辽宁盘锦稻区杂草地与稻田中蜘蛛动态
2015-11-25马晓慧桑海旭
马晓慧, 桑海旭, 刘 郁
(辽宁省盐碱地利用研究所, 盘锦 124010)
辽宁盘锦稻区杂草地与稻田中蜘蛛动态
马晓慧*, 桑海旭, 刘 郁
(辽宁省盐碱地利用研究所, 盘锦 124010)
采用吸虫器法,系统调查了辽宁盘锦稻区杂草地与稻田中蜘蛛种类及优势度情况,以及蜘蛛优势种、种类数和个体数的年度变化趋势。结果表明,杂草地全年共有4个优势种,为草间钻头蛛、横纹金蛛、三突花蛛和黄褐新圆蛛。水稻生长期内稻田共有3个优势种,为草间钻头蛛、横纹金蛛、粽管巢蛛。杂草地与稻田中优势蜘蛛数量与蜘蛛个体数量变化趋势大体一致,均为缓慢上升趋势,最高峰均在8月中下旬,分别达到139头/m2和155头/m2,至水稻成熟期迅速减少。水稻插秧前与收割后,杂草地蜘蛛种类数最高值分别为23种/10 m2和34种/10 m2,均高于稻田;稻田中蜘蛛种类为缓慢增加,至收割前达到峰值23种/10 m2。两个生境蜘蛛种群的相似性系数为0.666 7。
杂草地生境; 稻田; 蜘蛛; 优势种; 动态
非稻田生境及其植被中的天敌是稻田天敌种群建立或重建的重要来源[1-4]。杂草是稻田周边最常见的非作物生境,其对稻田生境内的生物具有一定的影响。杂草改变了农作物生境的特征,为捕食性节肢动物的猎物提供替代寄主植物等[5]。
蜘蛛是稻田内仅次于昆虫的种类多、数量大的节肢动物类群,全部为捕食性,也是水稻害虫的重要天敌之一。盘锦稻区是辽宁省水稻主产区,蜘蛛对控制稻田生境内水稻害虫起着重要的作用。然而,北方稻区关于稻田和杂草内蜘蛛种群特征的调查分析鲜有报道。本文通过调查两个生境内蜘蛛资源,初步分析其种群动态特征,探明蜘蛛种群在两个生境中的发生特点及其地位,为后续蜘蛛研究提供依据。
1 材料和方法
1.1 调查地概况
试验在辽宁省盐碱地利用研究所新建基地完成,杂草地与稻田地相邻,南北并列,东西长度均为40 m。北面为杂草地,面积600 m2,历年植保资料显示,盘锦地区杂草种类40~50余种,稗草、双穗雀稗、鸭舌草、苦草等10余种为优势杂草,高度以上层和中层为主,由于常年施用适量的除草剂控制,杂草丰富程度不高,其北面毗邻灌溉水渠;南地是面积为800 m2的试验稻田。两地块受人为干扰较轻,调查时间为2013年5-10月。水稻品种采用本地主栽品种‘盐丰47’。
1.2 调查方法
杂草地采取随机平行线跳跃式5点取样,每样方面积为1 m2,样方间距10 m,样方距稻田1.5 m以上[6]。稻田采取对角线5点取样法,每个样方选1 m2左右,约22穴(水稻成熟期株数约为350株)。蜘蛛样本采用手提式吸虫器抽吸。采集的蜘蛛样本参照陈孝恩等人方法,经室内清样、鉴定,统计蜘蛛种类数、每种个体数[7-12]。
盘锦稻区为单季稻,水稻插秧一般在5月26日前后,2013年基地水稻插秧时间为5月27日,6月10日起首次调查,每月的10日、25日调查一次,直至10月2日水稻收割,共计调查8次。杂草地在水稻生长期与水稻同步调查,但插秧前5月10日、25日各调查1次,水稻收割后10月10日、25日各调查1次,共计12次。
1.3 数据统计与分析
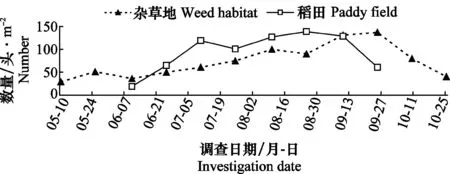
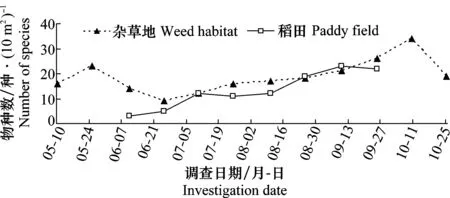
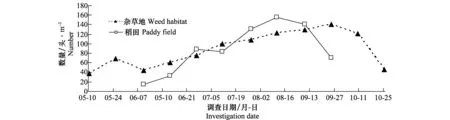
用物种丰富度S、个体数量N、Berger-Parker的优势度指数D对调查数据进行分析。按优势度D判定蜘蛛各种类的丰盛程度:D≥0.1时定义该物种为优势种、0.05≤D<0.1定义为丰富种、0.01 2.1 稻田和杂草地蜘蛛物种组成及优势度对比 通过系统取样调查,水稻插秧前,杂草地共采集到23种蜘蛛,其中优势种3种,丰富种2种,常见种5种。水稻生长期内,杂草地共采集到44种蜘蛛,其中优势种2种,丰富种5种,常见种10种;稻田共采集到49种蜘蛛,其中优势种3种,丰富种3种,常见种10种。水稻收割后,杂草地共采集到蜘蛛35种,其中优势种2种,丰富种4种,常见种达14种(表1)。 表1 杂草地和稻田中主要蜘蛛种群组成及优势度比较1) 1) Do优势种;A丰富种;C常见种;优势度指数D≥0.1、0.05≤D<0.1和0.01 Do=Dominant species; A=Abundant species; C=Common species;D≥0.1, 0.05≤D<0.1 and 0.01 2.2 稻田和杂草地蜘蛛种群动态变化对比 2.2.1 蜘蛛优势种数量的季节动态 通过对杂草地12次调查和稻田8次调查,发现杂草地蜘蛛种群共有4种优势种,分别为草间钻头蛛、三突花蛛、黄褐新圆蛛和横纹金蛛,它们在不同调查时期各自数量变化均不同,其中黄褐新圆蛛在水稻插秧前为优势种,到水稻生长期开始演变为丰富种;三突花蛛在水稻收割后为丰富种;横纹金蛛仅在水稻收割后成为杂草地的优势种。稻田蜘蛛种群共有3个优势种,分别为草间钻头蛛、横纹金蛛和粽管巢蛛。 优势蜘蛛的总量在全年调查期呈现一定的规律性(图1)。杂草地优势蜘蛛总量9月底之前呈逐渐上升的趋势,8月上旬出现第一个数量高峰100头/m2,到8月中下旬稍有减少,但9月上旬迅速增多至137头/m2。稻田优势蜘蛛全年出现两个高峰,第一个高峰在7月上中旬为119头/m2,第二个高峰在8月中下旬为139头/m2,蜘蛛数量整体为上升趋势直至水稻接近收割,数量才开始减少。可见,同一时期不同优势种在种群中的数量比例不同,同一优势种不同时期在种群中的数量比例也是不相同的。 图1 杂草地与稻田蜘蛛优势种的数量动态(盘锦,2013)Fig.1 Population dynamic of dominant spiders in weed habitat and paddy field (Panjin,2013) 2.2.2 蜘蛛物种数的季节动态 5月10日调查杂草地蜘蛛为17种/10 m2,5月25日达到23种/10 m2,水稻插秧后杂草地蜘蛛种类明显减少,继而种类再缓慢增加,至水稻收割后迅速增加,达到峰值34种/10 m2。稻田蜘蛛在插秧2周时,种类为3种/10 m2,然后逐渐增加,在9月上中旬达到最大值23种/10 m2。这些变化可能与蜘蛛在杂草地和稻田间的来回迁移有关(图2)。 图2 杂草地与稻田蜘蛛物种数的动态(盘锦,2013)Fig.2 Dynamics of species abundance of spiders in weed habitat and paddy field (Panjin,2013) 2.2.3 蜘蛛总数量动态 水稻插秧前,杂草地蜘蛛数量从37头/m2增加至69头/m2,水稻插秧后,杂草地蜘蛛总量减少至44头/m2,随后蜘蛛在杂草地中的数量逐渐增加,至9月上中旬为140头/m2,10月数量迅速减少。稻田蜘蛛在刚开始调查时14头/m2,逐渐增加至88头/m2,7月下旬小幅度减少后继续增加,至8月底为155头/m2,近收割时蜘蛛总数量迅速减少。杂草地与稻田中蜘蛛个体数量与优势蜘蛛数量变化趋势大体一致,均为缓慢上升趋势,至水稻成熟期迅速减少。见图3。 图3 杂草地与稻田蜘蛛总数量动态(盘锦,2013)Fig.3 Dynamic of all spiders in weed habitat and paddy field (Panjin,2013) 2.2.4 蜘蛛种群相似性分析 盘锦稻区水稻生长期集中在6月至9月,分别调查杂草地及稻田内主要蜘蛛物种组成情况(表1),杂草地主要蜘蛛物种17种,稻田主要蜘蛛物种16种,含共有物种11种,因此,杂草地与稻田蜘蛛种群相似性系数计算为:Cs=2J/(a+b)=2×11/(17+16)=0.666 7,大于0.5,相似程度较高。 盘锦稻区为单季稻,水稻生长期集中在6月至9月,在形成的稻田生态系统中,天敌生物在水稻病虫害防控方面发挥着不可替代的作用。蜘蛛作为稻田生境内捕食性节肢动物群落中最主要的一大类群,其整个种群世代的发生发展一般不会只在一个生境内完成,在稻田生境建立前和消失后,必然有第二生境(或主要生境)完成整个世代,杂草地生境便可以认为是主要的第二生境(或主要生境)。 农业景观结构的多样性强烈地影响着发生在作物地中的天敌昆虫的物种丰富度和效能,理解这些景观结构是如何影响作物、害虫、天敌间的相互作用,是利用昆虫进行生物防治成功与否的关键[14]。杂草地的存在增加了农田系统生态多样性,且其中包含的多种生物,尤其是捕食性节肢动物,直接影响着稻田生境内的生物群落结构与状态。在研究中我们发现稻田生境内蜘蛛种群的建立离不开其周边杂草地,杂草地中的蜘蛛类群随着水稻插秧,逐渐地迁移至稻田生境,在稻田生境消失(水稻收割)前后,又迁回至杂草地生境内。然而,两个生境内蜘蛛种群的迁移规律并不是完全对应的。分析其原因,首先,杂草地在水稻生长初期迁入稻田的蜘蛛类群仅为一部分,可能是部分种类,也可能是部分数量,其余一定种类数量的蜘蛛仍然生活在杂草地生境中;其次,随着水稻生长,两个生境内的蜘蛛均繁殖后代,且因食源逐渐充足,气候等因素的逐渐适宜,各类群蜘蛛均呈现扩繁的趋势。这说明,蜘蛛类群在由杂草地转入稻田后,杂草地中蜘蛛类群并不一定减少,这与刘雨芳[6]研究的结果相一致。这些蜘蛛也会在稻田与周边草地之间来回迁移。 另外,研究中发现,蜘蛛在稻田生境内的发生趋势同稻飞虱发生趋势具有一定的相关性,盘锦稻区稻飞虱发生有两个高峰期,而蜘蛛发生高峰期稍滞后于稻飞虱。稻飞虱是蜘蛛的主要食源之一,推测食源昆虫的发生直接影响了蜘蛛种群的发展变化。两者的对应关系及影响因子等还有待后续试验继续研究。 群落的相似性系数是相似性的表现值,其大小直接表明两个群落物种组成的相似程度。本文采用相似性计算方法,研究了杂草地生境和稻田生境内两个蜘蛛种群的相似性程度,及各生境蜘蛛物种组成的相似情况,结果也表明了两个生境内蜘蛛种群的相似程度很高,且有很多的共有种,这也进一步说明了杂草地生境对稻田蜘蛛群落的形成具有一定的影响。因此,非作物生境的存在直接或间接影响着稻田生境内很多物种的发生及其群落的形成,而非作物生境在面积、相对位置、人为干扰等因素的共同作用下,其对稻田系统内生物的具体作用还需进一步的研究。 [1] Liss W J, Gut L J, Westigard P H.Perspectives on arthropod community structure, organization and development in agricultural crops [J].Annual Review of Entomology,1986,31: 455-478. [2] 张古忍,古德祥,张文庆. 稻田捕食性节肢动物群落的种库与群落的重建[J].中国生物防治,1997,13(2):65-68. [3] 古德祥,张古忍,张文庆. 稻田蜘蛛群落的重建及与其种库的相关性[J].蛛形学报,1999,8(2):89-94. [4] 刘雨芳. 稻田生态系统节肢动物群落结构研究[M].广州:中山大学,2000. [5] Norris R F,Kogan M. Interactions between weeds, arthropod pests, and their natural enemies in managed ecosystems [J].Weed Science,2000,48(1):94-158. [6] 刘雨芳,古德祥,张古忍.广东双季稻区杂草地和稻田中捕食性节肢动物的群落动态[J].昆虫学报,2003,46(5):591-597. [7] 陈孝恩,高君川. 四川农田蜘蛛彩色图册[M].成都:四川科学技术出版社, 1990:41-195. [8] 李永禧,周玉宏,王助引,等. 广西经济昆虫图册(捕食性昆虫)[M].南宁:广西科学技术出版社,1990:1-95. [9] 彭贤锦,谢莉萍,肖小芹,等. 中国跳蛛(蛛形纲:蜘蛛目)[M].长沙:湖南师范大学出版社,1993:21-250. [10]尹长民,彭贤锦,谢莉萍,等, 中国狼蛛(蛛形纲:蜘蛛目)[M].长沙:湖南师范大学出版社, 1997:23-289. [11]宋大祥,朱明生.中国动物志(蛛形纲:蜘蛛目:蟹蛛科,逍遥蛛科)[M].北京:科学出版社, 1997:136-149. [12]朱明生.中国动物志(蛛形纲:蜘蛛目:球蛛科)[M].北京:科学出版社, 1998:69-73. [13]丁岩软.昆虫数学生态学[M].北京:科学出版社,1994:438-451. [14]Paul C, Marino D, Landis A.Parasitoid community structure; implications for biological control in agricultural landscapes [M]∥Ekbom B S,Irwin M E, Robert Y, et al. Interchanges of insects between agricultural and surrounding landscapes. Kluwer Academic Publishers,2000:183-193. (责任编辑:王 音) Fluctuation of spiders in paddy field and adjacent weed habitat in Panjin, Liaoning Province, China Ma Xiaohui, Sang Haixu, Liu Yu (Liaoning Saline or Alkaline Land Utilization and Research Institute, Panjin 124010, China) The annual seasonal fluctuation of spider species abundance and dominance in paddy field and adjacent habitat was investigated with the suction sampler. Throughout the year, there were 4 dominant spider species in weed habitat, such asHylyphantesgraminicola,Argiopebruennichi,MisumenopstricuspidatusandNeosconadoenitzi, while there were 3 dominant spider species in paddy field, such asH.graminicola,A.bruennichiandClubionajaponicola. The seasonal fluctuation pattern of dominant spiders and density in these two habitats were similar, with the peak value of 139 individual/m2and 155 individual /m2, respectively. The abundance of spider species in weed habitat was more than that in the paddy field, and the peak abundance of spider species pre-and post-planting of rice was 23 species/10 m2and 34 species/10 m2, but 23 species/10 m2in paddy field in rice growing season. The similarity coefficient of spider populations is 0.666 7 on these two habitats. weed habitat; paddy field; spider; dominant species; dynamic 2014-04-21 2014-07-18 辽宁省科研事业公益研究基金项目(2011005010) S 476 A 10.3969/j.issn.0529-1542.2015.03.032 * 通信作者 E-mail: jodiexiaohui@163.com2 结果与分析

3 讨论
