)A1/A2 和多嘧啶序列结合蛋白(PTB)剪切拼接后可以形成PKM1 和PKM2,其中PKM1 包含外显子9,PKM2则包含外显子10[15-16]。PKM2主要在胚胎时期表达,在成人组织中多被其他同工酶替代,肿瘤组织中多伴有PKM1 向PKM2 的转变[17-19]。与PKM1不同,PKM2的催化活性受外显子10编码的一个特殊结构域调控,当与1,6-二磷酸果糖(FBP)结合时,可引起PKM2 二聚体的变构、结合,形成四聚体结构,这种PKM2 四聚体对磷酸烯醇式丙酮酸(PEP)具有较强的亲和力和催化能力,可以促进糖酵解的进行;在不与FBP 结合时,PKM2 以二聚体形式存在,这种二聚体对PEP的催化能力较弱,从而引起糖酵解中间产物的累积,促进生物合成,支持细胞的生长和繁殖[20-21]。四聚体/二聚体的比例受一些代谢中间产物如FBP 的调节,PKM2 通过这一方式调控肿瘤细胞能量代谢与生物合成之间的平衡。近年发现PKM2 还具有调节基因表达、细胞周期进程的功能[22-23]。
PKM2 在多种肿瘤组织中过量表达,且在乳腺癌、结肠癌、泌尿系统肿瘤、胃肠道肿瘤患者的血清中可观测到PKM2 的升高。肾癌是泌尿系统常见的恶性肿瘤,Brinck通过免疫组化发现PKM2分布在正常肾脏的远曲小管和癌组织中,RT-PCR 显示癌组织中PK活性升高2~7倍[10]。Wechsel观察到,成功切除肾细胞癌后,血清PKM2 可在11 周内恢复到正常水平,而当肿瘤复发或转移时,血清PKM2 会再次升高,提示PKM2与肾癌关系密切[24]。
综上所述,我们构建的myc-PKM2 重组质粒在真核细胞中获得表达,且能够促进肾癌细胞系Caki-1 的生长。这为后续研究PKM2 在肾癌发生发展中的作用奠定了基础,有望为肾癌的诊断和靶向治疗提供新的思路。
[1]Noguchi T,Yamada K,Inoue H,et al.The L-and R-type isozymes of rat pyruvate kinase are produced from a single gene by use of different promoters[J].J Biol Chem,1987,262(29):14366-14371.
[2]Yamada K,Noguchi T.Nutrient and hormonal regulation of pyruvate kinase gene expression[J].Biochem J,1999,337(Pt 1):1-11.
[3]Imamura K,Tanaka T.Multimolecular forms of pyruvate kinase from rat another mammalian tissues I.Electrophoretic studies[J].J Biochem,1972,71(6):1043-1051.
[4]Chaneton B,Gottlieb E.Rocking cell metabolism:revised functions of the keyglycolytic regulator PKM2 in cancer[J].Trends Biochem Sci,2012,37(8):309-316.
[5]Muñoz M E,Ponce E.Pyruvate kinase:current status of regulatory and functional prop-erties[J].Comp Biochem Physiol B Biochem Mol Biol,2003,135(2):197-218.
[6]Lüftner D,Mesterharm J,Akrivakis C,et al.Tumor type M2 pyruvate kinase expression in advanced breast cancer[J].Anticancer Res,2000,20(6D):5077-5082.
[7]Li J,Yang Z,Zou Q,et al.PKM2 and ACVR1 care prognostic markers for poor pro-gnosis of gallbladder cancer[J].Clin Transl Oncol,2014,16(2):200-207.
[8]Christofk H R,Vander Heiden M G,Harris M H,et al.The M2 splice isoform of pyruv-ate kinase is important for cancer metabolism and tumor growth[J].Nature,2008,452(7184):230-233.
[9]Schneider J,Neu K,Grimm H,et al.Tumor M2-pyruvate kinase in lung cancer patients:immunohistochemical detection and disease monitoring[J].Anticancer Res,2002,22(1A):311-318.
[10]Brinck U,Eigenbrodt E,Oehmke M,et al.L-and M2-pyruvate kinase expression in renal cell carcinomas and their metastases[J].Virchows Arch,1994,424(2):177-185.
[11]Christofk H R,Vander Heiden M G,Harris M H,et al.The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumor growth[J].Nature,2008,452(7184):230-233.
[12]Vander Heiden M G,Christofk H R,Schuman E,et al.Identification of small molecule inhibitors of pyruvate kinase M2[J].Biochem Pharmacol,2010.79(8):1118-1124.
[13]Anastasiou D,Yu Y,Israelsen W J,et al.Pyruvate kinase M2 activators promote tetr-amer formation and suppress tumorigenesis[J].Nat Chem Biol,2012,8(10):839-847.
[14]Yang W,Lu Z.Nuclear PKM2 regulates the Warburgeffect[J].Cell Cycle,2013,12(19):3154-3158.
[15]David C J,Chen M,Assanah M,et al.HnRNP proteins con-trolled by c-Myc deregul-ate pyruvate kinase mRNA splicing in cancer[J].Nature,2010,463(7279):364-368.
[16]Sun Q,Chen X,Ma J,et al.Mammalian target of rapamycin up-regulation of pyruv-ate kinase isoenzyme type M2 is critical for aerobic glycolysis and tumor growth[J].Proc Natl Acad Sci USA,2011,108(10):4129-4134.
[17]Ashizawa K,Willingham M C,Liang C M,et al.In vivo regulation of monomer-tetramer conversion of pyruvate kinase subtype M2 by glucose is mediated via fructose1,6-bisphosphate[J].J Biol Chem,1991,266(25):16842-16846.
[18]Lv L,Xu Y P,Zhao D,et al.Mitogenic and oncogenic stimulation of K433 acetylation promotes PKM2 protein kinase activity and nuclear localization[J].Mol Cell,2013,52(3):340-352.
[19]Chiavarina B,Whitaker-Menezes D,Martinez-Outschoorn U E,et al.Pyruvate kinase expression(PKM1 and PKM2) in cancer-associated fibroblasts drives stromal nutrient production and tumor growth[J].Cancer Biol Ther,2011,12(12):1101-1113.
[20]Duvel K,Yecies J L,Menon S,et al.Activation of a metabolic gene regulatory network downstream of mTOR complex 1[J].Mol Cell,2010,39(2):171-183.
[21]Eigenbrodt E,Leib S,Kramer W,et al.Structural and kinetic differences between th-e M2 type pyruvate kinases from lung and various tumors[J].Biomed Biochim Acta,1983,42(11-12):S278-282.
[22]Gao X,Wang H,Yang J J,et al.Pyruvate kinase M2 regulates gene transcription by acting as a protein kinase[J].Mol Cell,2012,45(5):598-609.
[23]Jiang Y,Li X,Yang W,et al.PKM2 regulates chromosome segregation and mitosis progression of tumor cells[J].Mol Cell,2014,53(1):75-87.
[24]Wechsel H W,Petri E,Bichler K H,et al.Marker for renal cell carcinoma(RCC):thedimeric form of pyruvate kinase type M2(Tu M2-PK)[J].Anticancer Res,1999,19(4A):2583-2590.
