姜黄素对老年鼠脑内氧化性平衡和星形胶质细胞的影响
2015-10-31段雅乐
张 盼, 俞 彬, 刘 莎, 段雅乐, 赵 政
(华东师范大学 脑功能基因组学教育部重点实验室,上海市脑功能基因组学重点实验室,神经药理研究室,上海 200062)
0 引 言
氧化应激是指由体内活性氧(ROS)的产生与清除失衡导致的组织和细胞的氧化损伤.脑组织的氧化损伤是导致认知功能障碍的重要原因.Maugeri研究了氧化应激和衰老之间的关系,结果发现自由基水平和认知功能呈负相关(p<0.000 1)[1].Fukui等将3月龄的 Wistar大鼠连续5 d置于高氧环境中(氧化应激模型),结果大鼠在水迷宫和八臂迷宫任务中表现出记忆能力明显下降,使用Vitamin E(抗氧化剂)则可逆转氧化应激造成的认知损伤[2].
作为哺乳动物脑内分布最广泛的一类细胞,星形胶质细胞是脑内抗氧化系统的重要组成部分,其专属标志酶谷氨酰胺合成酶(GS)在脑内GSH的合成代谢中发挥着重要作用[3],可保护神经元免受ROS诱导的损伤[4];同时星形胶质细胞分泌的神经营养因子(GDNF、BDNF等)对于神经再生起到非常重要的作用[5];此外,星形胶质细胞分泌的D-丝氨酸是诱发LTP的必要条件[6].以上几方面体现了星形胶质细胞对于认知功能的重要性.
大量研究表明,姜黄素具有抗炎、抗氧化的功能[7-8].Wang等利用Aβ诱导的大鼠Alzheimer’s disease模型发现姜黄素可以通过抑制星形胶质细胞的过度活化来改善大鼠的认知损伤[9].但是较长时间的姜黄素处理对正常衰老大鼠脑内的氧化状态及星形胶质细胞的影响还未见报道,这方面的研究对于姜黄素的临床应用具有更加实际的指导意义.
前期研究中分别用姜黄素处理正常衰老的大鼠6周和12周,结果表明,姜黄素处理12周对于正常老年鼠认知功能的改善作用更加明显,较长时间的姜黄素处理可以提高老年鼠的认知功能,此外还发现其机制与姜黄素促进老年鼠的神经发生和对一些功能基因表达的影响有关,其中一部分基因与氧化应激(如Met、Nts、ATP8)和星形胶质细胞功能有关(如Slc38a4,参与谷氨酸谷氨酰胺循环)[10].本研究观察姜黄素处理12周对老年鼠脑内氧化性平衡和星形胶质细胞(形态、营养因子、谷氨酰胺合成酶和D-丝氨酸等)的影响,为姜黄素改善老年鼠认知功能提供分子依据.
1 材料与方法
1.1 材料
1.1.1 实验动物
雄性SD大鼠,购自上海西普尔-必凯实验动物有限公司.本实验分3组:年轻对照组(Young control)(2月龄SD大鼠),对照组(Control)和姜黄素处理组(Curcumin)(均为18月龄SD大鼠).姜黄素处理组给予姜黄素处理12周,其他两组给予正常饲料.大鼠正常饲料与实验饲料(姜黄素含量为480 mg/kg)均购自苏州双狮实验动物饲料科技有限公司.姜黄素来自于SanjivaniPhytopharma Pvt.Ltd,India[10].根据大鼠摄食量测量结果,大鼠日平均摄食量约为25 g,因此预计实验组大鼠每日摄入姜黄素为12 mg/只.每组15只大鼠,其中免疫荧光实验每组9只,其他实验每组6只.本动物实验方法和目的符合人类的道德伦理标准和国际惯例,并且经由华东师范大学动物实验伦理委员会批准(AR201404022),在动物实验期间遵守有关的法规、实验动物伦理福利原则.
1.1.2 实验仪器
冰冻切片机(Leica),荧光显微镜(NIKON),Spectra Max M5多功能酶标仪(Molecular Devices公司),电泳凝胶成像仪(Bio-rad公司),micrOTOFQⅡ型质谱仪(德国Bruker Dal-tonics公司),HPLC(Agilent Technologies 1200).
1.1.3 试剂
GFAP抗体(Abcam),GS抗体(Bioworld),GAPDH 抗体(康城生物),RIPA裂解液、Bradford蛋白浓度测定试剂盒、SDS-PAGE凝胶配制试剂盒和超敏ECL化学发光试剂盒(碧云天),丙二醛(MDA)、8-羟基脱氧鸟苷(8-OHdG)、还原型谷胱甘肽(GSH)、γ-谷氨酰半胱氨酸合成酶(GCS)和谷氨酰胺合成酶(GS)试剂盒(南京建成),BDNF Elisa和GDNF Elisa试剂盒(ShanghaiWanLang Bio-TechnoloqyCa.,Led),D-丝氨酸和 Marfey试剂(Sigma).
1.2 实验方法
1.2.1 Western Blot实验
处理12周后的老年鼠断头处死,迅速剥离脑组织,在冰上分离出皮层和海马,液氮中速冻.每20 mg组织加入200μL RIPA(PMSF浓度为1 mmol/L)裂解液,充分裂解后14 000 g离心5 min,取上清,测定蛋白浓度.跑胶条件:5%浓缩胶,10%分离胶,恒压80 V跑到底.转膜条件:PVDF膜0.2 A恒流55 min.转完膜后5%脱脂牛奶室温封闭1 h,TBST洗膜(摇床3×5 min),4℃过夜孵育一抗,二抗室温孵育1 h,TBST洗膜(摇床3×5 min)后上机检测.
1.2.2 酶活性试验
依据试剂盒说明书依次添加各反应成分,MDA于532 nm处检测各组吸光值,GSH于420 nm处检测各组吸光值,GCS于340 nm处检测各组吸光值,GS于412 nm处检测各组吸光值,8-OhdG、BDNF和GDNF采用双抗体夹心ELISA法进行检测,450 nm处检测各组吸光值.
1.2.3 免疫荧光实验
1.2.3.1 脑灌注固定
处理12周后老年鼠腹腔注射戊巴比妥(50 mg/kg),将针头从心尖部插入左心室,生理盐水(37℃预热)200 mL迅速冲净脑内血液,4℃的4%多聚甲醛灌注结束后断头并分离脑组织.将取出的脑组织置于4%多聚甲醛溶液中固定4 h,然后用蔗糖溶液梯度脱水至脑组织沉降至瓶底.
1.2.3.2 冰冻切片
将脑组织从蔗糖中取出,平稳放置于载物台的包埋剂上并迅速放置于切片机的冷冻台上快速冷冻至脑组织变白.将包埋冷冻好的脑组织固定于切片机的标本台上,刀片厚度为30μm,切片晾干后于37℃烘箱中烘烤过夜.
1.2.3.3 免疫荧光
将-80℃保存的切片取出,室温放置10 min.用0.1%Triton X-100的PBS溶液漂洗5 min.室温封闭(5%山羊血清,3%Triton,3%BSA in PBS)2 h.一抗4℃孵育过夜,二抗37℃孵育30 min.PBS清洗干净,封片.
1.2.4 高效液相色谱-质谱(LC-MS)检测皮层和海马的D-丝氨酸
D-丝氨酸测定按本实验室建立的LC-MS法进行[11].大鼠脑组织样品匀浆离心后,取上清液并加入Marfey试剂对样品中L/D-丝氨酸进行衍生化处理.HPLC为Agilent Technologies 1 200,色谱柱采用Agilent,ZORBAXSB-C18,(150 mm×2.1 mm,5μm),流动相A为0.025%甲酸铵(pH 用甲酸调至4.6),B为乙腈.洗脱程序;0~15 min,5%~50%B;15~19 min,50%~5%B;19~25 min,5%B.质谱采用电喷雾离子源(ESI),负离子模式,在m/z 50~500范围内进行全离子扫描,并选择m/z为356.1的分子离子峰用于定量.
1.2.5 数据统计
应用SPSS 10.0软件,组间比较采用LSD或 Mann-Whitney U test,结果用mean±SEM 表示.*,#p<0.05;**,##p<0.01;***,###p<0.001.
2 结果
2.1 姜黄素对老年鼠脑内氧化性平衡的影响
与年轻对照组相比,对照组老年鼠脑内MDA和8-OHdG含量明显增加,GSH水平明显减少,GCS活性明显降低.经过姜黄素处理的老年鼠脑内MDA和8-OHdG含量明显减少(见图1 A,B),GSH的水平大幅度提升(见图1 C),GCS的活性明显升高(见图1 D).

图1 姜黄素对MDA、8-OHdG和GSH的水平与GCS活性的影响Fig.1 The effect of curcumin on MDA,8-OHdG,GSH levels and GCS activity
2.2 姜黄素对老年鼠星形胶质细胞的影响
与年轻对照组相比,在对照组老年鼠皮层、海马DG区和CA1区的星形胶质细胞过度活化,突起变短,有突破折样表现,给予姜黄素后形态明显改善,细胞突起变得细长,胞体也有所减小(见图2);GFAP在对照组老年鼠脑内表达增加,反映了星形胶质细胞的过度活化状态,而姜黄素的处理可明显降低老年鼠皮层(见图3 A,B)中GFAP的表达,在海马虽然没有统计学差异,但是也能观察到GFAP有下降趋势(见图3 C,D,p=0.058),表明姜黄素抑制了星形胶质细胞的过度活化.
2.3 姜黄素对老年鼠星形胶质细胞功能的影响
皮层(见图4 A,C)和海马(见图4 B,D)中GS蛋白表达在3组之间没有差异.酶活性实验结果表明与年轻对照组相比,GS活性在老年鼠中明显减低,而姜黄素则可明显增加其活性(见图4 E);BDNF和GDNF的含量在老年鼠中降低,姜黄素对BDNF的含量没有影响(见图5 A),但是可明显增加老年鼠皮层和海马中GDNF的含量(见图5 B);老年鼠皮层和海马中的D-丝氨酸明显降低,姜黄素可增加皮层中D-丝氨酸的含量,海马中的D-丝氨酸则有上升的趋势(见图6).
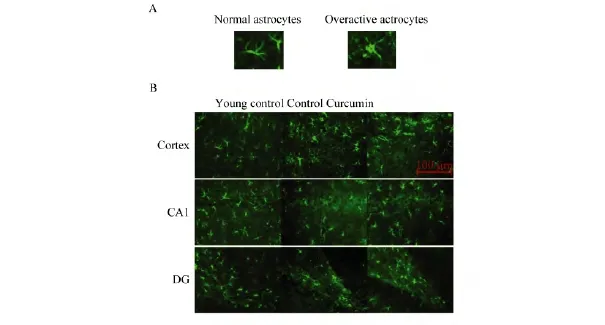
图2 姜黄素对星形胶质细胞形态的改善作用Fig.2 The effect of curcumin on the improvement of the morphology of astrocytes
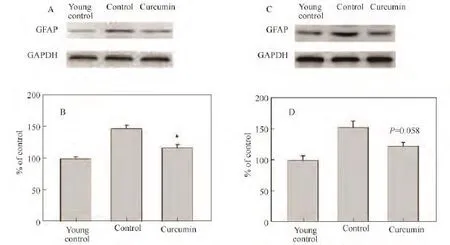
图3 老年鼠GFAP蛋白的表达Fig.3 Expression of GFAP in aged rats detected by western
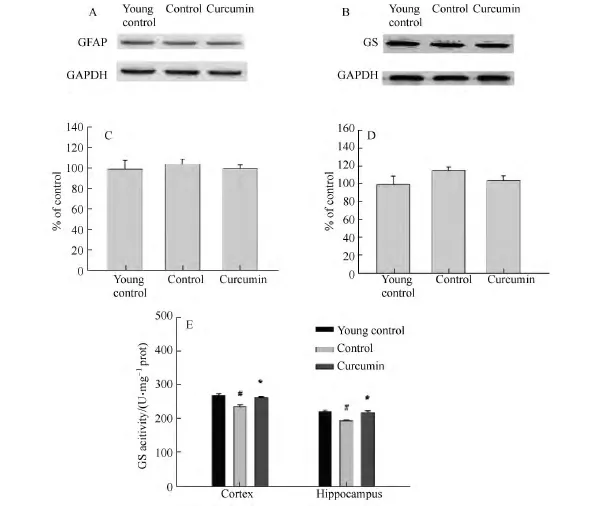
图4 姜黄素对GS的影响Fig.4 The effect of curcumin on GS

图5 姜黄素对BDNF和GDNF水平的影响Fig.5 The effect of curcumin on BDNF and GDNF levels

图6 姜黄素对老年鼠皮层和海马中D-丝氨酸含量的影响Fig.6 The effect of curcumin on the content of D-serine in the cortex and hippocampus of aged rats
3 结 论
在脑内GSH既可以阻断毒性化合物对DNA、RNA及蛋白质的损害,又可以作为重要的还原剂,保护体内蛋白质或酶分子中的巯基免遭氧化,还可参与蛋白质合成、细胞免疫和酶促反应[12-13].脑内的GSH随着年龄的增加进行性减少,并且这种减少可以导致老年认知功能障碍[14].本实验结果表明,长期摄入姜黄素可以改善正常老年鼠脑内的氧化应激状态:降低皮层和海马内氧化产物MDA和8-OHdG的含量(见图1 A,B),提高GCS活性(GCS是GSH合成反应中的限速酶,决定着GSH生物合成的速率和量)(见图1 D),并增加脑内的GSH水平(见图1 C),这表明姜黄素通过GCS和GSH起到了抗氧化的作用.
氧化应激是大脑衰老进程中的一个关键性的损害因素,虽然星形胶质细胞内含有很多抗氧化物质并发挥保护神经元的作用,但是星形胶质细胞自身也会受到氧化应激的影响而发生一系列形态和功能的变化.星形胶质细胞的形态随着年龄的增长呈现过度活化的状态,此时胞体变的肥大,突起增多,严重时有突破折样表现[15-17],其标记物GFAP在脑内绝大部分区域的表达都会增加[18-19].结果显示,姜黄素可以抑制老年鼠星形胶质细胞的过度活化(见图2),降低其标记物GFAP的蛋白过度表达(见图3).有研究表明,GSH可以抑制神经毒性物质导致的星形胶质细胞过度活化[20],而减轻氧化应激和抑制GFAP表达可以进一步改善AD大鼠的认知功能[21].因此认为姜黄素通过诱导脑内GSH的产生,减轻了老年鼠皮层和海马的氧化损伤,这些效应有可能与姜黄素抑制星形胶质细胞的过度活化和改善星形胶质细胞的功能有关.
星形胶质细胞在脑内GSH的合成代谢中发挥着重要的作用[3]:谷氨酰胺合成酶(GS)是星形胶质细胞专属标志酶,参与谷氨酸谷氨酰胺循环,对于神经递质的代谢具有重要作用,星形胶质细胞吸收谷氨酸和γ-氨基丁酸,在胞体内由GS催化合成谷氨酰胺,合成的谷氨酰胺被神经元吸收可以脱氨参与到GSH的形成中去.研究表明GS酶活性降低可以导致脑内GSH水平下降[22],随着年龄的增加星形胶质细胞的GS酶活性明显下降[23],但是其基因和蛋白表达水平未见改变[24].在研究结果中,老年鼠脑内的GS蛋白表达未见改变,而酶活性却明显降低,与相关报道一致.姜黄素处理虽然对GS蛋白表达没有影响,但是增强了GS酶的活性(见图4),由此可见,长期的姜黄素治疗可能通过改善星形胶质细胞功能从而促进脑内GSH的合成,产生氧化应激的良性循环,进一步起到对老年鼠认知功能的改善作用.
星形胶质细胞参与构成“干细胞微环境”,其分泌的神经营养因子包括GDNF、BDNF等可促进神经发生[25-26].在实验中,姜黄素增加了GDNF的水平(见图5 B),但对BDNF没有影响(见图5 A).这提示姜黄素可能通过促进GDNF的分泌来诱导神经发生.D-丝氨酸在体内是由丝氨酸消旋酶将L-丝氨酸转化而来的[27],主要由星形胶质细胞合成释放,其脑内分布与 N-甲基-D-天门冬氨酸(N-methyl-d-aspartate,NMDA)受体一致[28].D-丝氨酸作为NMDA受体的内源性配体,能激活NMDA受体的甘氨酸结合位点,为突触提供营养、支持以及传递调控,有助于突触的形成,并参与学习和记忆等生理活动[29-30].而衰老时D-丝氨酸的含量和合成功能明显下降,导致NMDA受体依赖的突触可塑性下降,并出现认知损伤[31].有研究表明长期喂食抗氧化剂L-NAC可以改善氧化还原状态,增加D-丝氨酸及NMDA受体依赖的突触可塑性[32],但是姜黄素与D-丝氨酸的关系却未见报道.在本文结果中,老年鼠脑内的D-丝氨酸含量明显降低,而姜黄素提高了老年鼠脑内D-丝氨酸的含量(见图6).这提示姜黄素可能通过调控星形胶质细胞对D-丝氨酸的合成释放改善了老年鼠的认知能力.
[1] MAUGERI D,SANTANGELO A,BONANNO M R,et al.Oxidative stress and aging:Studies on an East-Sicilian,ultraoctagenarian population living in institutes or at home[J].Arch Gerontol Geriatr Suppl,2004(9):271-277.
[2] FUKUI K,OMOI N O,HAYASAKA T,et al.Cognitive impairment of rats caused by oxidative stress and aging,and its prevention by vitamin E[J].Ann N Y AcadSci,2002,959:275-284.
[3] ASCHNER M.Neuron-astrocyte interactions:Implications for cellular energetics and antioxidant levels[J].Neurotoxicology,2000,21(6):1101-1107.
[4] FEMANDEZ-FEMANDEZ S,ALMEIDA A,BOLANOS J P.Antioxidant and bioenergetic coupling between neurons and astrocytes[J].Biochem J,2012,443(1):3-11.
[5] GORDON T.The physiology of neural injury and regeneration:The role of neurotrophicfactors[J].J Commun Disord,2010,43(4):265-273.
[6] YANG Y,GE W,CHEN Y,et al.Contribution of astrocytes to hippocampal long-term potentiation through release of D-serine[J].Proc Natl Acad Sci USA,2003,100(25):15194-15199.
[7] CHHUNCHHA B,FATMA N,KUBO E,et al.Curcumin abates hypoxia-induced oxidative stress based-ER stress-mediated cell death in mouse hippocampal cells(HT22)by controlling Prdx6 and NF-kappa Bregulation[J].Am J Physiol Cell Physiol,2013,304(7):C636-C655.
[8] SOETIKNO V,SARI F R,LAKSHMANANAK A P,et al.Curcumin alleviates oxidative stress,inflammation,and renal fibrosis in remnant kidney through the Nrf2-keap1 pathway[J].Mol Nutr Food Res,2013,57(9):1649-1659.
[9] WANG Y,YIN H,WANG L,et al.Curcumin as a potential treatment for Alzheimer’s disease:A study of the effects of curcumin on hippocampal expression of glial fibrillary acidic protein[J].Am J Chin Med,2013,41(1):59-70.
[10] DONG S,ZENG Q,MITCHELL E S,et al.Curcumin enhances neurogenesis and cognition in aged rats:Implications for transcriptional interactions related to growth and synaptic plasticity[J].Plos One,2012,7(2):e31211.
[11] 王伟莉.HPLC-ESI-MS法测定大鼠脑组织中神经递质L/D-丝氨酸的含量[J].华东师范大学学报:自然科学版,2013(6):165-170.
[12] AOYAMA K,WATABE M,NAKAKI T.Regulation of neuronal glutathione synthesis[J].J Pharmacol Sci,2008,108(3):227-238.
[13] DRINGEN R,HIRRLINGER J.Glutathione pathways in the brain[J].Biol Chem,2003,384(4):505-516.
[14] CURRAIS A,MAHER P.Functional consequences of age-dependent changes in glutathione status in the brain[J].Antioxid Redox Signal,2013,19(8):813-822.
[15] CERBAI F,LANA D,NOSI D,et al.The neuron-astrocyte-microglia triad in normal brain ageing and in a model of neuroinflammation in the rat hippocampus[J].Plos One,2012,7(9):e45250.
[16] BERCIANO M T,ANDRES M A,CALLE E,et al.Age-induced hypertrophy of astrocytes in rat supraoptic nucleus:A cytological,morphometric,and immunocytochemicalstudy[J].Anat Rec,1995,243(1):129-144.
[17] ITOH Y,YAMADA M,SUEMATSU N,et al.An immunohistochemical study of centenarian brains:Acomparison[J].J Neurol Sci,1998,157(1):73-81.
[18] ZHANG R,KADAR T,SIRIMANNE E,et al.Age-related memory decline is associated with vascular and microglial degeneration in aged rats[J].Behav Brain Res,2012,235(2):210-217.
[19] LIVELY S,SCHLICHTER L C.Age-related comparisons of evolution of the inflammatory response after intracerebral hemorrhage in rats[J].Transl Stroke Res,2012,3(Suppl 1):132-146.
[20] ENONGENE E N,SUN P N,MEHTA C S.Sodium thiosulfate protects against acrylonitrile-induced elevation of glial fibrillary acidic protein levels by replenishing glutathione[J].Environ Toxicol Pharmacol,2000,8(2):153-161.
[21] JAVED H,KHAN M M,AHMAD A,et al.Rutin prevents cognitive impairments by ameliorating oxidative stress and neuroinflammation in rat model of sporadic dementia of Alzheimer type[J].Neuroscience,2012,210:340-352.
[22] GHODDOUSSI F,GALLOWAY M P,JAMBEKAR A,et al.Methionine sulfoximine,an inhibitor of glutamine synthetase,lowers brain glutamine and glutamate in a mouse model of ALS[J].J Neurol Sci,2010,290(1-2):41-47.
[23] HEAD E,LIU J,HAGEN T M,et al.Oxidative damage increases with age in a canine model of human brain aging[J].J Neurochem,2002,82(2):375-381.
[24] WU Y,ZHANG A Q,YEW D T.Age related changes of various markers of astrocytes in senescence-accelerated mice hippocampus[J].Neurochem Int,2005,46(7):565-574.
[25] ISLAM O,LOO T X,HEESE K.Brain-derived neurotrophic factor(BDNF)has proliferative effects on neural stem cells through the truncated TRK-B receptor,MAP kinase,AKT,and STAT-3 signaling pathways[J].Curr Neurovasc Res,2009,6(1):42-53.
[26] CHEN Y,AI Y,SLEVIN J R,et al.Progenitor proliferation in the adult hippocampus and substantianigra induced by glial cell line-derived neurotrophic factor[J].Exp Neurol,2005,196(1):87-95.
[27] SCHELL M J.The N-methyl D-aspartate receptor glycine site and D-serine metabolism:An evolutionary perspective[J].Philos Trans R Soc Lond B Biol Sci,2004,359(1446):943-964.
[28] BILLARD J M.D-serine signalling as a prominent determinant of neuronal-glial dialogue in the healthy and diseased brain[J].J Cell Mol Med,2008,12(5B):1872-1884.
[29] NAGAI T,YU J,KITAHARA,et al.D-serine ameliorates neonatal PolyI:C treatment-induced emotional and cognitive impairments in adult mice[J].J Pharmacol Sci,2012,120(3):213-227.
[30] FOSSAT P,TURPIN F R,SACCHU S,et al.Glial D-serine gates NMDA receptors at excitatory synapses in prefrontal cortex[J].Cereb Cortex,2012,22(3):595-606.
[31] TURPIN F R,POTIER B,DULONG J R,et al.Reduced serine racemase expression contributes to age-related deficits in hippocampal cognitive function[J].Neurobiol Aging,2011,32(8):1495-1504.
[32] HAXAIRE C,TURPIN F R,POTIER B,et al.Reversal of age-related oxidative stress prevents hippocampal synaptic plasticity deficits by protecting D-serine-dependent NMDA receptor activation[J].Aging Cell,2012,11(2):336-344.
