影响杨树苗期光合速率多因素研究
2015-11-17汤玉喜李永进吴立勋
吴 敏, 汤玉喜, 唐 洁, 李永进, 吴立勋
(湖南省林业科学院, 湖南 长沙 410004)
影响杨树苗期光合速率多因素研究
吴 敏, 汤玉喜, 唐 洁, 李永进, 吴立勋
(湖南省林业科学院, 湖南 长沙 410004)
为了显示光合测定时所选叶片性状及测定时间对光合速率的影响,试验采用其他试验条件一致、研究目的条件按设计等级差异的方法进行光合速率的测定。试验结果显示:在同一光通量条件下,1年生苗干上叶的净光合速率可分为光合速率上升段、光合速率平稳段、光合速率下降段3个区分段;四足蟎不同为害程度叶的净光合速率下降13.2%~27.3%;杨树苗的净光合速率,上午极显著高于下午。ZH — 17、XL — 90无性系,在同一时间所发生的叶片总数相近,但保有功能旺盛叶的数量 ZH — 17比 XL — 90少,前者是后者的69%。因此,在植物光合速率测定时,不仅要选定叶龄相近、无病虫为害的正常叶,还要注意测定时间,不把上午测定结果与下午比较。
叶龄; 蟎虫为害; 测定时间; 净光合速率; 杨树苗
植物生长发育的有机物质来源于绿色植物光合产物的积累。光合产物的产量与其光合特性、叶片面积和立体空间结构等有关[ 1-4]。绿色植物的光合作用是十分灵敏的光-化学反应过程,有很多的酶参与其中,是复杂的生理生化反应过程[5 ],受影响的因子很多,但往往不被重视。林木光合特性研究的报告很多[6-11],但将光合潜能的发挥与其时叶片所处的生理生态环境、时空状态相联系的报道则不多见。本研究从同等试验条件、不同叶龄、不同虫害程度、不同测定时间时光合速率的变化入手,探讨在林木光合测定中最易忽视的内外因子对叶光合速率的影响,以求证何种试验方法所得试验结果才具可比性,更能客观反映杨树苗的光合本质。
1 试验材料与方法
1.1 叶龄与光合速率
试验用杨树为中汉 — 17(ZH — 17)及新选育的湘林 — 90(XL — 90)无性系[12],苗木为当年扦插的1年生苗,时间为6月中旬。苗干上的叶依发生顺序自下而上叶龄依次减小,试验用叶遂将苗干自下而上现存叶区分为9段,每段有叶4~5片,取其1叶进行光合速率测定。每叶测定在光子通量密度分别为100、800、1600μmol/(m2·s)时的净光合速率。试验重复3次。
1.2 四足蟎为害与光合速率
试验在XL — 90无性系1年生苗干中上段,取无蟎虫为害正常叶、虫口密度50~100轻度为害叶、虫口密度在100以上重度为害叶三种类型,分别测光 — 光响应曲线。光子通量密变分设为0、10、20、40、60、100、200、400、600、800、1000、1200、1400、1600、1800、2000μmol/(m2·s)等16个梯度。试验重复3次。
1.3 测定时间与光合速率
试验材料为XL — 90、XL — 77、XL75、XL — 101、XL — 92、ZH — 17、I — 69等7个无性系[13-14]1年生扦插苗,测定时间为6月中旬。参试叶为苗干中上部正常叶,每无性系分别在上午和下午测光 — 光响应曲线,光通量密度分设16个梯度。每无性系重复3次。以7个无性系上午和下午的均值检验试验差异的显著水平。
1.4 试验采用LI — 6400光合测定仪进行光合测定,数据采DPS数据处理系统进行统计分析。
2 结果与分析
2.1 苗期叶位叶龄对净光合速率的影响
黑杨在亚热带地区,1年生苗高达4~5m或更高,叶在苗干上分3个方位轮替向上生长。在叶的生长发育过程中,自下而上随叶位升高、叶龄变小,并随光通量条件变化、光照历程差异,其光合能力亦有所不同。因此,在对植物光合能力的研究过程中应重视叶的部位和叶龄。以XL — 90、ZH — 17两新、老无性系为例,至6月中旬,XL — 90和ZH — 17已发生的叶分别为58片、54片,基部老化且已脱落的叶分别为6片和l3片。在3种恒定光子通量密度条件下,不同部位叶的光合速率见图1、图2。
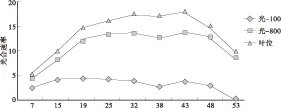
图1 XL — 90 光合速率与叶位Fig.1 XL — 90’s photosynthetic rate and leaf position

图2 ZH — 17 光合速率与叶位Fig.2 ZH — 17’s photosynthetic rate and leaf position
图1、图2显示,同一圃地XL — 90、ZH — 17两无性系,不同部位 — 叶龄叶分别在100、800、1600三个梯度μmol/(m2·s)光通量条件下,光合速率均随光通量梯度的增加而增加(ZH — 17的第49位叶例外);在同一光通量条件下,1年生苗干上叶的净光合速率可分为3个区分段。①光合速率上升段:即光合速率随叶位升高、叶龄变小、光合功能增强,光合速率呈增加趋势,如XL — 90的第7 — 15、ZH — 17的第14~20叶。在苗圃群体内,除叶龄因素外,苗干上的叶越往下叶所接受的光通量越小,较长时间的弱光照历程,也往往导致叶的光合能力的减弱。②光合速率平稳段: 光合速率维持在较高、较平稳的水平上,如XL — 90的第19~48、ZH — 17的第24~44叶。此段叶光合功能旺盛,是有机物积累的主要来源。③光合速率下降段:叶位升高、叶龄更小,此段叶的生长发育还处于早期、不完全成熟阶段,光合速率随叶位升高而降低,如XL — 90的第53、ZH — 17的第49及其以上各叶。无性系间这种变化趋势较为一致。光合功能在苗干上的这种叶位-叶龄差异,提示在田间杨树苗木光合速率的测定时,应尽量选准相似功能叶,以增加实验数据的可比性。
XL — 90、ZH — 17两无性系,在同一时间所发生的叶片总数较为接近,但二者功能旺盛叶的数量则有所不同: XL — 90无性系为29片,ZH — 17为20片,后者是前者的69%。可见,与XL — 90比较,ZH — 17苗木基部叶的老化、脱落进程较快,维持旺盛光合功能叶片的数量较少、时间较短。
2.2 蟎虫为害对净光合速率的影响
6—8月,杨树苗顶端的幼嫩叶有时易发生四足蟎为害。蟎虫吮吸叶汁,使叶片皱缩、变褐、变小,发育不良;严重时提早脱落,影响苗木生长。为检验蟎虫为害对光合速率的影响,XL — 90无性系在蟎虫不同为害程度叶的光 — 光响应曲线见图3。
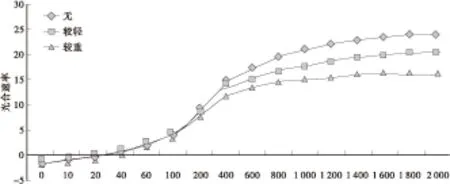
图3 蟎虫不同为害叶光 — 光响应曲线Fig.3 Leaf light and light response curves of different mite bug damage
光响应曲线显示,当光通量<200μmol/(m2·s)时,不同为害叶的光合速率非常接近,>400μmol/(m2·s)以后,不同为害叶光合曲线的分离则越来越大。方差分析结果见表1。

表1 蟎虫不同为害叶光合速率方差分析Tab.1 Varianceanalysisofleafphotosyntheticrateofdifferentmitebugdamage变异来源平方和自由度均方F值P值光通量梯度间3505.17815233.678685.2830.0000为害程度间94.2824247.141217.2050.0000误差82.2006302.74总变异3681.66247
表2显示,蟎虫不同为害的杨树苗光合速率的光响应曲线在光通量梯度间及为害程度间均有极显著差异(显著水平P<0.000 1)。对不同为害程度的光响应曲线作多重比较,结果见表2。
表2显示,蟎虫不同为害程度叶的光合速率均存在显著差异,尤其是重度为害能造成光合速率的极显著降低。就光合曲线均值看,重度为害的光合速率仅为正常叶的72.7%。可见,蟎虫防治对杨树育苗具有重要意义。

表2 蟎虫不同为害光合速率Tukey法多重比较Tab.2 Tukeymultiplecomparisonofleafphotosyntheticrateofdifferentmitebugdamage处理均值5%显著水平1%极显著水平无为害12.578aA轻为害10.919bAB重为害9.145cB
2.3 测定时间对光合速率的影响
田间林木光合速率测定常在白天整日进行,在光合速率非日变化研究情况下,则涉及上午和下午测定的结果是否存在差异及差异显著水平问题。为此,对XL — 90、ZH — 17、I — 69等7个杨树无性系苗光-光响应曲线分别上午和下午进行统计,见图4。经检验,光合速率在上、下午时间段存在极显著差异(P<0.000 1),多重比较见表3。
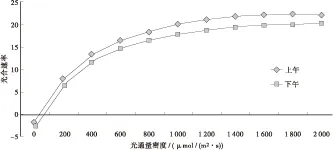
图4 测定时间与杨树苗期光合速率图Fig.4 The determination with time and the photosynthetic rate of poplar seedlings
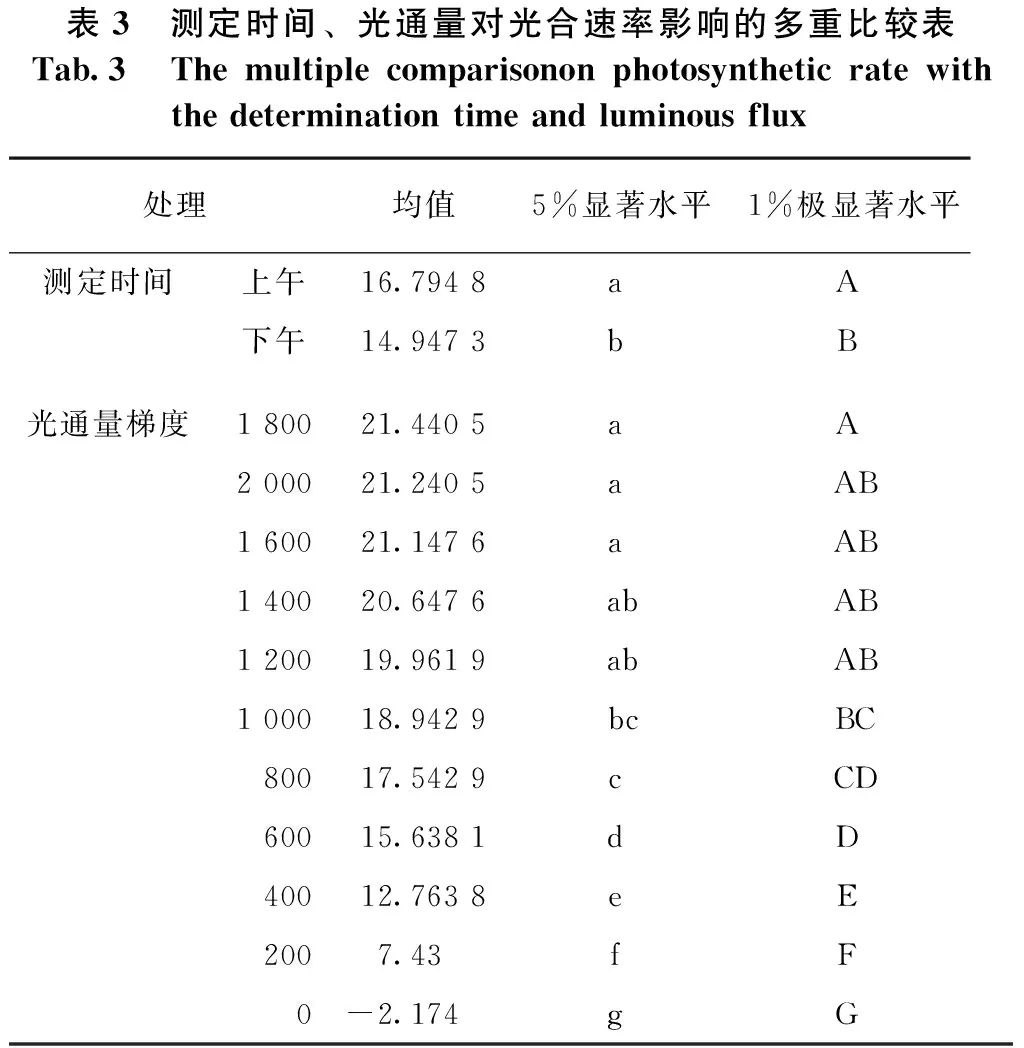
表3 测定时间、光通量对光合速率影响的多重比较表Tab.3 Themultiplecomparisononphotosyntheticratewiththedeterminationtimeandluminousflux处理均值5%显著水平1%极显著水平测定时间上午16.7948a A 下午14.9473b B 光通量梯度180021.4405a A 200021.2405a AB160021.1476a AB140020.6476abAB120019.9619abAB100018.9429bcBC80017.5429c CD60015.6381d D 40012.7638e E 2007.43f F 0-2.174g G
表3显示,杨树苗的光合速率,上午的测定数值比下午高,差异达极显著水平。如以光-光响应曲线均值计,下午的测定数值仅为上午的89%。这种差异的产生,可能源自叶片经过上午连续的光合作用,光合产物的输送不完全、在叶片中的积累增加,从而抑制了下午光合能力的充分发挥。此现象提示,叶的光合作用是绿色植物物质生产系统复杂的生理生化过程,光合产物的生产、运输可能存在源-壑关系。因此,光合速率的测定应在较短时间内完成,如此所测数据才具有较高可比性。
表3还显示,光合速率在光通量梯度间亦存在极显著差异,在光通量0~800 μmol/(m2·s)梯度间,光合速率随光通量增而增加,差异达显著水平;当光通量在1 200~2 000 μmol/(m2·s)间时,其光合速率无显著差异。因比,在1 200~2000 μmol/(m2·s)范围内任选某一光通量,所测光合速率基本上都能够反映杨树苗期的最大光合能力。
3 结论与讨论
(1) 1年生杨树苗干上叶的净光合速率可分为3个区分段:光合速率上升段、光合速率平稳段、光合速率下降段。
(2) XL — 90、ZH — 17两无性系,在同一时间所发生的叶片总数较为接近,但二者在光合速率平稳段保有功能旺盛叶的数量则有所不同: ZH — 17无性系是XL — 90的69%。无性系间叶的这种生长发育过程差异往往会导致林木个体间光合功能旺盛叶的总量产生差异,从而导致有效干物质积累的总叶面积存在差异,有可能导致无性系间的生长量差异,叶的这种生长发育特性可为杨树无性系育种提供参考性指标。
(3) 杨树苗期受蟎虫为害后,叶的光合速率将下降13.2%~27.3%。考虑到叶的发育不全、叶面积缩小、提早脱落,其为害程度将会更大。
(4) 杨树苗的光合速率,下午测定数值比上午低,差异达极显著水平。因此,为增加光合试验数据的可比性,光合测定时不仅要重视叶位 — 叶龄的同等性,还要重视测定时间的同等性,以提高光合试验准确性。
(5) 光通量密度梯度在0~800 μmol/(m2·s)范围内,光合速率随光通量增加而增加,且均有显著差异;在1 200~2 000 μmol/(m2·s)间,其光合速率无显著差异。因此,杨树苗圃的育苗密度,应尽力使苗干中部光合功能旺盛叶较长时间处于800~1 200 μmol/(m2·s)光通量条件下,以获取最大光能利用效力和土地利用率。
[1] 王金辉, 李轶女, 倪丕冲,等.叶绿体转化体系研究进展[J].生物技术通报,2012(1):1-6.
[2] 巩智刚, 周海鹏, 徐芳,等. 叶绿体转化及其应用于作物改良研究的最新进展[J].核农学报,2012,26(2):288-294.
[3] 周菲, 路史展, 高亮,等.植物质体基因工程:新的优化策略及应用[J].遗传,2015(8):777-792.
[4] 孙洪助, 林春波,孙文华,等. 光质对作物光合作用影响的研究进展[J].2015(23):14-16.
[5] 夏来坤,刘京宝, 黄璐,等.植物光合作用生物增效剂对夏玉米生长及产量品质的影响[J].中国农学通报,2015,32(3)122-125.
[6] 盛杰; 陈月华; 吴际友;等.闽楠家系苗期光合特性的研究[J].中南林业科技大学学报,2015,36(6):45-49
[7] 王真真, 李宏,苗乾乾,等.坐果期不同灌溉条件下枣树光合特性研究[J].中南林业科技大学学报,2015,36(5):59-63.
[8] 李晓芳,杨瑞,安明态,等.岩生红豆苗木光合特性初步研究[J]. 安徽农业科学,2015(23):45-47.
[9] 陈淑钦, 陈清西.铁皮石斛栽培及光合特性研究进展[J].江苏农业科学,2015(7):66-70.
[10] 周永斌, 马学文,姚鹏,等. 不同生长速度杨树品种的光合生理特性研究[J]. 沈阳农业大学学报,2007,56(3):336-339.
[11] 王小平;诸葛强. 转PEPE基因杨树的光合特性分析[D].南京:南京林业大学,2010.
[12] 吴敏, 吴立勋,汤玉喜;等. 湘林—90等5个美洲黑杨新无性系速丰性能研究[J].湖南林业科技,2008,35(6):13-20.
[13] 汤玉喜, 刘友全,吴敏,等. 淹水胁迫对美洲黑杨无性系保护酶系统的影响[J].中南林业科技大学学报,2008,29(5):59-63.
[14] 汤玉喜,刘友全, 吴敏,等. 淹水胁迫下美洲黑杨无性系生理生化指标的变化 [J].中南林业科技大学学报,2008,29(5):59-63.
[15] 唐洁,汤玉喜, 李永进,等.美洲黑杨无性系苗期生长与蒸腾特性研究[J].湖南林业科技,2013,40(6):9-13.
[16] 刘炳梅,齐芪,刘淑欣,等.5种杨树苗期形态及生理结构特征比较初探[J].北京林业大学学报,2015,37(6)35-44.
Theseveralaffectingfactorsonthephotosyntheticrateofpoplarinseedlingstage
WU Min, TANG Yuxi, TANG Jie, LI Yongjin, WU Lixun
(Hunan Academy of Forestry, Changsha 410004, China)
In order to show the effects on the photosynthetic rateof the selected leaf characters and the testing time, the experiment used the method to determine the rate of photosynthetic rate by the different class of the research purpose in same others conditions. The test results show that, under the same flux conditions, one year seedling stem leaf’s net photosynthetic rate can be divided into 3 section of photosynthetic rate of ascent, photosynthetic rate stable period, photosynthetic rate decreased; the leaf net photosynthetic rate by mite bug of different damage decreased 13.2%~27.3%; The net photosynthetic rate of poplar seedlings in the morning extremely significantly higher than it in the afternoon. Although ZH-17 and XL-90 clones in the same time and similar total number of leaves, but retains the quantity ZH-17 function effective leaves was less than XL-90 69% of XL-90. Therefore, in plant photosynthetic rate determination, not only to selected similar leaf age, no plant diseases and insect pests of normal leaf, but also pay attention to the determination of time, don’t compare the determination results with the morning and afternoon.
leaf age; pest mite; determination time; net photosynthetic rate; poplar seedlings
2015-09-29
湖南省林业厅科技计划项目“杨树工程结构材定向培育及增值利用关键技术研究与示范”(XLK201506)
S 792.11.01
A
1003 — 5710(2015)06 — 0094 — 05
10. 3969/j. issn. 1003 — 5710. 2015. 06. 018
(文字编校:杨 骏)
