不同种源木荷种子形态变异研究
2015-11-17汤玉喜李永进钟文斌
唐 洁, 汤玉喜, 杨 艳, 李永进, 吴 敏, 刘 秀, 钟文斌
(1.湖南省林业科学院, 湖南 长沙 410004; 2.广西壮族自治区林业科学研究院, 广西 南宁 530001;3.中国林业科学研究院亚林中心, 江西 分宜 336600)
不同种源木荷种子形态变异研究
唐 洁1, 汤玉喜1, 杨 艳1, 李永进1, 吴 敏1, 刘 秀2, 钟文斌3
(1.湖南省林业科学院, 湖南 长沙 410004; 2.广西壮族自治区林业科学研究院, 广西 南宁 530001;3.中国林业科学研究院亚林中心, 江西 分宜 336600)
对10个木荷种源的种子长、宽、长宽比及千粒重等性状变异进行研究。结果表明:木荷种源间种子长、宽、千粒重等的差异均达到了极显著水平,长宽比的差异达到了显著水平;种子长、宽、长宽比及千粒重的遗传力分别为0.7340、0.8682、0.6479和0.8165。木荷种子长与种子宽、年均温度均呈显著正相关;种子千粒重与北纬呈显著负相关,与年均温度呈显著正相关。木荷种子形态特征地理变异呈现区域板块变异模式和随机变异模式。
木荷; 种源; 种子; 形态变异
木荷(SchmasuperbeGardn.etchamp)又名荷树、荷木,属山茶科木荷属常绿乔木,是我国南方主要的乡土阔叶树种[1-3]。因其木材坚韧、纹理交错、结构细致、加工容易、不开裂,木荷常作为木制工艺用材树种。同时,由于其树干高大通直,树冠浓密,叶片厚革质,含易燃脂少,耐火性强,萌芽力高,木荷也用作生物防火林带建设的主要树种。在裸岩和土层较薄的陡坡上的严酷生境条件下也能形成较为单纯的木荷林群落。木荷具有改良土壤、提高肥力、保持水土、涵养水源的功能,是高效的生态树种,近年又被用作庭院美化和行道绿化的优良树种。由于木荷分布区广,分布区内气候、土壤等因子的差异,造成其不同种源间种子的形状、品质存在一定的差异。王瑞辉、谢文雷等对木荷苗木繁育和丰产栽培等进行了研究[4-10],关于木荷遗传变异的研究很少。许多研究表明[11-14],林木种子性状存在着一定的地理种源变异,往往与分布区的光照、温度、降水量及海拔、纬度等环境因素有关,而这些因素又通常是相互作用、相互影响的。作者对木荷10个种源种子的形态变异及其与地理气候因子的关系进行研究,为木荷遗传改良、种源区划、种子生产等提供参考。
1 材料与方法
1.1 试验材料
在湖南省内及江西、广西等周边省、区的木荷天然分布区进行采种,于2014年10—11月采集10个种源的木荷种子,各种源地理气候因子见表1。采种林分选择当地种源形成的优良林分,林分年龄在20年左右,每个种源采种5~10株母树。种子采集后,做好标记,分别装袋,经自然干燥后置于通风阴凉处贮藏备用。
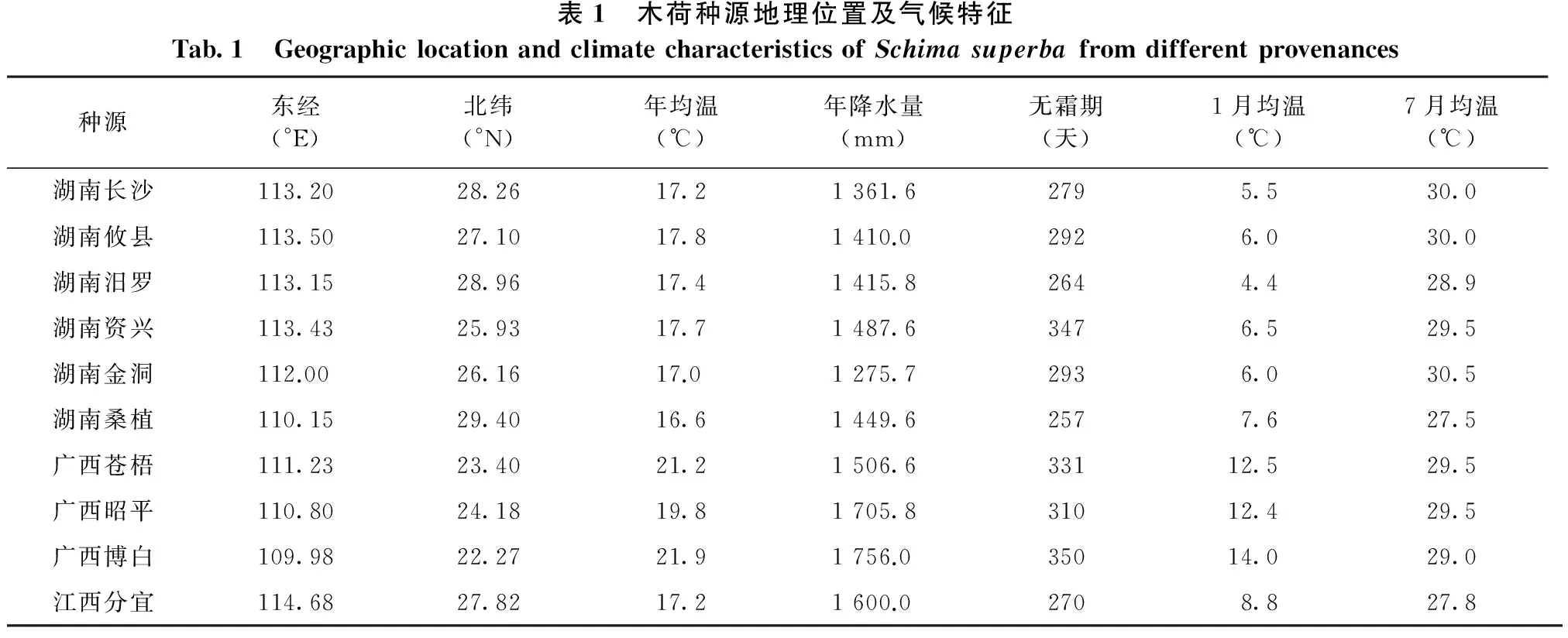
表1 木荷种源地理位置及气候特征Tab.1 GeographiclocationandclimatecharacteristicsofSchimasuperbafromdifferentprovenances种源东经(°E)北纬(°N)年均温(℃)年降水量(mm)无霜期(天)1月均温(℃)7月均温(℃)湖南长沙113.2028.2617.21361.62795.530.0湖南攸县113.5027.1017.81410.02926.030.0湖南汨罗113.1528.9617.41415.82644.428.9湖南资兴113.4325.9317.71487.63476.529.5湖南金洞112.0026.1617.01275.72936.030.5湖南桑植110.1529.4016.61449.62577.627.5广西苍梧111.2323.4021.21506.633112.529.5广西昭平110.8024.1819.81705.831012.429.5广西博白109.9822.2721.91756.035014.029.0江西分宜114.6827.8217.21600.02708.827.8
1.2 数据获取方法
(1) 种子形态指标的测定。每1个种源中每个家系随机取12粒种子,重复3次。用数显游标卡尺测量种子长度和宽度,并计算种子长宽比。测量精确到0.01mm。
(2) 千粒重测定。每个家系取100粒种子进行称重,重复3次。
(3) 遗传力计算。H2=1-1/F,其中H2为性状遗传力,F为方差分析中的F检验值[15]。
1.3 数据分析
采用SPSS19.0软件进行方差分析、相关分析和聚类分析等。
2 结果与分析
2.1 不同种源木荷种子形态特征差异性
林木种子的形态属一种较为稳定的性状,它比林木的根、茎、叶稳定性高[16]。对10个木荷种源种子形态性状进行统计分析,结果(表2)表明:种源间种子长度存在极显著性差异,平均长度为10.08mm;最长的是广西苍梧种源,为11.17 mm;最短的是湖南资兴种源,为9.30mm。种子宽度存在极显著性差异,平均宽度为5.18mm;最宽的为湖南汨罗种源,为6.02mm;最窄的是湖南资兴和湖南桑植种源,均为4.68mm。种子长宽比存在显著性差异,平均长度比值为1.96;长宽比最大的是湖南长沙和广西苍梧种源,均为2.11;最小的是湖南汨罗种源,为1.75。种子千粒重存在极显著性差异,千粒重平均值为5.08g;最大的是广西昭平种源,达6.8g;最小的是湖南桑植种源,仅为3.12g。变异系数表示性状的离散程度。木荷种源间种子长、宽、长宽比、千粒重的的变异系数分别为5.10%、5.11%、5.19%和10.90%,说明木荷种源内种子形态性状比较稳定,变幅较小。木荷种源间种子长、宽、长宽比、千粒重的遗传力分别为0.7340、0.8682、0.6479和0.8165,表明这些性状受到中等程度以上的遗传控制。这为木荷的遗传改良提供了可靠保障。
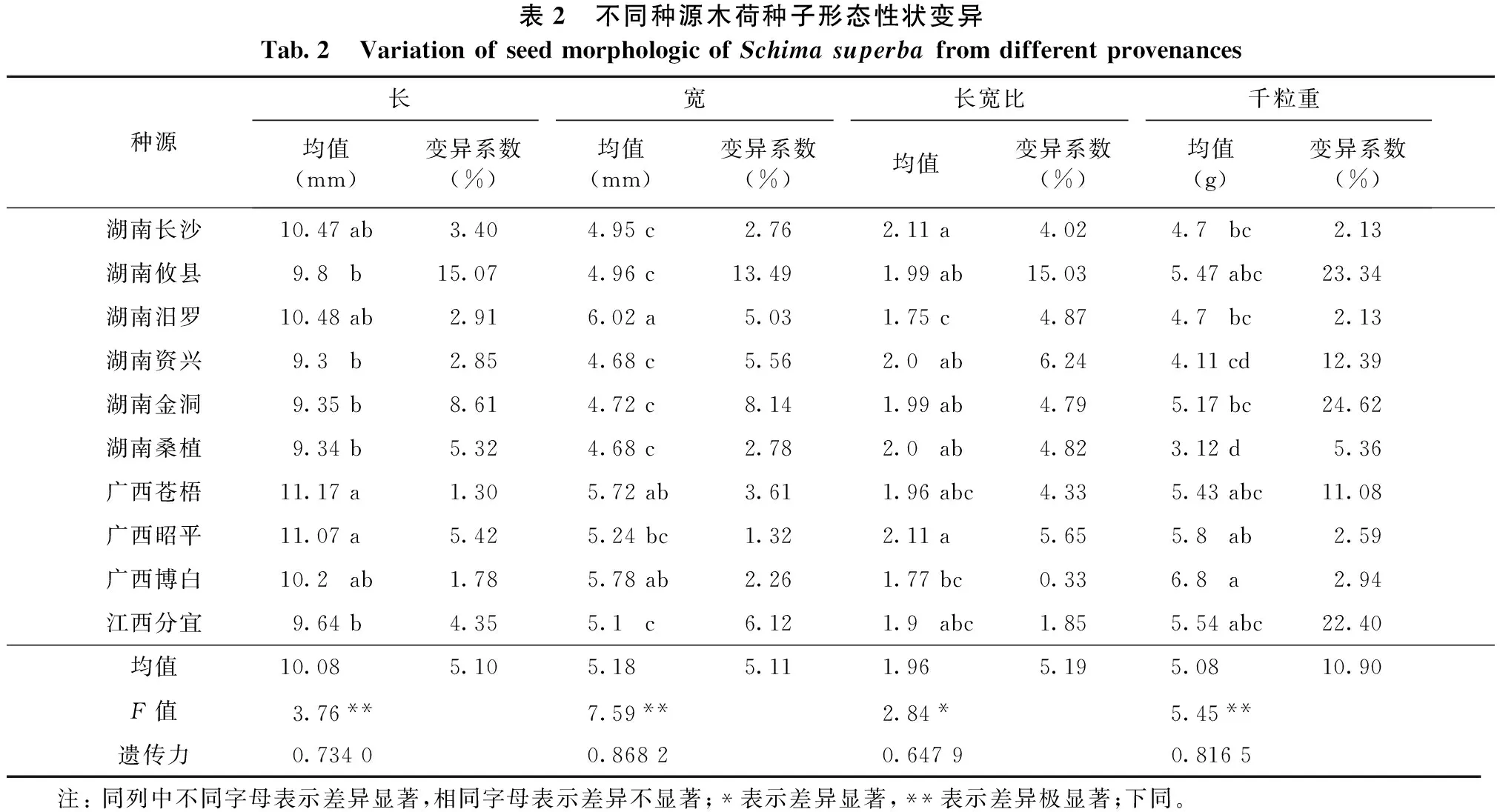
表2 不同种源木荷种子形态性状变异Tab.2 VariationofseedmorphologicofSchimasuperbafromdifferentprovenances种源长宽长宽比千粒重均值(mm)变异系数(%)均值(mm)变异系数(%)均值变异系数(%)均值(g)变异系数(%)湖南长沙10.47ab3.404.95c2.762.11a4.024.7 bc2.13湖南攸县 9.8 b 15.074.96c13.491.99ab15.035.47abc23.34湖南汨罗10.48ab2.916.02a5.031.75c4.874.7 bc2.13湖南资兴 9.3 b 2.854.68c5.562.0 ab6.244.11cd12.39湖南金洞 9.35b 8.614.72c8.141.99ab4.795.17bc24.62湖南桑植 9.34b 5.324.68c2.782.0 ab4.823.12d5.36广西苍梧11.17a 1.305.72ab3.611.96abc4.335.43abc11.08广西昭平11.07a 5.425.24bc1.322.11a5.655.8 ab2.59广西博白10.2 ab1.785.78ab2.261.77bc0.336.8 a2.94江西分宜 9.64b 4.355.1 c6.121.9 abc1.855.54abc22.40均值10.085.105.185.111.965.195.0810.90F值3.76**7.59**2.84*5.45**遗传力0.73400.86820.64790.8165 注:同列中不同字母表示差异显著,相同字母表示差异不显著;*表示差异显著,**表示差异极显著;下同。
2.2 不同种源木荷种子形态性状与地理环境因子的相关性
利用不同种源种子各性状间内在的联系,通过对容易测定的性状进行预测和间接选择,可以降低改良成本,提高选择效率[17]。不同种源木荷种子形态性状及地理环境因子相关分析结果(表3)表明:木荷种子长与种子宽、年均温度均呈显著正相关;种子千粒重与北纬呈显著负相关,与年均温度呈显著正相关。说明木荷种子的长度与种子宽度及当地年均温度变化有关,随着种子宽度的增加和年均温度的升高而增长;千粒重受地理环境因素中北纬、年均温度的影响较大。通过对不同种源木荷种子性状与地理气候因子的相关分析,可以了解不同地理环境条件对种子性状的影响,为种子区划与良种筛选提供依据[18]。

表3 木荷种子形态性状与地理环境因子的相关系数Tab.3 CorrelationcoefficientbetweenmorphologicaltraitsandgeographicenvironmentalfactlrsofSchimasuperbaariationseeds性状宽长宽比千粒重东经北纬年均温年降雨量无霜期1月均温7月均温长0.691*0.0530.447-0.265-0.4390.636*0.3640.1960.4970.209宽1-0.684*0.526-0.190-0.3660.6290.4140.1780.402-0.080长宽比-0.684*1-0.292-0.0060.073-0.236-0.236-0.042-0.0850.354千粒重0.526-0.2921-0.100-0.730*0.714*0.5700.4750.6190.303
2.3 不同种源木荷种子形态性状的聚类
以亲代特征为指标的地理变异模式与种源试验的结果基本相符。因此,对亲代表型性状进行聚类分析对于早期获得种源区划信息具有重要意义[19]。以木荷种子长、宽、长宽比和千粒重等为指标,采用系统聚类法对10个木荷种源进行种源划分(见图1)。聚类结果为: 木荷种源可聚为3类,湖南攸县、湖南资兴、湖南金洞、湖南桑植、江西分宜为一类;湖南长沙、广西苍梧、广西昭平为一类;湖南汨罗、广西博白为一类。根据聚类结果,湖南攸县、湖南资兴、湖南金洞等3个种源的地理位置接近,广西苍梧、广西昭平的地理位置接近,地理位置相近的聚为了一类;但湖南汨罗、广西博白等地理位置相隔较远,且其年均温度、降水量、无霜期等气候因子也相差较大,但仍然聚为了一类。表明,木荷种子形态特征既存在部分地区区域板块化特征,也存在随机变异的特征。广布种植物地理变异的形成与自然选择作用、基因流作用和基因飘动作用等多种因素相关,在诸多因素综合作用下,植物会形成连续变异、区域板块变异以及随机变异等多种变异模式[19]。
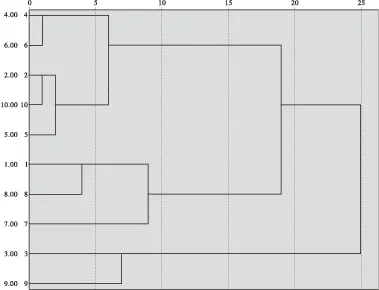
注: 1为湖南长沙;2为湖南攸县;3为湖南汨罗;4为湖南资兴;5为湖南金洞;6为湖南桑植;7为广西苍梧;8为广西昭平;9为广西博白;10为江西分宜。图1 木荷不同种源聚类图Fig.1 Dendrogram of different Schima superba from different provenances
3 结论与讨论
(1) 不同种源木荷的种子长、宽、千粒重等性状差异均达到极显著水平,说明种子形态性状在种源间存在较丰富的遗传变异,具有遗传改良的潜力;种源间种子长、宽、长宽比、千粒重的遗传力分别为0.7340、0.8682、0.6479和0.8165,各性状均受到中等程度以上的遗传控制,为木荷的遗传改良提供了可靠保障。
(2) 木荷种子长度与种子宽度、年均温度均呈显著正相关,说明种子的长度受种子宽度和当地年均温度变化的影响;种子千粒重与北纬呈显著负相关,与年均温度呈显著正相关,说明不同种源地理环境因子中北纬、年均温度的变化对木荷种子的形态变异影响较大。
(3) 10个种源木荷种子形态性状聚类结果,有地理位置相近的种源聚为一类,也有地理位置较远的种源聚为一类,说明木荷种子形态特征呈现地理位置区域板块变异模式和随机变异模式。
(4) 本文仅对10个种源木荷种子形态性状变异进行了分析,要对木荷种源的地理变异模式辨析以及科学的种源区划,还需扩大种源采集点做进一步研究。
[1] 童方平,吴际友.湖南主要乡土树种培育技术[M].中国林业出版社,2015:254-258.
[2] 郭起荣.南方主要树种育苗关键技术[M].中国林业出版社,2011:175-177.
[3] 余本付.安徽省乡土树种造林技术[M].中国林业出版社,2007:89-92.
[4] 王瑞辉.木荷育苗技术研究[J].湖南林业科技,2002,29(3):24-25.
[5] 谢文雷.木荷播种育苗的关键技术 [J].中南林业科技大学学报,2004,24(2):59-63.
[6] 袁冬明,林磊,严春风.木荷轻基质网袋容器育苗技术研究[J].南京林业大学学报,2011,35(6):53-58.
[7] 林沐恩.木荷防火林带营造技术[J].林业科技开发,2004,18(6):71-72.
[8] 刘钦.木荷人工混交林涵养水源的功能[J].福建农林大学学报,2004,33(4):481-484.
[9] 傅祥久.木荷优质干材培育修枝技术的研究[J].河北农业科学,2010,14(5):10-13.
[10] 王秀花,马丽珍,马雪红,等.木荷人工林生长和木材基本密度[J].林业科学,2011,47(7):138-144.
[11] 曾志光,肖复明,包国华,等.木荷不同种源种子品质检验分析[J].江西林业科技,2003(6):1-2.
[12] 王旭军,张日清,许忠坤,等.红榉不同种源种子形态性状变异[J].中南林业科技大学学报,2015,35(1):1-6.
[13] 李晓洁,徐化成.白皮松种子发芽习性及其种源变异的研究[J].林业科学,1989,25(2):97-104.
[14] 宋丽华,王娅丽.几个臭椿种源种子的生物学特性变异研究[J].农业科学研究,2005,26(1):18-22
[15] 续九如.林木数量遗传学[M ].北京:高等教育出版社,2006:34-51
[16]程诗明,顾万春.苦楝表型性状梯度变异的研究[ J].林业科学,2006,42(5):29-35
[17] 王军辉,顾万春,万军,等.桤木不同种源球果及种子性状的遗传变异[J].东北林业大学学报,2006,34(2):1-4.
[18] 黄雪方,金雅琴,李冬林.乌桕不同种源种子性状的地理变异[J].西南林业大学学报,2011,31(4):44-48.
[19] 魏胜利,王文全,秦淑贡,等.甘草种源种子形态与萌发特性的地理变异研究[J].中国中药杂志,2008,33(8):869-872.
SeedmorphologicalvariationofSchimasuperbafromdifferentprovenances
TANG Jie1, TANG Yuxi1, YANG Yan1, LI Yongjin1, WU Min1, LIU Xiu2, ZHONG Wenbing3
(1.Hunan Academy of Forestry,Changsha 410004, China;2.Guangxi Forestry Research Institute, Nanning 530001, China; 3.The Experimental Centre of Subtropical Forestry,Fenyi 336600, China)
The seed character variations of 10Schimasuperbaprovenances were researched, which including length, width, length-width ratio and thousand-seed weight of seed. The results were as follows: There was extremely significant difference between seed length, width and length-width ratio for differentsuperbaprovenances. The difference of seed length-width ratio also reached significant level. And the heritability of these characters on the provenance level was 0.734 0, 0.868 2, 0.647 9 and 0.734 0 separately. The seed length had a significant positive correlations with seed width and annual average temperature. Thousand-seed weight had significant negative correlation with north latitude, but significant positive correlations with annual average temperature. Geographic variation pattern ofS.superbaseed character were regional plate variation pattern and random mutation.
Schimasuperba; provenance, seed; morphological variation
2015-10-08
湖南省林业厅科技计划项目(XLK201502);湖南省林业血防工程建设科技支撑项目。
唐 洁(1980-),男,广西壮族自治区全州县人,助理研究员,主要从事杨树育种与栽培、林业生态工程、木荷种质资源收集与选育等研究工作。
S 685.14
A
1003 — 5710(2015)06 — 0061 — 04
10. 3969/j. issn. 1003 — 5710. 2015. 06. 010
(文字编校:唐效蓉)
