LYC-B基因沉默对紫色番茄果实主要色素及挥发性物质的影响
2015-10-29赵菁菁常培培李云洲
吕 洁,梁 燕*,赵菁菁,张 颜,常培培,秦 蕾,李云洲
(西北农林科技大学园艺学院,陕西 杨凌 712100)
LYC-B基因沉默对紫色番茄果实主要色素及挥发性物质的影响
吕洁,梁燕*,赵菁菁,张颜,常培培,秦蕾,李云洲
(西北农林科技大学园艺学院,陕西 杨凌712100)
利用病毒诱导的基因沉默(virus-induced gene silencing,VIGS)技术,抑制紫色番茄果实成熟过程中番茄红素β-环化酶(LYC-B)基因的表达,并分析番茄果实中主要色素及挥发性物质的种类与含量变化。结果表明:LYC-B基因沉默可提高紫色番茄果实中番茄红素的含量;顶空固相微萃取-气相色谱-质谱联用技术检测到6-甲基-5-庚烯-2-酮等类胡萝卜素相关的挥发性物质的含量增加,反-2-辛烯醛、己醇、反-2-庚烯醛、水杨酸甲 酯和顺-3-己烯醇等主要挥发性物质的释放量增加。因此,LYC-B基因沉默可以增加番茄红素及部分主要挥发性物质的含量,影响番茄果实的营养品质和风味品质。
番茄;LYC-B基因沉默;番茄红素;挥发性物质
番茄(Solanum lycopersicum L.)具有丰富的营养物质,如类胡萝卜素、维生素、氨基酸等,并因其风味多样而深受消费者青睐。类胡萝卜素是植物天然色素的总称,在预防人类疾病和维持健康方面有重要作用[1]。类胡萝卜素也是一些重要挥发性物质的前体,这些物质为番茄果实提供特殊的香气,决定着番茄果实的风味品质[2]。类胡萝卜素降解途径是形成番茄果实风味品质的最重要途径[3],其含量直接影响着类胡萝卜素衍生挥发性物质的释放[4],例如6-甲基-5-庚烯-2-酮、牻牛儿基丙酮、β-紫罗兰酮及相关成分均来自类胡萝卜素的氧化分解[2]。番茄红素作为主要的类胡萝卜素,对番茄果实风味物质形成具有重要的贡献[5]。Fay等[6]发现番茄红素特效裂解过氧化酶BoLCD可以裂解番茄红素形成6-甲基-5-庚烯-2-酮,可提高番茄果实的风味品质。
番茄红素β-环化酶(LYC-B)基因是类胡萝卜素代谢中番茄红素向下游转化的关键酶基因[7],可以催化番茄红素分子的两个末端形成β-环,即形成β-胡萝卜素。徐加新等[8]研究表明,反义LYC-B基因可以提高番茄果实中番茄红素的含量。目前对LYC-B基因与其他色素及挥发性物质的关系研究较少。
紫色番茄作为新兴的番茄品种,具有极强的抗氧化性等营养保健功能,风味独特[9]。本实验室常培培等[10]对其果实挥发性物质进行了系统全面的分析鉴定,本实验将在此基础上,利用病毒诱导的基因沉默技术[11-12],探究类胡萝卜素代谢途径中关键酶基因LYC-B与紫色番茄果实中主要色素及挥发性物质间的关系,为番茄品质性状研究与改良提供理论依据和实践基础。
1 材料与方法
1.1材料、菌种与试剂
供试紫色番茄品种为黑樱桃(保存单位编号:CI5004),果皮无色,果肉紫红色,由西北农林科技大学园艺学院番茄育种课题组提供。2014年3月底定植于园艺学院实验农场塑料大棚内,当植株第三花序第一、二朵花开放时挂牌标记开花期。在果实绿熟期[13](开花后30 d,此时果实充分长大,果色绿色,肉质坚硬)采收果实,离体条件下进行实验。
农杆菌GV3101、pTRV1、pTRV2和pTRV2:PDS等由园艺学院番茄育种课题组保存提供。
内切酶、连接酶Fermentas公司;反转录试剂盒大连宝生物工程公司;2-壬酮(色谱纯)上海迈瑞尔公司;氯化钠(分析纯)、番茄红素标品(色谱纯)北京中科仪友化工技术研究院。
1.2仪器与设备
聚合酶链式反应(polymerase chain reaction,PCR)扩增仪、IQ5实时荧光定量PCR扩增仪美国Bio-Rad公司;LC-20A高效液相色谱日本岛津公司;ISQ气相色谱-质联用仪(Xcalibur1.2数据处理系统)美国Thermo Fisher Scientific公司;HP-INNOWAX弹性石英毛细管柱(60 m×0.25 mm,0.25 .m)、恒温磁力搅拌器美国Troemner公司;100 μm PDMS固相微萃取头、SPME手动进样手柄美国Supelco公司;匀浆机美的集团。
1.3方法
1.3.1VIGS病毒载体pTRV2:LYC-B的构建
VIGS诱导基因沉默是指携带目的基因片段的pTRV2与pTRV1混合后浸染植物后,可诱导植物内源目的基因沉默。根据美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI)上登录的番茄红素β-环化酶(登录号为X86452.1)序列,选取3'端375 bp特异性较强的片段,添加BamHⅠ和XbaⅠ酶切位点,共392 bp,设计引物为LYC-B F:5'-CGC GGA TCC CAT CAC TCG TAG CTC GTC-3';LYC-B R:5'-GCT CTA GAT ATC CAT ACC GAA GCA GA-3'。载体构建过程参照李翠[14]的方法。
1.3.2VIGS诱导番茄果实LYC-B基因沉默
处理设置:以注射pTRV2:LYC-B菌液的果实为沉默LYC-B基因的处理,pTRV2:00为阴性对照,pTRV2:PDS为阳性对照,未注射菌液为空白对照。
番茄果实注射浸染:参照Orzaez等[15]的方法对果实进行注射处理。将含有pTRV2:LYC-B、pTRV2:00和pTRV2:PDS质粒的菌液分别在LB筛选培养基中振荡过夜,取培养产物以1∶25的比例稀释于诱导培养基中,培养至OD600nm为0.5~0.8。培养产物离心后,使用悬浮培养基MgCl2+MES(10 mmol/L,pH 5.5)等体积悬浮菌体,再次离心后,重悬菌体并调节OD600nm为1.0。同样方法准备pTRV1菌液,室温下放置3 h后,将pTRV1分别与pTRV2:LYC-B、pTRV2:00和pTRV2:PDS菌液混合注射离体果实。使用1 mL注射器从果顶注射果实,当有菌液从萼片处溢出时,停止注射,用吸水纸擦干溢出的菌液。每个处理重复3 次,每次20 个果实。注射后的果实放置在人工气候箱中,温度(20±2)℃,相对湿度70%,光周期为16 h昼长及8 h夜长。
取样分析:八氢番茄红素脱氢酶基因(phytoene desaturase,PDS)是VIGS诱导中的报告基因,抑制PDS基因表达会使果实不能正常转色,果实表面出现绿色区域或黄色区域[12]。在注射果实2 周后,PDS基因阳性对照组果实的沉默面积不再发生变化时,PDS基因沉默效率达到最高[11],此时其他处理中相应基因的沉默效率也达到最高。参照PDS基因沉默的部位,提取其他处理果实相同部位的RNA,反转录为cDNA。以cDNA为模板,EF1α为内参,进行实时荧光定量PCR分析,检测沉默部位LYC-B基因的表达量。分别将各处理中LYC-B基因表达量低的部位用匀浆机打成匀浆,液氮冷冻后保存在-80 ℃,用于后续实验的测定。
1.3.3指标测定
参照刘沐霖等[16]采用高效液相色谱法,测定VIGS诱导果实番茄红素及β-胡萝卜素的含量。
参照Viljanen[17]、常培培[10]等采用顶空固相微萃取-气相色谱-质谱(headspace solid phase micro-extraction gas chromatography-mass spectrometry,HS-SPME-GC-MS)联用技术,测定VIGS诱导果实挥发性物质。
样品制备:称取VIGS处理后果肉匀浆约14 g,加入3 g氯化钠,于40 mL顶空瓶中。将顶空瓶置于50 ℃恒温磁力搅拌器上,平衡10 min,顶空吸附40 min,进样口处解吸3 min,进行GC-MS分析。每个处理重复3 次取平均值。
色谱条件:色谱柱为HP-INNOWAX 弹性石英毛细管柱(60 m×0.25 mm,0.25 .m),进样口250 ℃;升温程序为40 ℃保持2.5 min,10 ℃/min升至110 ℃,然后以3.0 ℃/min升温至180 ℃,15.0 ℃/min 升温至230 ℃维持3 min;载气(He)流速1.0 mL/min,不分流进样。
质谱条件:电子轰击(electron ionization,EI)离子源;电子能量70 eV;离子源温度250 ℃;SM离子扫描,质量扫描范围为m/z 35~500。
1.4数据分析
用Excel 2013软件进行数据处理;用Origin 8.0软件进行作图;用SPSS 20.0统计分析软件进行Tukey's显著性检验。
2 结果与分析
2.1VIGS病毒载体pTRV2:LYC-B的构建
采取黑樱桃番茄幼苗新叶,使用TRIZOL试剂提取总RNA,反转录为cDNA。以cDNA为模板,使用引物LYC-B F和LYC-B R进行PCR扩增,得到预期的392 bp的特异序列(图1A)。该片段与pMD-18T载体连接,测序验证后,与TRV2病毒载体连接,转化农杆菌,进行菌液PCR和双酶切验证。
由图1B、1C可知,菌液PCR扩增和双酶切均得到392 bp的片段,与克隆的LYC-B片段大小相符,表明成功构建了VIGS病毒pTRV2:LYC-B载体,并获得了含有pTRV2:LYC-B质粒的阳性单克隆农杆菌。

图1 重组质粒pTRVV22:LLYYCC--BB的克隆及鉴定Fig.1 Cloneing and identification of pTRV2:LYC-B recombinant cloning vector
2.2VIGS诱导番茄果实LYC-B基因沉默
绿熟期离体番茄果实注射菌液2 周后,阳性对照组的果实出现PDS基因沉默症状,果实表面出现黄绿色区域(图2D),说明此时的VIGS诱导效果最大[12]。通过观察果实颜色变化情况发现,LYC-B基因沉默处理,阴性对照以及空白对照的果实外观没有明显区别,都转色均匀(图2B、2C、2E)。

图2 VIGS诱导番茄果实颜色变化Fig.2 Color change of tomato fruits induced by VIGS
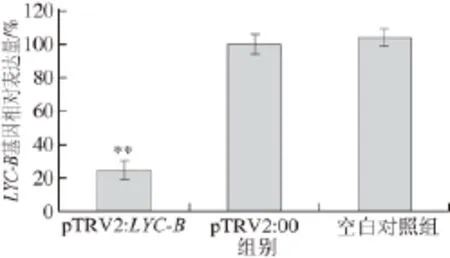
图3 VIGS果实中LYC-BLYC-B基因表达分析Fig.3 Transcript levels of LYC-B gene in gene-silenced tomato fruits
实时荧光定量PCR检测LYC-B基因的表达量。如图3所示,LYC-B基因沉默处理果实中,LYC-B的转录水平比阴性对照组降低了约75%。结果表明,虽然用肉眼观察不到果实颜色明显的变化,但是VIGS诱导极显著降低了果实内源LYC-B基因的表达量,阴性对照组与空白对照组的表达量无显著差异。
2.3LYC-B基因沉默对番茄果实番茄红素和β-胡萝卜素含量的影响
利用高效液相色谱法检测沉默果实和对照处理中番茄红素和β-胡萝卜素的含量。由图4可知,各处理组之间番茄红素含量差异显著,LYC-B基因沉默的果实中番茄红素含量最高,为12.27 mg/100 g,显著高于两个对照组(阴性对照组和空白对照组),与阴性对照组相比增加了14.03%,与空白对照组相比增加了29.15%。LYC-B基因沉默的果实中β-胡萝卜素含量显著高于阴性对照组和空白对照组,阴性对照组和空白对照组二者之间差异不显著。说明LYC-B基因沉默提高了果实中番茄红素和β-胡萝卜素的含量。

图 44 LLYYCC--BB基因沉默对番茄果实番茄红素(AA) 和β--胡萝卜素含量(BB)的影响Fig.4 Contents of lycopene (A) and β-carotene (B) in LYC-B gene silenced tomato fruits
2.4LYC-B基因沉默对番茄果实挥发性物质的影响
利用HS-SPME-GC-MS方法检测沉默果实中挥发性物质的变化情况。检测结果经Xcalibur软件处理,与图谱库(NIST2011)的标准质谱图对照,并结合有关文献的标准图谱进行匹配[18-20],共检测到了52 种正反匹配度大于800的化合物。运用峰面积归一法,求得各物质的相对含量,并按照化合物官能团对物质进行分类[21],结果见表1。
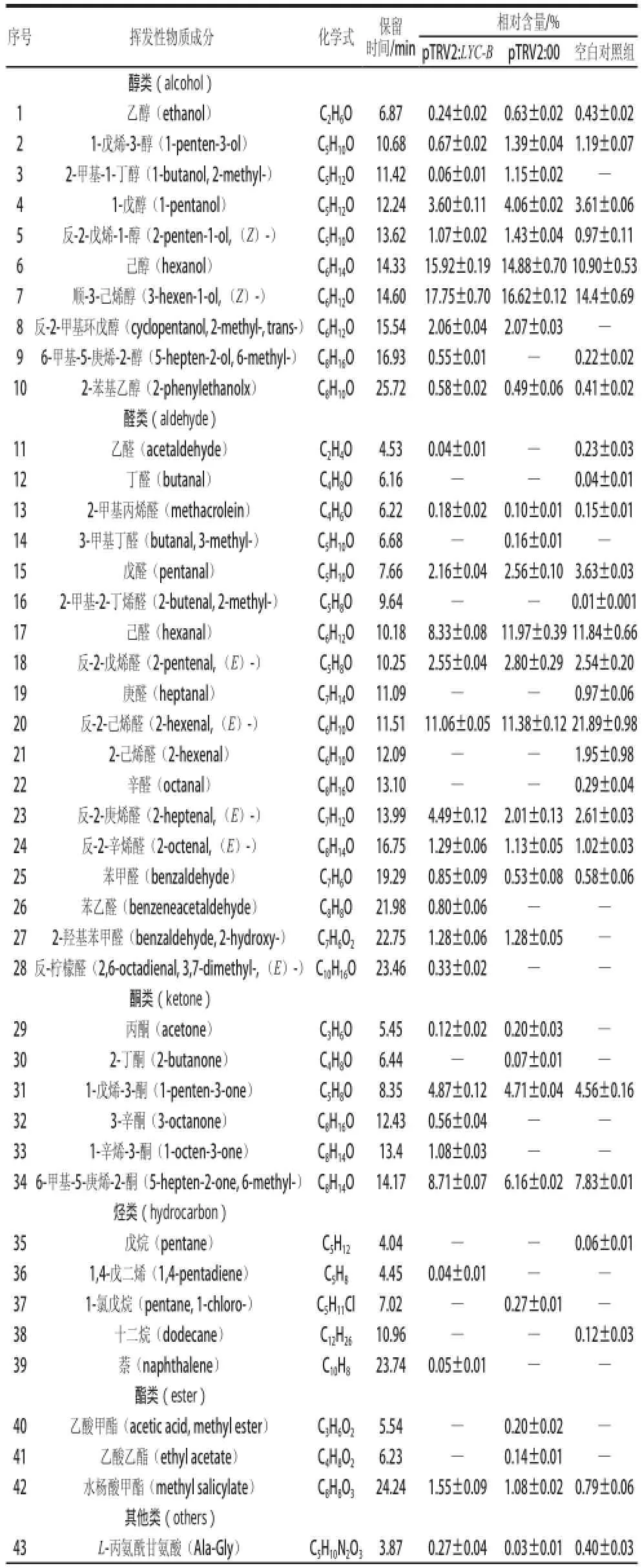
表1 LYC-B基因沉默番茄果实挥发性物质的相对含量Table 1 Relative contents of volatile compounds in LYC-B genesilenced tomato fruits

续表1
2.4.1LYC-B基因沉默对番茄果实挥发性物质种类的影响
由表1可知,所有处理中共检测到52 种挥发性物质,包括醇类10 种、醛类18 种、酮类6 种、烃类5 种、酯类3 种和其他类10 种。
在LYC-B基因沉默的处理组中,检测到37 种物质,与阴性对照组相比,醇类、酮类、烃类和其他类的个数均增加了1 种,醛类增加了2 种,酯类减少了1 种。检测到了苯乙醛、6-甲基-5-庚烯-2-醇、萘和反-柠檬醛等阴性对照组中没有检测到的物质,没有检出2-己烯醛和庚醛等阴性对照组中存在的物质。结果表明,LYC-B基因沉默增加了挥发性物质的种类,改变了挥发性物质的组成。
阴性对照组中酮类和酯类的个数均比空白对照组多2 种,醛类数量比空白对照减少4 种,二者间醇类、烃类和其他类数量差别不大,挥发性物质总数一致。
2.4.2LYC-B基因沉默对番茄果实挥发性物质相对含量的影响
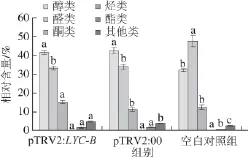
图5 5 LYC-BLYC-B基因沉默番茄果实各类挥发性物质的相对含量Fig.5 Relative contents of volatile compounds in LYC-B gene-silenced tomato fruits
由图5可知,LYC-B基因沉默处理组与阴性对照组相比,酮类相对含量(15.34%)增加了37.7%(阴性对照组为11.14%),其他类相对含量(4.76%)增加了27.6%(阴性对照组为3.73%),而醇类、醛类、烃类和酯类相对含量均没有显著变化。结果表明,LYC-B基因沉默显著增加了酮类物质和其他类物质的相对含量,对剩余物质含量影响不大。
阴性对照组中醇类、酯类和其他类物质的相对含量高于空白对照组,醛类相对含量低于空白对照组,酮类和烃类的相对含量与空白对照组间差异不显著(图5)。由此可见,注射空载体的菌液对挥发性物质的含量造成了影响,推测可能是由于注射过程对果实造成了伤害或病毒空载体本身的作用,从而导致了挥发性物质的变化。
此外,根据番茄风味特征性物质以及对番茄风味品质贡献较大的物质[3,17,20],对LYC-B基因沉默后番茄主要挥发性物质变化情况进行了分析(表1,图6)。在LYC-B基因沉默后,某些主要挥发性物质含量发生了明显变化,如反-2-庚烯醛、6-甲基-5-庚烯-2-酮、水杨酸甲酯、反-2-辛烯醛、己醇和顺-3-己烯醇等的含量显著增加,其中,与阴性对照相比,反-2-庚烯醛的含量增加了2.48 倍,增幅最大;水杨酸甲酯增加了43.51%;6-甲基-5-庚烯-2-酮含量提高了41.40%;反-2-辛烯醛增加了14.16%;己醇和顺-3-己烯醇分别增加了约7%。结果表明LYC-B基因沉默显著(P<0.05)或极显著(P<0.01)增加了部分主要挥发性物质的含量,促进了挥发性物质的释放。

图6 6 LYC-BLYC-B基因沉默番茄果实主要挥发性物质的相对含量Fig.6 Relative contents of the major volatile compounds in LYC-B gene-silenced tomato fruits
3 讨 论
LYC-B基因表达的下调可阻碍番茄红素向具环类胡萝卜素的转化,使番茄红素的分解受阻,从而提高番茄果实中番茄红素的含量,改善番茄果实的品质[22]。在本研究中,利用VIGS技术诱导紫色番茄果实中LYC-B基因沉默后,LYC-B基因的表达量降低了75%(图3),且LYC-B基因沉默后,果实中番茄红素的含量为12.27 mg/100 g,显著高于阴性对照组和空白对照组(图4),这与前人研究结果类似[8,23]。另外,沉默LYC-B基因后β-胡萝卜素的含量也高于两个对照组(图4),这与前人[24]采用农杆菌进行遗传转化LYC-B反义基因使得转基因番茄果实中β-胡萝卜素含量降低的结果有所不同。推测可能是由于本实验是在果实绿熟期沉默LYC-B基因,果实早期发育过程中形成的残余的LYC-B基因可以低水平的编码番茄红素β-环化酶,使部分番茄红素转化为β-胡萝卜素,可能导致β-胡萝卜素的含量高于对照。阴性对照组与空白对照组相比番茄红素含量增加,β-胡萝卜素含量差异不显著。这可能是因为类胡萝卜素代谢是植物体内一个主要而复杂的次生代谢系统,不仅仅与植物的营养和风味相关,可能与植物的应激反应也有关系,病毒载体的进入以及注射很可能启动了应激反应系统,引起色素含量发生变化,具体原因还需要进一步研究。
在番茄中,已经鉴定出超过400 种挥发性物质,其中仅有16 种挥发性物质对番茄风味具有重要意义[2]。本研究在紫色番茄中检测出了顺-3-己烯醇、6-甲基-5-庚烯-2-酮、水杨酸甲酯、1-戊烯-3-酮、反-2-庚烯醛、反-2-己烯醛、己醛、2-苯基乙醇、苯乙醛、2-异丁基噻唑等10 种物质,其他如顺-3-己烯醛、β-紫罗兰酮、β-大马酮等物质没有检测到(表1),常培培等[10]在紫色品种中检测到的香叶基丙酮、棕榈酸乙酯在本研究中也没有检测出,这可能是由于栽培条件以及离体研究对挥发性物质的种类产生了影响[25-26]。
番茄果实挥发性物质中包含一些特殊气味物质,能增加果实的清香气味,提高果实的风味品质[27],如本研究中检测到LYC-B基因沉默后含量增加的物质:6-甲基-5-庚烯-2-酮、反-2-辛烯醛、己醇、反-2-庚烯醛、水杨酸甲酯、顺-3-己烯醇等,LYC-B基因沉默后出现的特有物质6-甲基-5-庚烯-2-醇和反-柠檬醛。6-甲基-5-庚烯-2-酮和6-甲基-5-庚烯-2-醇的前体物质是番茄红素,当番茄红素的含量升高时,二者的释放量也相应的增加[5];反-柠檬醛是类胡萝卜素裂解的产物,是合成紫罗兰酮及大马酮等物质的原料,呈柠檬香味[28];反-2-辛烯醛能提高果实的青叶香气,己醇能释放果香味。关于番茄果实挥发性物质与品质性状相关性分析的研究表明[29],反-2-辛烯醛与番茄红素的相关性达到了0.931,反-2-辛烯醛与己醇成显著正相关;反-2-庚烯醛与番茄甜味物质有关,呈现脂肪气味、青草香;水杨酸甲酯,又称冬青油,有独特的气味;顺-3-己烯醇具有类似异戊醇的香味,呈现新鲜的青叶香气[30]。本研究利用VIGS技术将LYC-B基因沉默后,番茄果实中的这些特殊气味物质的含量普遍增加(表1,图6),影响了番茄果实的风味品质。
4 结 论
利用病毒诱导的基因沉默技术成功抑制了LYC-B基因的表达量,LYC-B基因沉默一方面可通过抑制番茄红素的环化来增加紫色番茄果实中番茄红素的含量,提高番茄果实的营养品质;另一方面可通过增加以类胡萝卜素为前体的挥发性物质6-甲基-5-庚烯-2-酮、6-甲基-5-庚烯-2-醇和反-柠檬醛的释放为番茄果实提供独特的香气成分,通过增加反-2-辛烯醛、己醇等番茄红素相关物质以及反-2-庚烯醛、水杨酸甲酯、顺-3-己烯醇等番茄主要挥发性物质的含量影响番茄果实的风味品质。但LYC-B基因如何对这些挥发性物质进行调控,以及这些挥发性物质如何相互作用于风味品质目前尚不得而知,需要进一步地探讨和验证。总之,LYC-B基因既影响番茄果实的类胡萝卜素代谢,又影响挥发性物质的形成,类胡萝卜素代谢与挥发性物质形成密切相关。
[1]摆玉芬, 刘玉梅. 番茄红素分析方法的研究进展[J]. 食品科学, 2014,35(3): 264-268. doi:10.7506/spkx1002-6630-201403052.
[2]刘明池, 郝静, 唐晓伟. 番茄果实芳香物质的研究进展[J]. 中国农业科学, 2008, 41(5): 1444-1451.
[3]LEWINSOHN E, SITRIT Y, BAR E, et al. Not just colors-carotenoid degradation as a link between pigmentation and aroma in tomato and watermelon fruit[J]. Trends in Food Science & Technology, 2005,16(9): 407-415.
[4]VOGEL J T, TIEMAN D M, SIMS C A, et al. Carotenoid content impacts flavor acceptability in tomato (Solanum lycopersicum)[J]. Journal of the Science of Food and Agriculture, 2010, 90(13): 2233-2240.
[5]SOCACI S A, SOCACIU C, MURESAN C, et al. Chemometric discrimination of different tomato cultivars based on their volatile fingerprint in relation to lycopene and total phenolics content[J]. Phytochemical Analysis, 2014, 25(2): 161-169.
[6]FAY L B, NEWTON A, SIMIAN H, et al. Potential of gas chromatography-orthogonal acceleration time-of-flight mass spectrometry (GC-oaTOFMS) in flavor research[J]. Journal of Agricultural and Food Chemistry, 2003, 51(9): 2708-2713.
[7]朱长甫, 陈星, 王英典. 植物类胡萝卜素生物合成及其相关基因在基因工程中的应用[J]. 植物生理与分子生物学学报, 2005, 30(6):609-618.
[8]徐加新, 梁燕. LYC-B基因反义表达对番茄果实番茄红素含量的影响[J]. 西北农林科技大学学报: 自然科学版, 2009, 37(12): 127-132.
[9]阮美颖, 叶青静, 周国治, 等. 不同颜色樱桃番茄果实成熟过程中色素的变化[J]. 浙江农业科学, 2013(5): 526-528.
[10] 常培培, 张静, 杨建华, 等. 紫色番茄果实挥发性风味物质分析[J].食品科学, 2014, 35(14): 165-169. doi:10.7506/spkx1002-6630-201414032.
[11] QUADRANA L, RODRIGUEZ M C, L.PEZ M, et al. Coupling virus-induced gene silencing to exogenous green fluorescence protein expression provides a highly efficient system for functional genomics in Arabidopsis and across all stages of tomato fruit development[J]. Plant Physiology, 2011, 156(3): 1278-1291.
[12] ROMERO I, TIKUNOV Y, BOVY A. Virus-induced gene silencing in detached tomatoes and biochemical effects of phytoene desaturase gene silencing[J]. Journal of Plant Physiology, 2011, 168(10): 1129-1135.
[13] 李锡香, 杜永臣. 番茄种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2006: 9.
[14] 李翠. 番茄SpMPKs基因响应非生物胁迫的功能分析[D]. 杨凌: 西北农林科技大学, 2014.
[15] ORZAEZ D, MIRABEL S, WIELAND W H, et al. Agroinjection of tomato fruits. A tool for rapid functional analysis of transgenes directly in fruit[J]. Plant Physiology, 2006, 140(1): 3-11.
[16] 刘沐霖, 惠伯棣, 庞克诺. 番茄及其制品中番茄红素含量的C18-HPLC-PDA定量分析[J]. 食品科学, 2007, 2 8(7): 453-456.
[17] VILJANEN K, LILLE M, HEINI R L, et al. Effect of high-pressure processing on volatile composition an d odour of cherry tomato puree[J]. Food Chemistry, 2011, 129(4): 1759-1765.
[18] 唐晓伟, 刘明池, 何洪巨,等. 新鲜番茄风味组分研究[J]. 食品科学,2007, 28(2): 28-30.
[19] 甘霖, 申琳, 生吉萍. 秸秆源品质改良因子采前处理对番茄果实品质的影响[J]. 食品科学, 2013, 34(4): 221-225.
[20] 侯雷平, 吴俊华, 李梅兰, 等. 供锌水平对番茄果实抗氧化性及风味品质的影响[J]. 植物营养与肥料学报, 2010, 16(3): 763-767.
[21] 陈书霞, 林海军.番茄果实不同发育阶段香气成分组成及变化[J].西北植物学报, 2010, 30(11): 2258-2264.
[22] 万群, 张兴国, 宋明. 果实特异性RNAi介导的Lcy基因沉默来增加番茄中番茄红素的含量[J]. 生物工程学报, 2007, 23(3): 429-433.
[23] 吴江敏, 孙亚东, 梁燕, 等. 番茄红素-β-环化酶(LYCb)反义基因在番茄转基因后代的表达及其遗传稳定 性[J]. 农业生物技术学报, 2011,19(5): 801-807.
[24] ROSATI C, AQUILANI R, DHARMAPURI S, et al. Metabolic engineering of beta-carotene and lycopene content in tomato fruit[J]. The Plant Journal, 2000, 24(3): 413-420.
[25] CARLI P, BARONE A, FOGLIANOV, et al. Dissection of genetic and environmental factors involved in tomato organoleptic quality[J]. BMC Plant Biology, 2011, 11(1): 58-68.
[26] TIEMAN D, BLISS P, MCINTYRE L M, et al. The chemical interactions underlying tomato fla vor preferences[J]. Current Biology,2012, 22(11): 1035-1039.
[27] 郝静. 番茄果实风味物质及其农艺影响因子的研究[D]. 北京: 中国农业大学, 2007.
[28] SIMKIN A J, S CHWARTZ S H, AULDRIDGE M, et al. The tomato carotenoid cleavage dioxygenase 1 genes contribute to the formation of the flavorvolatiles β-ionone, pseudoionone, and geranylacetone[J]. The Plant Journal, 2004, 40(6): 882-892.
[29] 常培培,梁燕, 张静, 等. 5 种不同果色樱桃番茄品种果实挥发性物质及品质特性分析[J]. 食品科学, 2014, 35(22): 215-221. doi:10.7506/spkx1002-6630-201422042.
[30] 密森. 番茄果实主要芳香物质含量和营养品质组成 变化的研究[D].扬州: 扬州大学, 2012.
Effect of LYC-B Gene Silencing on Major Pigments and Volatile Compounds in Purple Tomato Fruits
L. Jie, LIANG Yan*, ZHAO Jingjing, ZHANG Yan, CHANG Peipei, QIN Lei, LI Yunzhou
(College of Horticulture, Northwest A&F University, Yangling712100, China)
In the current study, the expression of lycopene beta-cyclase (LYC-B) gene was inhibited during ripening through virus-induced silencing (VIGS) technique and the corresponding impact on major pigments and volatile compounds in purple tomato fruits was analyzed. The results showed that LYC-B gene silencing could increase the content of lycopene in purple tomato fruits. Tomato volatile compounds detected by headspace solid phase micro-extraction (HS-SPME) coupled with gas chromatography-mass spectrometry (GC-MS) revealed that LYC-B gene silencing could increase the content of 6-methyl-5-hepten-2-one, which is derived from carotenoids. The release of (E)-2-octenal, hexanol, (E)-2-heptenal, methyl salicylate and (Z)-3-hexen-1-ol was also increased. In addition, LYC-B gene silencing could increase the contents of lycopene and some major volatile compounds, thus improving the nutritional and flavor quality of tomato fruits.
tomato; LYC-B gene silencing; lycopene; volatile compounds
S641.2
A
1002-6630(2015)23-0221-07
10.7506/spkx1002-6630-201523041
2015-01-31
陕西省科技统筹创新计划项目(2011KTCL02-03);西北农林科技大学农业科技推广基金项目(TGZX2012-02)
吕洁(1990—),女,硕士研究生,研究方向为番茄果实品质性状。E-mail:lvjielj@163.com
梁燕(1963—),女,教授,博士,研究方向为番茄遗传育种与蔬菜种质资源。E-mail:liangyan@nwsuaf.edu.cn
