Liver expression of Nrf2-related genes in different liver diseases
2015-11-21MingLiangChengYuanFuLuHongChenZhongYangShenandJieLiu
Ming-Liang Cheng, Yuan-Fu Lu, Hong Chen, Zhong-Yang Shen and Jie Liu
Guiyang, China
Liver expression of Nrf2-related genes in different liver diseases
Ming-Liang Cheng, Yuan-Fu Lu, Hong Chen, Zhong-Yang Shen and Jie Liu
Guiyang, China
BACKGROUND: The KEAP1-Nrf2 antioxidant signaling pathway is important in protecting liver from various insults. However, little is known about the expression of Nrf2-related genes in human liver in different diseases.
METHODS: This study utilized normal donor liver tissues (n=35), samples from patients with hepatocellular carcinoma (HCC, n=24), HBV-related cirrhosis (n=27), alcoholic cirrhosis (n=5) and end-stage liver disease (n=13). All of the liver tissues were from the Oriental Liver Transplant Center, Beijing, China. The expressions of Nrf2 and Nrf2-related genes, including its negative regulator Kelch-like ECH-associated protein 1 (KEAP1), its targeted gene NAD(P)H-quinone oxidoreductase 1 (NQO1), glutamate-cysteine ligase catalytic subunit (GCLC) and modified subunit (GCLM), heme oxygenase 1 (HO-1) and peroxiredoxin-1 (PRDX1) were evaluated.
RESULTS: The expression of Nrf2 was decreased in HCC, increased in alcoholic cirrhosis and end-stage liver disease. The expression of KEAP1 was increased in all of the liver samples. The most notable finding was the increased expression of NQO1 in HCC (18-fold), alcoholic cirrhosis (6-fold), endstage liver disease (5-fold) and HBV-related cirrhosis (3-fold). Peri-HCC also had 4-fold higher NQO1 mRNA as compared to the normal livers. GCLC mRNA levels were lower only in HCC, as compared to the normal livers and peri-HCC tissues. GCLM mRNA levels were higher in HBV-related cirrhosis and end-stage liver disease. HO-1 mRNA levels were increased in all liver tissues except for HCC. Peri-HCC had higher PRDX1 mRNA levels compared with HCC and normal livers. CONCLUSION: Nrf2 and Nrf2-related genes are aberrantly expressed in the liver in different diseases and the increase of NQO1 was the most notable finding, especially in HCC.
(Hepatobiliary Pancreat Dis Int 2015;14:485-491)
Nrf2-related gene expression;
cirrhosis;
hepatocellular carcinoma;
end-stage liver disease
Introduction
Nuclear factor-erythroid 2 related factor 2 (Nrf2) plays an important role in mediating cellular antioxidation that rescues the liver from a wide variety of insults.[1,2]Accumulating evidence suggests that the Nrf2-ARE (antioxidant responsive element) pathway exhibits diverse biological functions against viral hepatitis, alcoholic and nonalcoholic liver diseases, fibrosis, cirrhosis and hepatocellular carcinoma,[3,4]and is proposed to be a novel therapeutic target for liver fibrosis.[5]
The transcription factor Nrf2 is bound to Kelch-like ECH-associated protein 1 (KEAP1) in the cytosol under physiological conditions.[6]In response to toxic insults, the Nrf2-KEAP1 interaction is disrupted, and Nrf2 is translocated into the nucleus. Once in the nucleus, Nrf2 complexes with small musculo-aponeurotic fibrosarcoma proteins and binds to the ARE, promoting transcription of a large battery of cytoprotective genes.[7,8]Nrf2-ARE-targeted genes include the NAD(P)H-quinone oxidoreductase 1 (NQO1), glutamate-cysteine ligase catalytic and modified subunits (GCLC and GCLM), heme oxygenase 1 (HO-1), peroxiredoxin (PRDX1) and many others.[7-9]
Significant progress has been made to illustrate the role of Nrf2 in liver injury and liver diseases in animal models; however, little is known about Nrf2 and Nrf2-related gene expressions in human liver diseases. Through the Liver Tissue Cell Distribution System, Hardwick and coworkers[10]analyzed normal, steatotic and nonalcoholic steatohepatitis (fatty and non-fatty) tissues forNrf2 and Nrf2-targeted genes/proteins. The Nrf2 stain was evident in all stages of nonalcoholic fatty liver disease (NAFLD). NQO1 tended to increase with disease progression, while the mRNA of glutathione S-transferase (GST) isoforms showed an increased trend, total GST activity decreased with NAFLD progression. Glutathione (GSH) synthesis did not seem to be significantly dysregulated, and malondialdehyde concentrations increased with disease progression.[10]In biopsy samples from 13 primary biliary cirrhotic patients receiving ursodeoxycholic acid treatment, Nrf2 and Nrf2-mediated antioxidative proteins (thioredoxin and thioredoxin reductase-1) increased.[11]Takahashi et al[12]compared non-alcoholic steatohepatitis tissues and normal liver tissues, and found that non-alcoholic steatohepatitis livers had higher Nrf2 expression and its target genes such as γ-glutamylcysteine synthetase (γGCS), glutathione peroxidase 2 (GPx2), thioredoxin (TRX) and HO-1 were also overexpressed. However, in 24 "end-stage" livers, the expression of Nrf2 and Nrf2 targeted gene GCLC and GSTA1 were decreased, whereas HO-1 and NQO1 were not significantly altered, as compared to HCC-surrounding tissues from 6 patients.[13]These discrepancies are likely due to limited sample size and different stages of liver diseases. There is no data available on Nrf2 expression in livers of the Chinese population.
The present study evaluated the Nrf2 and Nrf2-related genes such as KEAP1, NQO1, GCLC, GCLM, HO-1 and PRDX1 in 93 samples from 69 patients with different liver diseases; each HCC patient contributed 2 samples, HCC and HCC-surrounding tissues. Thirty-five normal donor livers were used as controls.
Methods
Demographics
Informed consent was obtained from each patient. A total of 128 liver samples were collected from 104 patients in the Oriental Liver Transplant Center (Beijing, China). The samples are shown in Table 1 including the gender, age and diseases which we have recently published.[14]This study was approved by the Institutional Review Board of the General Hospital of Chinese People's Armed Police Forces and Guiyang Medical College of Human Subject Committee.
Total RNA isolation
The 128 human liver samples were used for real-time RT-PCR analysis. Total RNA was extracted with TRIzol (Invitrogen, Carlsbad, CA), purified with RNeasy columns (Qiagen, Valencia, CA). The quality and integrity of purified RNA was determined by spectrophotometry with 260/280 ratio>1.8 and by agarose gel electrophoresis showing clear 18S and 28S bands without degradation.
Real-time RT-PCR analysis
Total RNA was reverse-transcribed using a High Capacity RT kits from Applied Biosystems (Foster City, CA), and amplified with Power SYBR® Green PCR Master Mix in a 7900HT PCR System (Applied Biosystems, Foster City, CA). Oligonucleotide primers were designed with Primer 3 software (version 4), and are listed in Table 2. The standard real-time PCR (95 ℃ 15 seconds, and annealing and elongation at 60 ℃ for 60 seconds for 40 cycles) was used for cDNA amplification. Relative expression of genes was calculated by the 2-ΔΔCtmethod[2,14-17]and normalized to the housekeeping gene GAPDH.
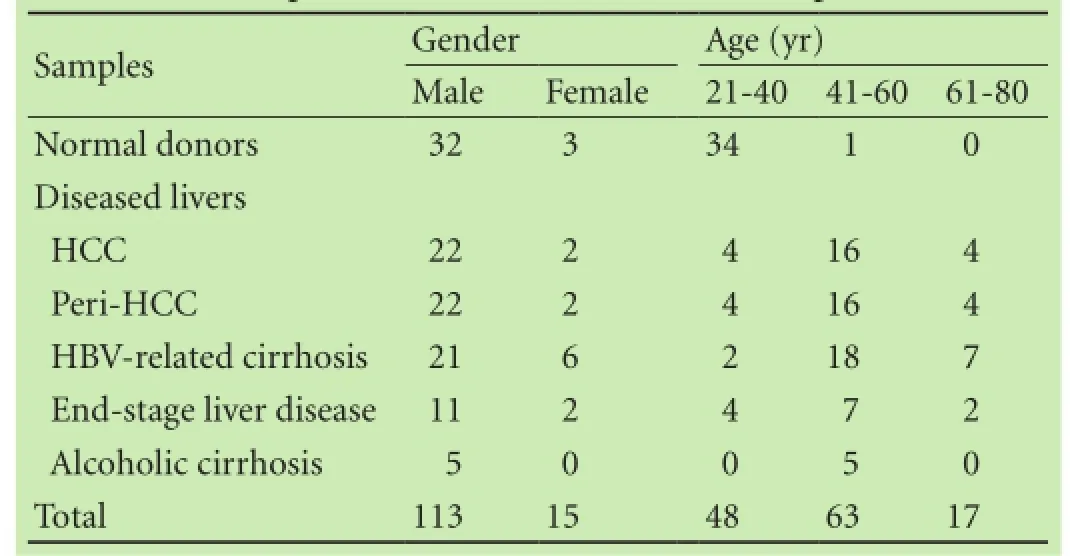
Table 1. Sample characteristics (128 from 104 patients)[14]

Table 2. Primer sequence for real-time RT-PCR analysis
Statistical analysis
Data were presented as mean±SEM. The SPSS17 software was used for statistical analysis. For comparisons among three or more groups, data were analyzed using one-way analysis of variance (ANOVA), followed by Duncan's multiple range test. The significant level was set at P<0.05 in all cases.
Results
Relative expression levels
Relative transcript levels of the Nrf2-related genes in normal Chinese human livers (n=35) and diseased liver samples (n=93) were significantly different, similar to that of P450 genes.[14]In general, diseased livers had higher variation in expression than normal livers. For example, in 35 normal livers, the individual variation of NQO1 was less than 10-fold, averaged 1.07%±0.05% of GAPDH; while in 93 diseased liver samples, the variations increased to 88-fold, with a 7-fold increase in average (7.98%±1.13% of GAPDH). Another example was HO-1, in 35 normal livers, the individual variation of HO-1 was less than 15-fold, averaged 17.40%±3.41% of GAPDH; while in 93 diseased liver samples, the variations increased to 32-fold, with a 2.4-fold increase in average (41.10%±2.96% of GAPDH). The expression of GCLC and PRDX1 was relatively stable. Although the average expression levels of Nrf2 remained the same, the variations increased from 5-fold in 35 normal livers to 30-fold in 93 diseased liver samples. Thus, Nrf2-related gene expressions were prone to change under liver pathology conditions (Table 3).
Nrf2 and KEAP1 expression
The expression of Nrf2 is shown in Fig. 1. There was no significant difference in Nrf2 mRNA levels between normal livers and peri-HCC tissues. Compared to peri-HCC, Nrf2 expression in HCC was decreased by 60% (38.7% vs 16.9% of GAPDH). Compared to normal livers (35.4% of GAPDH), Nrf2 mRNA levels were higher in alcoholic cirrhosis (59.4% of GAPDH) and end-stage liver disease (55.4% of GAPDH).
The expression of KEAP1 is shown in Fig. 1B. KEAP1 is a negative regulator of Nrf2,[6]and its expression in normal livers averaged 6.0% of housekeeping gene GAPDH. KEAP1 mRNA levels were increased by 5-12 fold in the different diseased livers, ranging from 28% of GAPDH in HBV-related cirrhosis to 58% of GAPDH in alcoholic cirrhosis. KEAP1 mRNA expression was independent of Nrf2 mRNA levels. For example, HCC had a decreased expression of Nrf2 and an increased expression of KEAP1; cirrhotic livers had increased KEAP1 levels but also had increased expression of Nrf2.
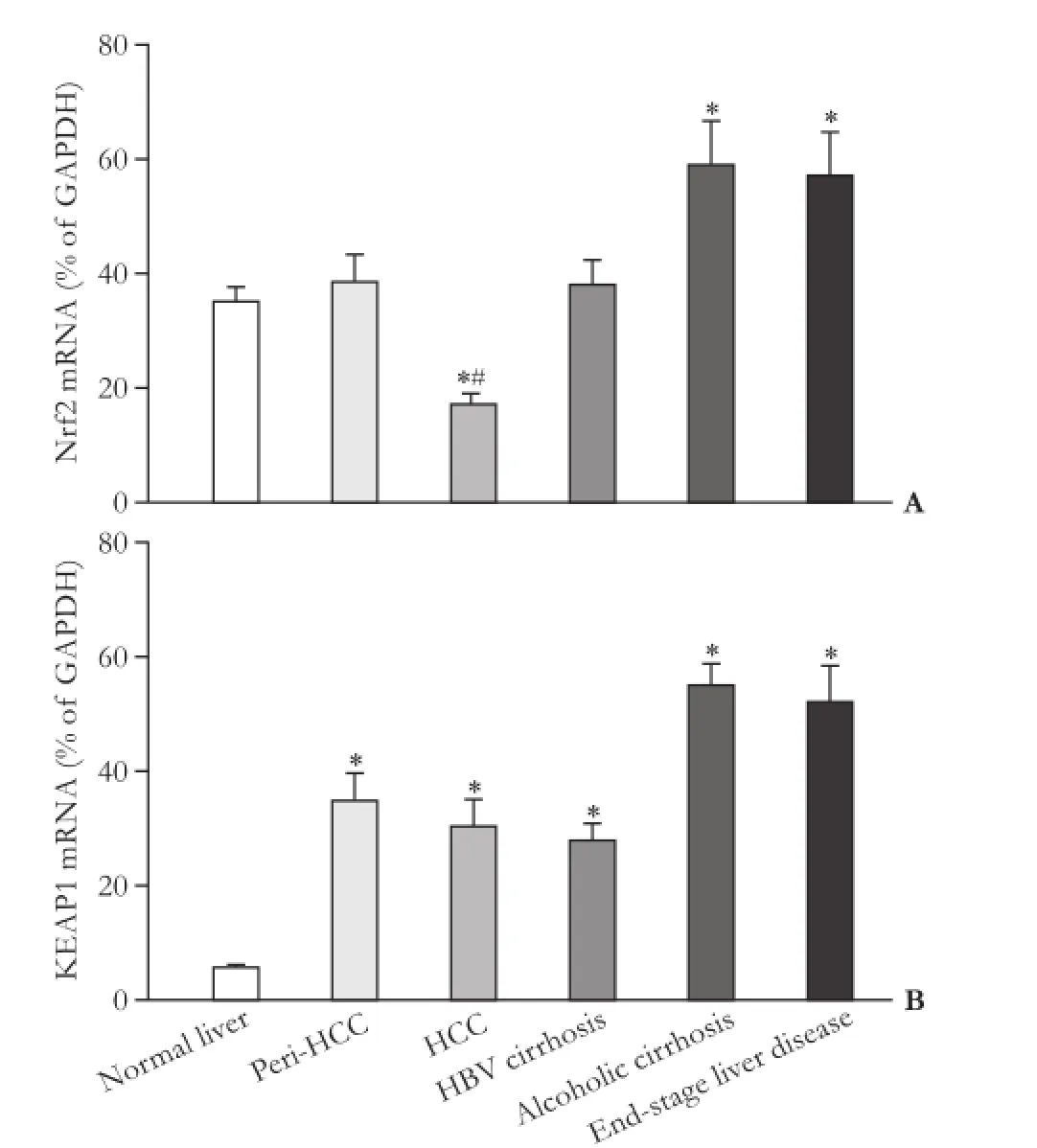
Fig. 1. Expression of Nrf2 and KEAP1 in human livers. *: significantly different from normal liver, P<0.05; #: significantly different from peri-HCC tissue, P<0.05.
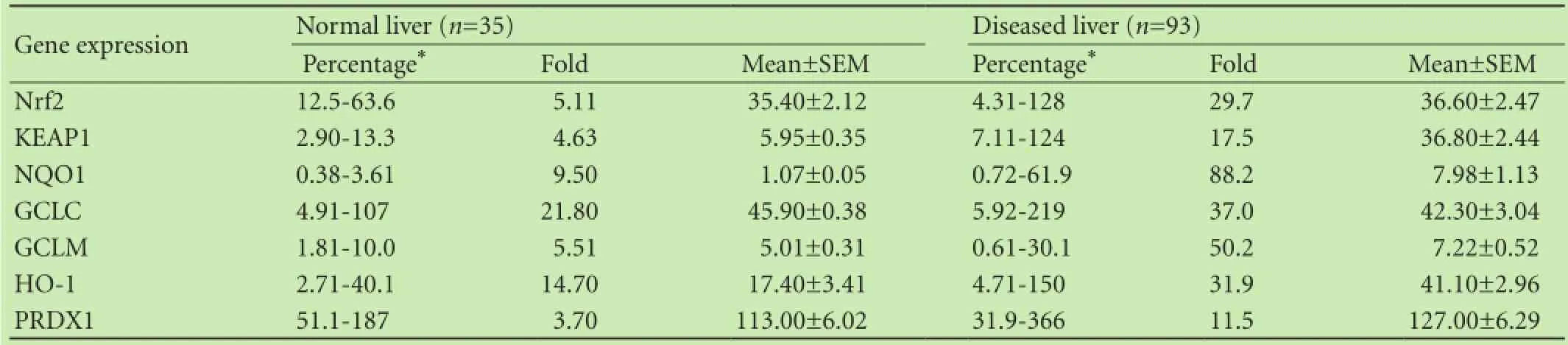
Table 3. Nrf2-related gene expressions under different pathological conditions of the liver
NQO1 expression
The mRNA expression of NQO1 is shown in Fig. 2. NQO1 expression in normal livers was low with an average of 1% of GAPDH, and its expression was increased in all diseased liver samples, with HCC the highest (18-fold increase over normal livers), followed by alcoholic cirrhosis (6-fold), end-stage liver disease (5-fold), peri-HCC (4-fold) and HBV-related cirrhosis (3-fold).
Since the up-regulation of NQO1 in HCC was significant, the individual expressions in HCC and peri-HCC tissues were examined. Most of HCC tissues had increased NQO1 expression, the biggest increase was seen in patient 2, and mild increase in patient 5 (Fig. 2B). Increased NQO1 mRNA levels were not observed in 5 of the 24 patients, i.e., the patient 4, 9, 10, 16 and 24. The patient 7 had a decreased NQO1 mRNA level, reflecting 88-fold individual variations in NQO1 mRNA levels of HCC samples.
GCLC and GCLM expression
The mRNA expressions of GCLC and GCLM, two key enzyme genes for glutathione synthesis, are shown in Fig. 3. The expression of GCLC in peri-HCC tissues is higher than that in normal livers and HCC. The expression of GCLC was decreased in HCC by 50% of normal livers. The expression of GCLC in HBV-related cirrhosis, alcoholic cirrhosis and end-stage liver disease was not statistically significant.
The mRNA expression of GCLM in peri-HCC showed the similar pattern as GCLC in which peri-HCC had higher GCLM mRNA levels than normal livers and HCC (Fig. 3B). GCLM mRNA levels were also higher in HBV-related cirrhosis and end-stage liver disease, while there was no difference in alcoholic cirrhotic livers.
HO-1 and PRDX1 expression
The mRNA expressions of HO-1 and PRDX1, two antioxidant enzyme genes targeted by Nrf2, are shown in Fig. 4. Compared to the normal livers, the expression of HO-1 was increased in peri-HCC tissue, HBV-related cirrhosis, alcoholic cirrhosis and end-stage liver disease, but it was unchanged in HCC. There was a significant difference in HO-1 mRNA levels between peri-HCC and HCC.
The basal expression of PRDX1 was slightly higher in 93 diseased livers (127.0%±6.3% of GAPDH) than that of 35 normal livers (113.0%±6.0% of GAPDH). However, the only difference was a higher PRDX1 mRNA level of 50% in peri-HCC than in normal livers, and it was also higher than HCC. PRDX1 mRNA levels were unchanged in the diseased livers such as end-stage liver disease and alcoholic cirrhosis.
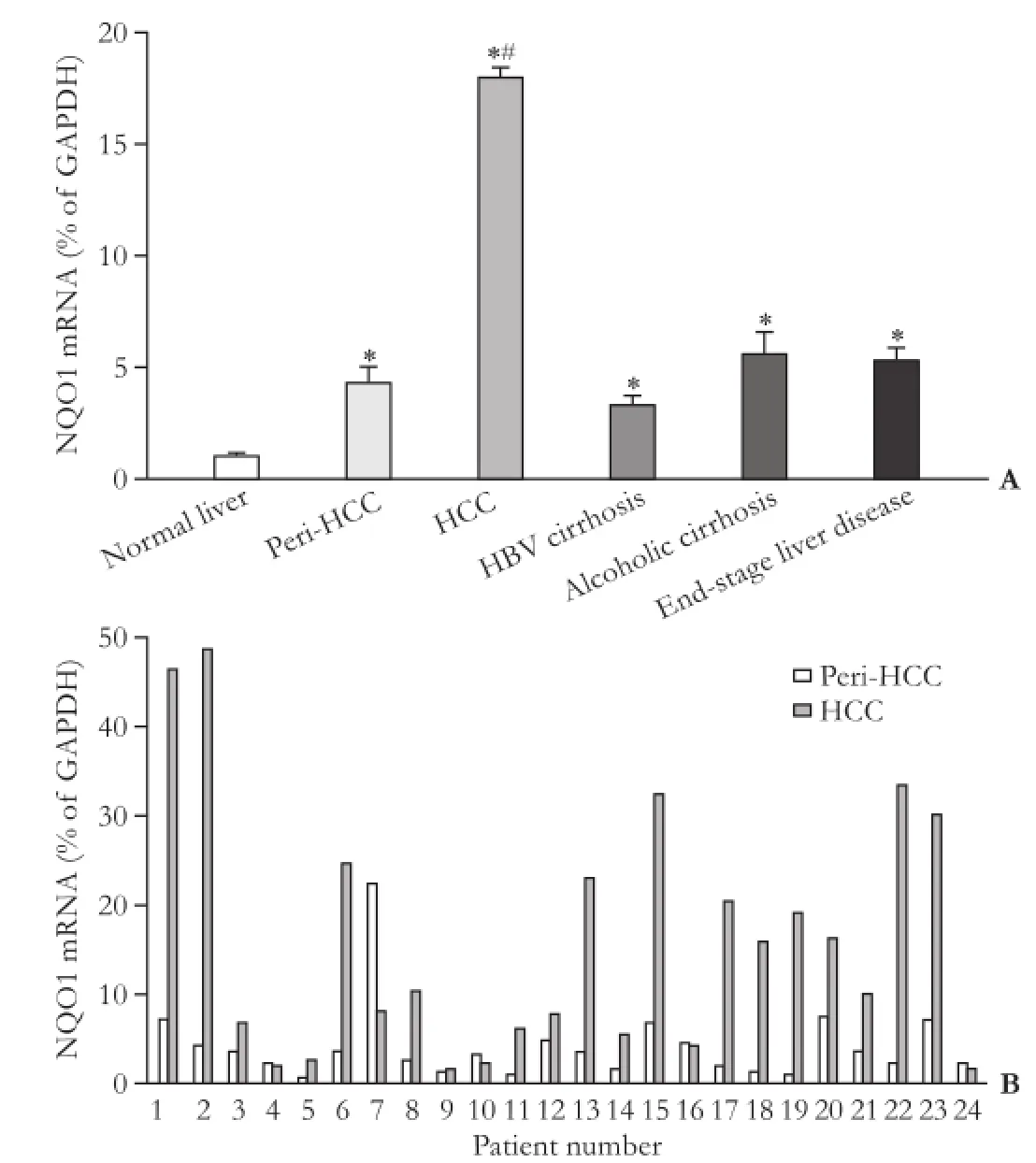
Fig. 2. Expression of NQO1 in human livers. The bottom panel represents individual HCC and corresponding peri-HCC tissues. *: significantly different from normal liver, P<0.05; #: significantly different from peri-HCC tissue, P<0.05.
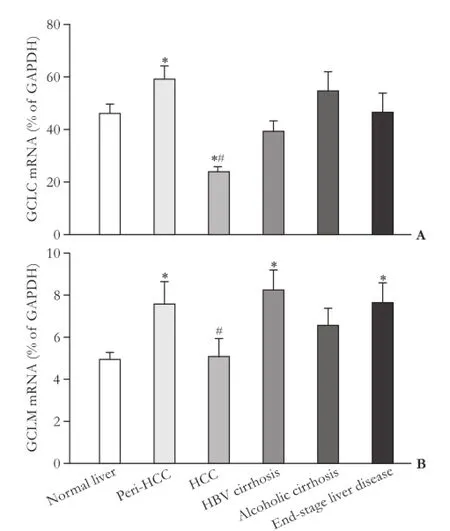
Fig. 3. Expression of GCLC and GCLM in human livers. *: significantly different from normal liver, P<0.05; #: significantly different from peri-HCC tissue, P<0.05.
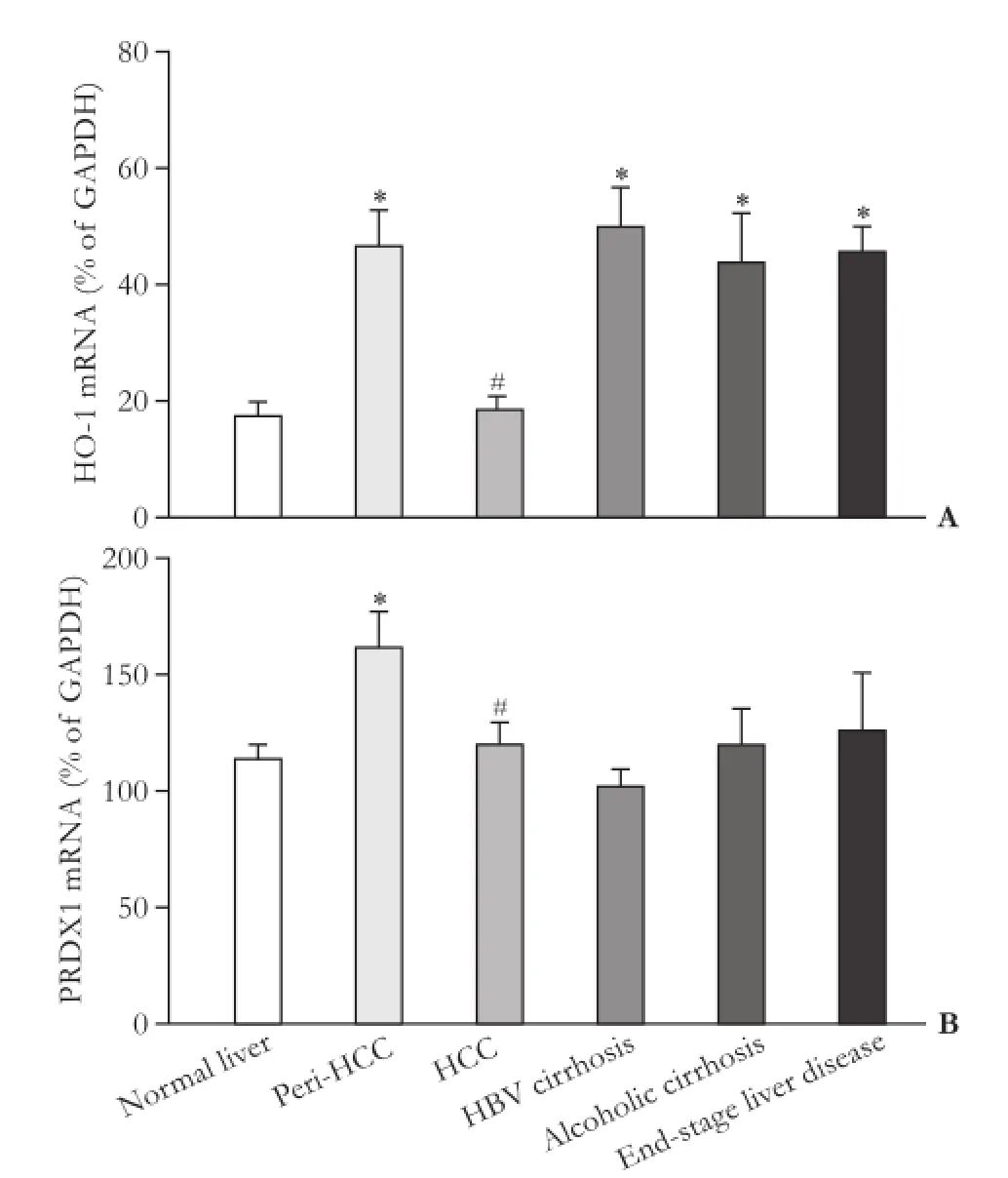
Fig. 4. Expression of HO-1 and PRDX1 in human livers. *: significantly different from normal liver, P<0.05; #: significantly different from peri-HCC tissue, P<0.05.
Discussion
The present study examined Nrf2 and Nrf2-related gene expressions in 35 normal and 93 diseased liver samples obtained from the Oriental Liver Transplant Center, including HCC, peri-HCC tissue, HBV-related cirrhosis, alcoholic cirrhosis and end-stage liver disease at various ages.[14]In general, the Nrf2-related gene expressions were prone to change under the pathological conditions of the liver, and the most notable gene changes were Nrf2 (decreased in HCC), KEAP1 (increased in diseased livers), NQO1 (increased in all diseased livers, especially in HCC), GCLC, GCLM (increased in peri-HCC) and HO-1 (increased in diseased livers except HCC). This is an extension of our recent work with P450 using the same liver samples.[14]
Nrf2 is a master regulator of cellular defense against oxidative stress and essential in regulating a network of genes involved in maintaining redox states and host defense mechanisms and critical for liver functions.[1-5]Mice deficient in Nrf2 (Nrf2-null) are sensitive to the increased hepatotoxicity produced by acetaminophen,[18]carbon tetrachloride,[19]arsenic,[20]1-bromopropane,[21]alcohol,[22]the MCD diet-induced steatohepatitis[23-25]and HFD-induced steatosis.[24,26]In contrast, mice overexpressing Nrf2 by knocking down its negative regulator KEAP1 (KEAP1-Kd) or hepatocyte-specific knockdown KEAP1 with maximum Nrf2 expression in liver (KEAP1-HKO) are resistant to a panel of hepatotoxicants such as cadmium,[15]ethanol,[16]phalloidin[17]and many others.[2]Thus, the role of Nrf2 in protecting liver damage has been extensively documented using genetic Nrf2 modulation animal models, as well as by using pharmacological Nrf2 activators.[5,27-29]
In comparison, little is known about the role of Nrf2 in human liver diseases. It is documented that Nrf2 signaling are involved in NAFLD,[10,12]and biliary cirrhosis.[11]Since the availability of human liver sample is very limited, analysis of surgically removed liver tissues for Nrf2 expression is of significance.[13]However, the sample size of above published work is relatively small, and the results obtained are somewhat contradictory. From the ethnicity standpoints, there is as yet no report on the expression of Nrf2 in Chinese patients with liver diseases. The current study utilized a total of 128 human liver specimens from 104 patients including 35 dissected normal livers and 93 diseased liver samples from the Oriental Liver Transplantation Center (Beijing, China) to examine the expression of Nrf2 and Nrf2-related genes.
We found that the expression of Nrf2 was lower in 24 HCC samples than in their peri-HCC tissues; however, the mRNA of Nrf2 was slightly increased in endstage liver disease and alcoholic cirrhosis (Fig. 1). The negative Nrf2 regulator KEAP1, on the other hand, was up-regulated to 6-fold on average in all diseased livers. A mutation in KEAP1 has been reported in human solid cancers,[30]and the down-regulation of Nrf2 in HCC[13]has been noted. It is likely that up-regulated KEAP1 may be responsible for the lower expression of Nrf2 in HCC, as it may act as a negative regulator of Nrf2.[5,6,8]
The Nrf2-related NQO1 was significantly increased in all of the diseased livers including peri-HCC tissues. Up-regulation of NQO1 in liver diseases was reported,[31]and NQO1 over-expression in human liver tumors was reported 20 years ago.[32]NQO1 is over-expressed in many cancer cell lines and human cancer tissues, NQO1 has protective effect against cancer.[33]On the other hand, NQO1 over-expression also confers chemo-resistance in human cancers.[34]In the present study, although Nrf2 was decreased in HCC, an 18-fold induction of NQO1 in HCC suggested that (1) NQO1 could be a sensitive biomarker for HCC, and (2) the increase of NQO1 does not necessarily match with the expression of Nrf2.
Different expression of Nrf2 regulates genes such as GCLC, GCLM and PRDX1 in liver diseases was also observed in the present study. In a study of 24 end-stage livers, the expression of GCLC was down-regulated,[13]which is consistent with our observation of downregulation of GCLC in HCC. In the present study, GCLMmRNA levels were similar to GCLC, slightly decreased in HCC, but increased in HBV-related cirrhosis and endstage liver disease.
HO-1 is an important molecule playing roles in alcoholic or viral hepatitis, chronic inflammation, fibrosis, cirrhosis and HCC.[35]HO-1 induction resulted in the protection of liver damage, down-regulation of proinflammatory cytokines, and interference with fibrosis progression in experimental animals.[35,36]On the other hand, HO-1 is over-expressed in various tumors including HCC, and proposed as a molecular brake on HCC migration.[37]In the present study, HO-1 was increased in chronic liver diseases, consistent with the role of HO-1 as an adaptive response. However, with the disease progression to HCC, increase in HO-1 was attenuated which is consistent with the notion that down-regulation of HO-1 correlated with HCC malignant progression.[38]
PRDX1 is a member of the peroxiredoxin family. Aberrant expression of PRDX1 has been described in various cancers including HCC, and has been proposed to act as a pro-cancer protein in liver diseases.[39]The present study showed that PRDX1 was over-expressed in peri-HCC, and unaltered in other end-stage diseased livers.
The Nrf2/KEAP1 pathway is also recognized as a "double-edged sword", its activation acts as a master defense against oxidative stress and protects cells from tumor formation, while Nrf2 is over-expressed in many cancer cells, and promotes tumor growth and metastasis.[33,40]Opposite to our initial expectation, we found that the expression of Nrf2 in HCC was decreased, but increased in cirrhotic livers. Perhaps at the end-stage of HCC, the regulation on Nrf2 is different from the earlystage of HCC, which needs further investigation. KEAP1 is proposed as "eye on the target" for the Nrf2 regulation in cancer chemoprevention,[41]KEAP1-knockout mice have over-expressed Nrf2 in the liver,[2,8]as KEAP1 is a negative regulator of Nrf2. However, the present study found that both KEAP1 and Nrf2 were increased in cirrhotic liver, suggesting that Nrf2 and KEAP1 could both be over-expressed in human liver diseases, depending on the progression stages of liver diseases.
In summary, this study profiled the constitutive expression of 7 major Nrf2-related genes in different liver diseases. Nrf2 and Nrf2-related genes were aberrantly expressed in cirrhotic livers and HCC, especially the significant increases in NQO1 could be an important biomarker for human liver disease progression.
Acknowledgments: The authors thank Dr. Curtis D. Klaassen for the guidance and critical comments on this work.
Contributors: CML and LJ conceived and designed the experiments; CH and SZY collected the samples; LYF and LJ performed RT-PCR analysis; CML, LYF and LJ wrote the draft. All authors contributed to the design and interpretation of the study and to further drafts. CML is the guarantor.
Funding: This work was supported by grants from the Chinese 863 Project (2012AA022409) and Guizhou Science and Technology Foundation (2009-70019).
Ethical approval: This study was approved by the Institutional Review Board of the General Hospital of Chinese People's Armed Police Forces and the Guiyang Medical College Human Subject Committee because the specimens were obtained without identification to the authors of the study.
Competing interest: No benefits in any form have been received or will be received from a commercial party related directly or indirectly to the subject of this article.
References
1 Klaassen CD, Reisman SA. Nrf2 the rescue: effects of the antioxidative/electrophilic response on the liver. Toxicol Appl Pharmacol 2010;244:57-65.
2 Liu J, Wu KC, Lu YF, Ekuase E, Klaassen CD. Nrf2 protection against liver injury produced by various hepatotoxicants. Oxid Med Cell Longev 2013;2013:305861.
3 Aleksunes LM, Manautou JE. Emerging role of Nrf2 in protecting against hepatic and gastrointestinal disease. Toxicol Pathol 2007;35:459-473.
4 Shin SM, Yang JH, Ki SH. Role of the Nrf2-ARE pathway in liver diseases. Oxid Med Cell Longev 2013;2013:763257.
5 Yang JJ, Tao H, Huang C, Li J. Nuclear erythroid 2-related factor 2: a novel potential therapeutic target for liver fibrosis. Food Chem Toxicol 2013;59:421-427.
6 Itoh K, Mimura J, Yamamoto M. Discovery of the negative regulator of Nrf2, Keap1: a historical overview. Antioxid Redox Signal 2010;13:1665-1678.
7 Taguchi K, Motohashi H, Yamamoto M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011;16:123-140.
8 Wu KC, Cui JY, Klaassen CD. Effect of graded Nrf2 activation on phase-I and -II drug metabolizing enzymes and transporters in mouse liver. PLoS One 2012;7:e39006.
9 Wang YP, Cheng ML, Zhang BF, Mu M, Zhou MY, Wu J, et al. Effect of blueberry on hepatic and immunological functions in mice. Hepatobiliary Pancreat Dis Int 2010;9:164-168.
10 Hardwick RN, Fisher CD, Canet MJ, Lake AD, Cherrington NJ. Diversity in antioxidant response enzymes in progressive stages of human nonalcoholic fatty liver disease. Drug Metab Dispos 2010;38:2293-2301.
11 Kawata K, Kobayashi Y, Souda K, Kawamura K, Sumiyoshi S, Takahashi Y, et al. Enhanced hepatic Nrf2 activation after ursodeoxycholic acid treatment in patients with primary biliary cirrhosis. Antioxid Redox Signal 2010;13:259-268.
12 Takahashi Y, Kobayashi Y, Kawata K, Kawamura K, Sumiyoshi S, Noritake H, et al. Does hepatic oxidative stress enhance activation of nuclear factor-E2-related factor in patients with nonalcoholic steatohepatitis? Antioxid Redox Signal 2014;20: 538-543.
13 Kurzawski M, Dziedziejko V, Urasińska E, Post M, Wójcicki M, Miętkiewski J, et al. Nuclear factor erythroid 2-like 2 (Nrf2) expression in end-stage liver disease. Environ Toxicol Pharmacol 2012;34:87-95.
14 Chen H, Shen ZY, Xu W, Fan TY, Li J, Lu YF, et al. Expressionof P450 and nuclear receptors in normal and end-stage Chinese livers. World J Gastroenterol 2014;20:8681-8690.
15 Wu KC, Liu JJ, Klaassen CD. Nrf2 activation prevents cadmium-induced acute liver injury. Toxicol Appl Pharmacol 2012;263:14-20.
16 Wu KC, Liu J, Klaassen CD. Role of Nrf2 in preventing ethanol-induced oxidative stress and lipid accumulation. Toxicol Appl Pharmacol 2012;262:321-329.
17 Lu YF, Liu J, Wu KC, Klaassen CD. Protection against phalloidin-induced liver injury by oleanolic acid involves Nrf2 activation and suppression of Oatp1b2. Toxicol Lett 2014;232:326-332.
18 Enomoto A, Itoh K, Nagayoshi E, Haruta J, Kimura T, O'Connor T, et al. High sensitivity of Nrf2 knockout mice to acetaminophen hepatotoxicity associated with decreased expression of ARE-regulated drug metabolizing enzymes and antioxidant genes. Toxicol Sci 2001;59:169-177.
19 Xu W, Hellerbrand C, Köhler UA, Bugnon P, Kan YW, Werner S, et al. The Nrf2 transcription factor protects from toxininduced liver injury and fibrosis. Lab Invest 2008;88:1068-1078.
20 Jiang T, Huang Z, Chan JY, Zhang DD. Nrf2 protects against As(III)-induced damage in mouse liver and bladder. Toxicol Appl Pharmacol 2009;240:8-14.
21 Liu F, Ichihara S, Valentine WM, Itoh K, Yamamoto M, Sheik Mohideen S, et al. Increased susceptibility of Nrf2-null mice to 1-bromopropane-induced hepatotoxicity. Toxicol Sci 2010;115: 596-606.
22 Lamlé J, Marhenke S, Borlak J, von Wasielewski R, Eriksson CJ, Geffers R, et al. Nuclear factor-eythroid 2-related factor 2 prevents alcohol-induced fulminant liver injury. Gastroenterology 2008;134:1159-1168.
23 Zhang YK, Yeager RL, Tanaka Y, Klaassen CD. Enhanced expression of Nrf2 in mice attenuates the fatty liver produced by a methionine- and choline-deficient diet. Toxicol Appl Pharmacol 2010;245:326-334.
24 Sugimoto H, Okada K, Shoda J, Warabi E, Ishige K, Ueda T, et al. Deletion of nuclear factor-E2-related factor-2 leads to rapid onset and progression of nutritional steatohepatitis in mice. Am J Physiol Gastrointest Liver Physiol 2010;298:G283-294.
25 Chowdhry S, Nazmy MH, Meakin PJ, Dinkova-Kostova AT, Walsh SV, Tsujita T, et al. Loss of Nrf2 markedly exacerbates nonalcoholic steatohepatitis. Free Radic Biol Med 2010;48: 357-371.
26 Wang C, Cui Y, Li C, Zhang Y, Xu S, Li X, et al. Nrf2 deletion causes "benign" simple steatosis to develop into nonalcoholic steatohepatitis in mice fed a high-fat diet. Lipids Health Dis 2013;12:165.
27 Bataille AM, Manautou JE. Nrf2: a potential target for new therapeutics in liver disease. Clin Pharmacol Ther 2012;92:340-348.
28 Okada K, Warabi E, Sugimoto H, Horie M, Tokushige K, Ueda T, et al. Nrf2 inhibits hepatic iron accumulation and counteracts oxidative stress-induced liver injury in nutritional steatohepatitis. J Gastroenterol 2012;47:924-935.
29 Zhou R, Lin J, Wu D. Sulforaphane induces Nrf2 and protects against CYP2E1-dependent binge alcohol-induced liver steatosis. Biochim Biophys Acta 2014;1840:209-218.
30 Yoo NJ, Kim HR, Kim YR, An CH, Lee SH. Somatic mutations of the KEAP1 gene in common solid cancers. Histopathology 2012;60:943-952.
31 Aleksunes LM, Goedken M, Manautou JE. Up-regulation of NAD(P)H quinone oxidoreductase 1 during human liver injury. World J Gastroenterol 2006;12:1937-1940.
32 Belinsky M, Jaiswal AK. NAD(P)H: quinone oxidoreductase1 (DT-diaphorase) expression in normal and tumor tissues. Cancer Metastasis Rev 1993;12:103-117.
33 Lau A, Villeneuve NF, Sun Z, Wong PK, Zhang DD. Dual roles of Nrf2 in cancer. Pharmacol Res 2008;58:262-270.
34 Wang XJ, Sun Z, Villeneuve NF, Zhang S, Zhao F, Li Y, et al. Nrf2 enhances resistance of cancer cells to chemotherapeutic drugs, the dark side of Nrf2. Carcinogenesis 2008;29:1235-1243.
35 Sass G, Barikbin R, Tiegs G. The multiple functions of heme oxygenase-1 in the liver. Z Gastroenterol 2012;50:34-40.
36 Luo YH, Li ZD, Liu LX, Dong GH. Pretreatment with erythropoietin reduces hepatic ischemia-reperfusion injury. Hepatobiliary Pancreat Dis Int 2009;8:294-299.
37 Zou C, Zhang H, Li Q, Xiao H, Yu L, Ke S, et al. Heme oxygenase-1: a molecular brake on hepatocellular carcinoma cell migration. Carcinogenesis 2011;32:1840-1848.
38 Caballero F, Meiss R, Gimenez A, Batlle A, Vazquez E. Immunohistochemical analysis of heme oxygenase-1 in preneoplastic and neoplastic lesions during chemical hepatocarcinogenesis. Int J Exp Pathol 2004;85:213-222.
39 Aguilar-Melero P, Prieto-álamo MJ, Jurado J, Holmgren A, Pueyo C. Proteomics in HepG2 hepatocarcinoma cells with stably silenced expression of PRDX1. J Proteomics. 2013;79:161-171.
40 Pi J, Freeman ML, Yamamoto M. Nrf2 in toxicology and pharmacology: the good, the bad and the ugly? Toxicol Appl Pharmacol 2010;244:1-3.
41 Yates MS, Kensler TW. Keap1 eye on the target: chemoprevention of liver cancer. Acta Pharmacol Sin 2007;28:1331-1342.
Accepted after revision August 18, 2015
10.1016/S1499-3872(15)60425-8
Author Affiliations: The Affiliated Hospital of Guizhou Medical University, Guiyang 550004, China (Cheng ML); Key Lab for Basic Pharmacology of Ministry of Education, Zunyi Medical College, Zunyi 563003, China (Lu YF and Liu J); The Institute of Organ Transplantation, The General Hospital of Chinese People's Armed Police Forces, Beijing 100039, China (Chen H and Shen ZY)
Ming-Liang Cheng, MD, the Affiliated Hospital of Guizhou Medical University, Guiyang 550004, China (Tel: +86-851-86752795; Email: Chengml@21cn.com)
© 2015, Hepatobiliary Pancreat Dis Int. All rights reserved.
Published online September 21, 2015.
December 13, 2014
杂志排行

Hepatobiliary & Pancreatic Diseases International的其它文章
- Extrahepatic right portal vein ligation allows parenchyma-sparing en bloc resection of segments 7, 8 and 4a for liver tumors engaging the right and middle hepatic veins
- 18F-FDG PET/CT in differentiating malignant from benign origins of obstructive jaundice
- Diagnostic and prognostic roles of soluble CD22 in patients with Gram-negative bacterial sepsis
- CT-guided high-dose-rate brachytherapy in the interdisciplinary treatment of patients with liver metastases of pancreatic cancer
- Congenital extrahepatic portosystemic shunt complicated by the development of hepatocellular carcinoma
- Multidisciplinary management of Mirizzi syndrome with choIecystobiIiary fistuIa: the vaIue of minimaIIy invasive endoscopic surgery