精河沙区土壤酶分布特征及其对土壤理化性状的响应
2015-10-13姚健等
姚健等

摘要:分析了新疆精河沙区土壤过氧化氢酶、蔗糖酶、脲酶的活性及土壤有机质等理化性状。结果表明,蔗糖酶活性变化范围在25~535 mL/μg,脲酶活性在0~150.29 mg/100 g,过氧化氢酶活性在18~20 mL/g。酶活性随时空动态变化的原因是人为活动、土壤环境、水文条件等因素综合作用的结果。酶活性的垂直分布受土壤通透性、酶属性等因素的影响,导致出现酶活性随土层深度增加而减弱的趋势,少部分样地在人为活动影响下则出现随土层深度增加而增强的趋势。依据通径分析原理,与过氧化氢酶活性相关性由大到小的顺序为pH、容重、温度、有机质;与蔗糖酶活性相关性由大到小的顺序为温度、容重、pH、有机质;与脲酶活性相关性由大到小的顺序为pH、有机质、容重、温度。有机质、温度及容重主要通过pH间接对过氧化氢酶及脲酶活性产生轻微影响,有机质、温度、pH则通过容重来间接影响蔗糖酶活性。
关键词:土壤酶活性;土壤有机质;理化性状;分布特征;通径分析
中图分类号:S154.1 文献标识码:A 文章编号:0439-8114(2015)18-4435-06
DOI:10.14088/j.cnki.issn0439-8114.2015.18.013
土壤酶是土壤组分中最活跃的有机成分之一,是土壤生物过程的主要调节者[1],其参与了土壤环境中的一切生物化学过程,与有机物质分解、营养物质循环、能量转移、环境质量等密切相关[2,3]。土壤生态系统中生物化学过程和物质循环均是在土壤酶的催化作用下进行的[4]。土壤酶的分解作用参与并控制着土壤中的生物化学过程在内的自然界物质循环过程,酶活性的高低直接影响物质转化循环的速率,因而土壤酶活性对生态系统功能有很大的影响。在几乎所有生态系统的监测和研究中,土壤酶活性的检测似乎成了必不可少的测定指标[5]。土壤酶活性的强弱也是土壤肥力的重要标志,故针对土壤酶活性的研究对于改善区域土壤肥力及土壤改良意义重大。土壤有机质是指土壤中含碳的有机化合物,是供给作物生长的营养元素,可直接为作物提供生长所需养分,也可以改善土壤的物理性状,是评价土壤自然肥力的一个重要因素,也是研究土壤可持续利用中一个重要指标[6]。
土壤有机质是土壤酶的直接载体,两者之间存在非常密切的联系,许多学者通过测量土壤酶活性强度来判断土壤肥力的高低[7],但两者之间的相关性则取决于土壤酶的种类与属性。刘建新[8]研究表明,土壤脲酶与土壤养分之间的相关性最好,它与土壤有机质、全氮、全磷、全钾及速效氮、磷之间都存在显著相关,磷酸酶和过氧化氢酶与有机质、全氮、全磷、全钾及速效氮都存在着显著相关,但转化酶和纤维素酶活性与土壤各养分之间没有良好的相关性。土壤酶活性的高低,与土壤自身生态理化性状之间(土壤温度、水分、pH等)均有着密切联系,故针对土壤酶活性的时空分布及其与理化性状之间的关系研究成为许多学者研究的热点。Luo等[9]研究表明随着土壤深度的改变,土壤酶活性差异较大。杨万勤等[10]对北碚缙云山森林生态系统中4种群落的过氧化氢酶、转化酶、蛋白酶和酸性磷酸酶活性的分布特征和季节动态进行了研究,结果表明土壤酶活性随土层加深酶活性降低;土壤酶活性的季节变化规律很明显,但不同的酶类,其变化规律不同,但均是冬季酶活性最低,并且,土壤酶活性的季节动态还与土温的季节变化及酶本身的性质有关。Hakulinen等[11]对位于芬兰北部郊区的3个湖泊水系沉积物的水解酶活性垂直分布情况进行了研究。结果表明,磷酸酶等10种水解酶活性随湖泊沉积物深度增加而降低,并与沉积物表层相比,位于深层的水解酶也具有较高活性,说明在缺氧条件下,水解酶依然具有活性。Tiwari等[12]研究表明脲酶和脱氢酶与土壤水分呈正相关关系。有研究[13,14]表明,土壤温度对土壤酶活性有较强的影响:当温度由10 ℃上升到60 ℃时,土壤酶活性显著增加;但当温度进一步升高,脲酶迅速钝化;在150 ℃下加热24 h,土壤酶会完全失活。在不同温度培养条件下,有机质含量最低的土壤,其酶活性也最低,温度对氧化还原酶活性有显著影响[15]。土壤pH主要通过影响土壤微生物种类而影响微生物释放酶的种类及速度,并能够控制酶促反应的速率。有关研究显示,H+可以改变酶促反应基点和酶的稳定性[16],有些酶促反应甚至只能在很小范围的pH内进行,并且由不同微生物分泌出的催化同一反应的酶的最适pH不同[17]。一般情况下,土壤湿度较大时,酶活性较高,但土壤过湿时,酶活性减弱[18]。
土地利用造成土地覆盖变化,而土地覆盖变化对陆地生态系统以及生物地球化学循环产生直接影响,并通过改变土壤特性,导致土壤退化,对土壤养分迁移变化以及水文循环有着十分显著的影响[19]。因此,研究干旱区土壤酶与土壤有机质对区域自然理化性状的响应和加深土壤有机质、酶活性等在生态系统物质循环过程中的作用等,有着十分重要的意义。在利用线性回归分析土壤酶活性与有机质及土壤温度、水分、pH的相关性的基础上,利用通径分析算法来分析土壤酶活性与土壤有机质及理化性状之间的通径系数等,旨在对沙区土地可持续利用,减缓区域土壤生态系统退化以及区域生态环境维护提供科学依据。
1 材料与方法
1.1 研究区概况
精河县隶属新疆维吾尔自治区博尔塔拉蒙古自治州,位于新疆维吾尔自治区西北部,天山支脉婆罗科努山北麓,准噶尔盆地西南边缘。南部为天山山区,面积4 642 km2,为总面积的41%。中部为博尔塔拉河冲积平原,面积约为6 111 km2,占总面积的54%,地势由南向北倾斜,地形平坦开阔,多为戈壁荒漠,其次是耕地,平均海拔230~440 m;北部为冲积-洪积平原,大部分地下水位偏高,多为盐碱沼泽地带,艾比湖在县区最北[20]。全县国土面积11 289 km2,耕地面积412.6 km2,境内自然荒漠以及海拔800 m以上的山地占全县66%以上的面积。精河县气候属典型的北温带干旱荒漠型大陆性气候。主要特点是光照充足,干燥少雨,蒸发量大。1月份平均气温-15 ℃,7月份平均气温26 ℃,极端最低气温为-34 ℃,极端最高气温为42 ℃;年平均降水量102 mm。
1.2 样品采集
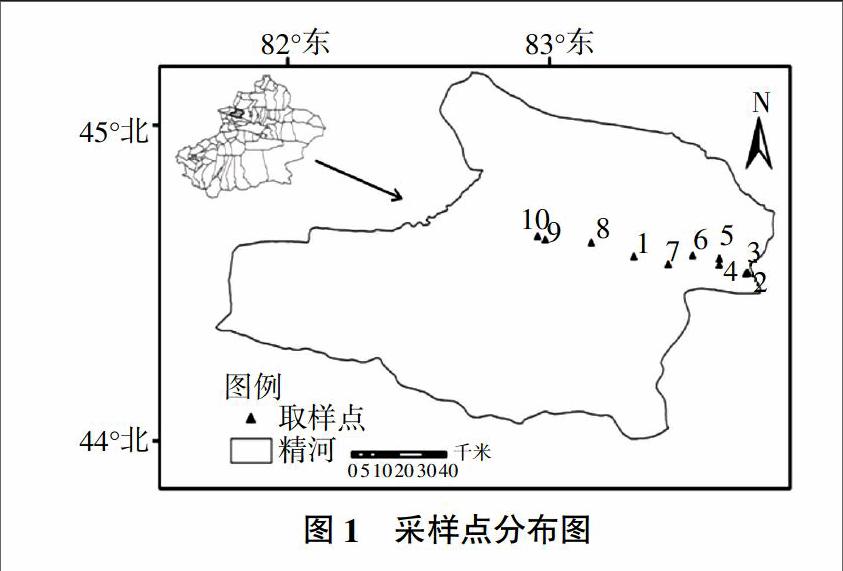
土样采自新疆博尔塔拉自治州精河县,依据宏观自然地理特征及区域景观格局现状,利用便携式GPS进行定位,合理选取具有代表性土壤样地(共10个土壤剖面调查取样,如图1所示)。其中10个样地分别代表不同土地覆被及不同土地利用类型:1号样地为荒漠沙土地;2号样地为农田荒漠交错带;3号样地为人工小型农田防护林;4号样地为农田枸杞样地;5号样地为农田棉花地;6号样地为绿洲自然灌丛;7号样地为高速公路边绿洲内部低盖度土壤样地;8号样地为沙区北部盐碱草地;9号样地为高速公路旁人工防护林;10号样地为人工片林。在采集土样的过程中,分别按照0~20、20~40、40~60、60~80、80~100 cm不同剖面深度利用环刀采集土壤样本,去除植物根系、碎屑等,样品装入塑料密封袋,带回室内5 ℃保存,以便进行各项测试[21]。取样过程中利用便携式Hydra土壤水分/温度/电导率测量仪测量不同深度剖面各项理化数据。
1.3 试验方法
将采集的新鲜土壤样品磨碎,自然风干并装袋。做3次平行测定,具体测定方法如下。土壤容重采用环刀法;土壤蔗糖酶活性采用硫代硫酸钠滴定法测定;过氧化氢酶活性采用高锰酸钾滴定法测定;土壤脲酶活性采用可见光分光光度计法测定;土壤有机质采用重铬酸钾氧化法[22]测定。利用EXCEL 2007对试验数据进行处理。
2 结果与分析
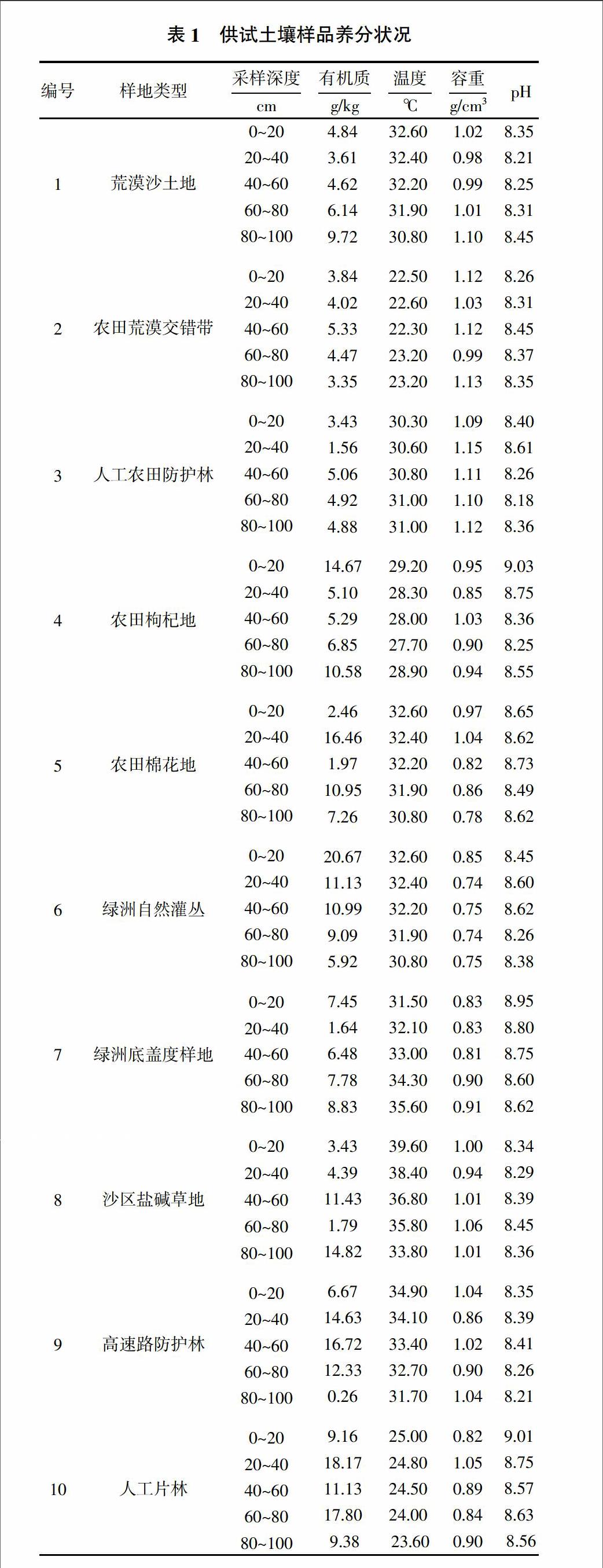
利用上述试验方法,对10个土壤样地,50个土壤样本进行试验,供试土壤养分状况如表1所示。
2.1 土壤酶活性的时空动态变化分析
土壤酶活性对于土壤中有机物质转化、释放植物能利用的碳、氮、磷等养分有直接的作用[23]。其中,过氧化氢酶能够酶促土壤中对植物或生物有负效应的过氧化氢物质的分解,脲酶能够将尿素水解成胺,可供植被吸收利用,蔗糖酶参与的反应是土壤碳循环的主要环节,其主要作用是催化蔗糖水解成葡萄糖和果糖,为植物生长提供所需能量。这3种酶在土壤碳、氮元素循环代谢过程中有重要的作用,部分学者认为土壤酶活性是土壤中生物化学过程的总体现,可以综合反映出土壤性质和肥力水平[21]。
由表2可以得出,蔗糖酶的变化范围为25~535 mL/μg NaS2O4,其活性强弱随时空动态变化具有很强的差异性,其最高值出现在4号样地,其次是5、3、6、7、8、9、10、2、1号样地。过氧化氢酶活性变化范围不大,大多数处于18~20 mL/g KMnO4,其中最高值出现在10号样地,最低值出现在2号样地,其余各样地过氧化氢酶活性变化不是十分明显。脲酶活性由表2可以看出,变化范围主要集中在0~150.29 NH3-N(mg/100 g),最高值出现在5号样地,最低值在3号样地,脲酶活性几乎趋近于0,其次2号样地与1号样地脲酶活性较低。相对而言,4号、5号等样地土壤酶活性较强,而1号,2号等样地土壤酶活性较弱。由于每个采样点具有不同的土壤类型与环境,包括气候及水文条件等,是导致酶活性具有较强时空动态变化的原因。从表2中可以看出,酶活性强度较高的样地,主要是农田、人工林地等被利用地,而酶活性强度较低的样地主要集中在荒漠、盐碱地等未被利用地。土壤酶活性的时空动态变化是土壤环境、气候、水分、人为活动等因素的综合作用的结果。
2.2 土壤酶活性的垂直分布特性分析
从表2中我们可以得出,随着土层深度的增加,蔗糖酶与脲酶活性基本处于逐渐降低的趋势。这是由于随着土层深度的增加,土壤通气性逐渐降低,分泌土壤酶的微生物含量减少,且酶活性分子的存活需要空气,导致酶活性随土层深度增加而减少。只有较少样地出现酶活性随土层深度的增加而增加的现象,这可能与样地本身的土壤利用方式有关,如5号样地的蔗糖酶活性与脲酶活性随土层深度增加而增加,而5号样地是农田棉花地,人为耕作过程增大了土壤的通透性及水热状况,从而对土壤酶活性的垂直分布产生了影响。过氧化氢酶活性随土层深度的增加变化趋势并不太大,这可能与研究区土壤基本状况和过氧化氢酶基本属性相关,各样地土层中过氧化氢含量较少,则消耗掉的酶较少,且过氧化氢酶是一种氧化还原酶,随着土层深度增加,氧化还原反应的难度增大,故过氧化氢酶活性的变化范围不大,它不同于脲酶与蔗糖酶,二者均属于水解类酶,是水解土层中的蔗糖,尿素等物质的,水解反应的要求较低,并不需要很苛刻的反应条件,故蔗糖酶与脲酶变化范围较大,但三者基本上遵循随土层深度增加而减少这一规律。从表2可以得出,大部分酶活性的最大值一般集中在前3个土层中,这是因为这3个土层相对较浅,对水分、阳光等能源的吸收与传递较强等原因所决定的。
2.3 土壤酶活性对土壤理化性状的响应
从土壤发生学角度,土壤酶与土壤有机质等理化性状之间的相关性主要取决于土壤自身类型,沙区典型干旱气候及特殊植被分布是构成沙区土壤类型的重要因素;自然因素及人为因素发展和变化制约着沙区土壤的形成和演化,两种因素也是导致土壤各理化性状间相互依存和作用的原因。土壤酶活性的强弱,是土壤酶对土壤理化性状、土壤类型、植被、气候等环境因素的综合响应[23]。通过对10个样地3种土壤酶活性与土壤有机质、pH、土壤容重及温度的测定,利用通径系数分析计算土壤酶活性对土壤各理化性状的动态响应。将土壤有机质、pH、土壤容重及温度看作原因因素(Xi),土壤酶活性看作结果因素(Y),如表3所示,首先对各原因因素(Xi)进行相关性分析,建立各变量间相关系数矩阵。有机质、温度、容重及pH在对土壤酶活性的变化影响中是相互影响的复杂关系,彼此之间不是独立的,各要素间具有一定的相关性;其次,将土壤过氧化氢酶活性看作为Y1,蔗糖酶活性为Y2,脲酶活性为Y3,利用通径分析原理,计算各原因因素(Xi)对3种酶活性的直接通径系数,如表4所示。然后结合表3数据,计算各原因因素对3种酶活性的间接作用,结果如表5~表7所示。
由表3数据可以得出过氧化氢酶活性与pH的相关性较高,其次是容重和温度,土壤有机质对过氧化氢酶活性的影响相对较低;蔗糖酶活性与温度和容重的相关性较强,其次是pH与有机质;脲酶活性与pH的相关性最高,土壤有机质含量与脲酶活性之间的相关性也较为显著。这也与前人的研究相一致[24-26]。4种土壤理化性状在对3种土壤酶活性的影响中,彼此之间并不孤立,而是存在相互制约与促进的复杂关系。其中pH与容重、容重与有机质具有较为显著的相关性。
根据通径分析原理,pH及温度在对各土层中过氧化氢酶活性的变化直接影响较大,容重和土壤有机质含量的直接通径系数较小;蔗糖酶活性主要依靠土壤容重与温度的直接影响,pH与有机质含量影响较小;针对脲酶活性,pH及有机质含量对其活性的直接影响较大,而温度与容重直接影响较小。由表5-表7的数据可知,土壤有机质、温度及容重主要通过pH间接对过氧化氢酶及脲酶活性产生影响,但作用程度并不是很强烈;而有机质、温度、pH则是通过容重间接影响蔗糖酶活性。
3 结论
1)从总体上看,蔗糖酶与脲酶随时空动态变化的差异性较大,蔗糖酶活性变化范围在25~535 mL/μg NaS2O4;脲酶变化范围在0~150.29 mg/100 g NH3-N。导致酶活性分布差异性较大的原因主要是因为不同采样点具有不同的土壤理化性状、土壤类型、植被覆盖、土壤含水率,盐渍化程度等自然环境因素,也包括人为耕作、利用土地对该区域土壤酶活性产生的影响。利用耕作及种植林地等手段改变干旱区土壤生态环境,在一定程度上对区域土壤酶活性分布起到一定的积极作用。过氧化氢酶随时空动态变化范围不大,大多数均处于18~20 mL/g KMnO4,这可能与酶自身属性和区域土壤中过氧化氢含量较少有关,过氧化氢酶属于氧化还原酶,蔗糖酶与脲酶均为水解酶,其反应条件相对与水解酶而言较为苛刻,故消耗的过氧化氢酶较少,变化范围不大。研究区土壤3种酶自身属性不同,不同酶属性有着不同的反应条件与机理,从而导致3种酶活性变化范围存在差异,但3者基本上均遵循随土层深度增加而减少这一规律,这也与前人的研究结果相一致[7,8-11],且大部分酶活性的最大值一般集中在前3个土层中,这是因为这3个土层相对较浅,对水分、阳光等能源的吸收与传递较强,土壤通透性较强等原因所决定的。
2)在4种土壤理化性状对3种土壤酶活性的影响中,彼此之间并不孤立,其中pH与容重、容重与有机质具有较为显著的相关性。过氧化氢酶活性与pH的相关性较高,其次是容重、温度和土壤有机质;蔗糖酶活性与温度和容重的相关性较强,其次是pH与有机质;脲酶活性与pH的相关性最高,土壤有机质含量与脲酶活性之间的相关性也较为显著。根据通径分析原理可知,土壤有机质、温度及容重主要通过pH间接对过氧化氢酶及脲酶活性产生影响,但作用程度并不是很强烈;而有机质、温度、pH则通过容重间接影响蔗糖酶的活性。
3)通过研究精河沙区土壤酶活性时空动态以及垂直分布特征及其对各理化性状包括土壤有机质的响应,旨在对沙区土地资源合理与可持续利用,维护区域生态环境现状,水土资源高度耦合模式提供依据。结果表明,精河沙区土壤酶活性与土壤有机质含量总体偏低,作者通过实地调查取证发现沙区土壤利用程度相对较低,一般都选择生态环境较好的区域进行耕作、种植等,针对较为干燥和盐渍化程度较深,水土资源较为匮乏的土地,开发和利用程度较低,长久如此则可能导致精河沙区土壤生态格局出现不平衡发展趋势。建议在土壤环境较弱,资源相对缺失的地区推广种植耐旱耐盐碱的作物,如罗布麻等作物,不仅能为沙区带来经济收益,还能改善区域生态环境,提高土壤理化性状水平,实现沙区水土资源高度耦合的发展模式。
参考文献:
[1] MARX M C, WOOD M, JARVIS S C. A microplate fluorimetric assay for the study of enzyme diversity in soils [J]. Soil Biology and Biochemistry,2001,33(12):1633-1640.
[2] YAO XH,MIN H,L?譈 ZH, et al. Influence of acetamiprid on soil enzymatic activities and respiration[J]. European Journal of Soil Biology,2006,42(2):120-126.
[3] 杨万勤,王开运.土壤酶研究动态与展望[J].应用与环境生物学报,2002,8(5):564-570.
[4] 周智彬,徐新文.塔里木沙漠公路防护林土壤酶分布特征及其与有机质的关系[J].水土保持学报,2005,18(5):10-14.
[5] 邱莉萍,刘 军,王益权,等.土壤酶活性与土壤肥力的关系研究[J].植物营养与肥料学报,2004,10(3):277-280.
[6] WIESMEIER M,PRIETZEL J,BARTHOLD F,et al. Storage and drivers of organic carbon in forest soils of southeast Germany(Bavaria)–Implications for carbon sequestration[J]. Forest Ecology and Management,2013,295:162-172.
[7] 王 娟,谷雪景,赵 吉.羊草草原土壤酶活性对土壤肥力的指示作用[J].农业环境科学学报,2006,25(4):934-938.
[8] 刘建新.不同农田土壤酶活性与土壤养分相关关系研究[J].土壤通报,2004,35(4):523-525.
[9] LUO J,TILLMAN R W, WHITE R E, et al. Variation in denitrification activity with soil depth under pasture[J]. Soil Biology and Biochemistry,1998,30(7):897-903.
[10] 杨万勤,钟章成.缙云山森林土壤酶活性的分布特征,季节动态及其与四川大头茶的关系研究[J].西南师范大学学报:自然科学版,1999,24(3):318-324.
[11] HAKULINEN R,K?魧HK?魻NEN M A,SALKINOJA-SALONEN M. Vertical distribution of sediment enzyme activities involved in the cycling of carbon, nitrogen,phosphorus and sulphur in three boreal rural lakes[J]. Water Research,2005,39(11):2319-2326.
[12] TIWARI M B,TIWARI B K, MISHRA R R. Enzyme activity and carbon dioxide evolution from upland and wetland rice soils under three agricultural practices in hilly regions[J]. Biology and Fertility of Soils,1989,7(4):359-364.
[13] 林玉锁.土壤对重金属缓冲性能的研究[J].环境科学学报,1995,15(3):289-293.
[14] 和文祥,朱铭莪, 张一平.土壤酶与重金属关系的研究现状[J].土壤与环境,2000,9(2):139-142.
[15] TRASAR C C, GIL S F, LEIROS M C. Thermodynamic parameters of enzymes in grassland soils from Galicia, NW Spain[J]. Soil Biology and Biochemistry,2007,39(1):311-319.
[16] KANG H, FREEMAN C. Phosphatase and arylsulphatase activities in wetland soils: annual variation and controlling factors[J]. Soil Biology and Biochemistry,1999,31(3):449-454.
[17] NIEMI R M, VEPS?魧L?魧INEN M. Stability of the fluorogenic enzyme substrates and pH optima of enzyme activities in different Finnish soils[J]. Journal of Microbiological Methods, 2005,60(2):195-205.
[18] 关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.
[19] 王让会,宁虎森,赵振勇,等.绿洲土壤水盐耦合关系及景观地球化学特征[J].南京信息工程大学学报:自然科学版,2009, 1(2):97-101.
[20] 李秀花,郭 凯.新疆精河县近20年土地利用/覆被变化动态研究[J].干旱区资源与环境,2011,25(6):88-91.
[21] 陈祝春.科尔沁沙地沙丘土壤生物学活性[J].中国沙漠,1995,15(3):283-287.
[22] 王让会,樊自立,马英杰.干旱区水域生态系统的水盐耦合关系——以新疆塔里木河流域为例[J].应用生态学报.2002, 13(2):204-208.
[23] DOELMAN P,HAANSTRA L. Short-term and long-term effects of cadmium,chromium,copper,nickel,lead and zinc on soil microbial respiration in relation to abiotic soil factors[J]. Plant and Soil,1984,79(3):317-327.
[24] WALLENIUS K, RITA H, SIMPANEN S, et al. Sample storage for soil enzyme activity and bacterial community profiles[J]. Journal of Microbiological Methods,2010,81(1):48-55.
[25] 邱现奎,董元杰,万勇善,等.不同施肥处理对土壤养分含量及土壤酶活性的影响[J].土壤,2010,42(2):249-255.
[26] 陆 梅,田 昆,陈玉惠,等.高原湿地纳帕海退化土壤养分与酶活性研究[J].西南林学院学报,2004,24(1):34-37.
