竹柳无性系光合特性的研究
2015-09-17任宣百陆志民
任宣百,陆志民
(吉林省林业科学研究院,吉林 长春 130033)
竹柳(Salix‘Zhuliu’)为杨柳科(Salicaceae)柳属(Salix)植物,是经过选育及驯化而培育出的优良树种,具有树形美观、生长速度快、抗性强等优点。竹柳作为工业用材料和速生丰产林已在北京、河北、山东、安徽等地大面积种植,并取得良好经济和生态效益。目前在东北吉林地区种植竹柳虽尚处试验阶段,但竹柳引种到该地区就表现出速生优势,当年生长量巨大。作者对2 a 生竹柳3 个无性系进行光合生理特性研究,以确定其光合适应性差异以及环境因子对物质积累的影响效应。
1 试验地概况
试验地位于松原市前郭县蒙古屯乡,地理位置:45°15'~45°17'N,124°38'~124°40'E。该地块处于松嫩平原地区,属于中温带大陆性季风气候区,年平均气温4.5℃,≥10℃有效积温3 160℃,年平均日照时数2 897 h,无霜期142 d 左右。年平均降水量400 ~500 mm。土壤类型为薄层草甸黑钙土。
2 试验材料
从美国引进实生苗,分别通过组织培养技术培育出组培苗,组培苗移栽大田并生长1 a后截取插穗,再利用扦插技术培育出3 个无性系苗木,在试验地生长2 a。以2 a 生‘垂暴109’柳为对照。
3 试验方法
2013 年7 月下旬选择晴朗天气条件,采用手持CI-340 型光合作用测量系统,从5:00 开始至19:00 结束,每隔30 min 测量1 次,测量指标为叶片瞬时净光合速率(Pn)、光照强度(PAR)、环境CO2浓度(Ci)等,重复测量3 次并取平均值。所有数据均采用Microsoft Excel 2003 进行汇总整理、SPSS 19.0 进行数据分析。
3.1 光合作用光响应过程
设置的光合有效辐射强度梯度为:1 600、1 400、1 200、1 000、800、600、400、200、100、50、25 μmol·m-2·s-1,利用非直角双曲线模型对竹柳3 个无性系和对照‘109’柳实测光照强度(PAR)光响应曲线数据进行拟合,计算出迭代参数值并绘制出光合作用光响应曲线。当光照强度小于200 μmol·m-2·s-1时,对该段曲线进行线性回归拟合,利用公式Pn=αPAR+ Rd求得光饱和点(LSP)、光补偿点(LCP)和表观光合量子效率(AQY)的数值。
3.2 光合作用CO2 响应过程
在光照强度(PAR)接近光饱和点时,设置CO2浓度梯度为:1 200、1 000、800、600、500、400、300、200、150、100、50 μmol·mol-1,利用直角双曲线修正模型对竹柳3 个无性系和对照‘109’柳的实测CO2浓度(Ci)响应曲线数据进行拟合,做出CO2响应曲线。当CO2浓度小于200 μmol·mol-1时,对该段曲线进行线性回归拟合,利用公式Pn=αCi+Rp求得CO2饱和点(CSP)、CO2补偿点(CCP)和羧化效率(CE)的数值。
3.3 净光合速率(Pn)日变化
根据不同时间点的实测数值,利用2 次多项式线性回归做出趋势线,模型表达式为:

式中:y——净光合速率(Pn);
x——实测光照强度(PAR)。
3.4 非直角双曲线模型
非直角双曲线模型具有参数丰富、模拟效果好等特点,其模型表达式为:

式中:Pn——净光合速率;
α——表观量子效率;
Pmax——最大净光合速率;
PAR——光合有效辐射;
k——光响应曲线曲角;
Rd——暗呼吸速率。
3.5 直角双曲线修正模型
直角双曲线修正模型适用范围广,拟合结果更接近实测值,其模型表达式为:

式中:Pn——净光合速率;
α——CO2响应曲线初始斜率;
β、γ——修正系数;
Rp——暗呼吸速率。
4 结果与分析
4.1 光响应参数
利用非直角双曲线模型对竹柳3 个无性系和对照‘109’柳实测光照强度(PAR)数据进行拟合,结果见表1。

表1 非直角双曲线模型参数SPSS 迭代值Tab.1 Model parameter SPSS substituting value of non-rectangular hyperbola
从表1 中可以看出,所有R2数值在0.999及以上,说明拟合程度非常好,可以真实反映出光照强度(PAR)与叶片瞬时净光合速率(Pn)的关系。暗呼吸速率(Rd)在 0.737 ~1.750 μmol·m-2·s-1之间,对照‘109’柳暗呼吸速率为1.750 μmol·m-2·s-1,为最大值;竹柳无性系 1 暗呼吸速率为0.737 μmol·m-2·s-1,为最小值。最大光合速率(Pmax)在21.6 ~29.5 μmol·m-2·s-1之间,竹柳无性系 3 最大光合速率为29.5 μmol·m-2·s-1,为最大值;对照‘109’柳最大光合速率为21.6 μmol·m-2·s-1,为最小值。竹柳3 个无性系最大光合速率都明显大于对照‘109’柳最大光合速率,而对照‘109’柳暗呼吸速率则明显大于竹柳3 个无性系。
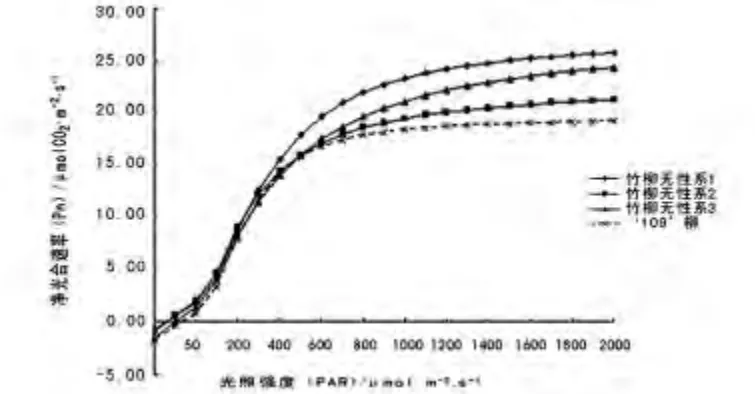
图1 竹柳无性系与‘109’柳的光响应曲线Fig.1 The photoresponse curve of Salix‘Zhuliu’clone and Salix בChuibao109’
4.2 光响应曲线
竹柳3 个无性系与对照‘109’柳树光响应曲线如图1 所示,X 坐标为光照强度(μmol·m-2·s-1),Y 坐标为净光合速率(μmol CO2·m-2·s-1)。
从光响应曲线可以看出,竹柳与对照净光合速率随着光照强度上升而快速达到某个临界值后,其净光合速率随光照强度上升而变为缓慢增加,逐步趋于饱和状态。当光照强度小于25 μmol·m-2·s-1时,净光合速率大小顺序为:竹 柳 无 性 系 1 > 竹 柳 无 性 系 2 >竹柳无性系3 >对照;当光照强度在25 ~200 μmol·m-2·s-1时,净光合速率大小顺序为:竹柳无性系2 >竹柳无性系1 >竹柳无性系3 >对照;当光照强度在300 ~400 μmol·m-2·s-1时,净光合速率大小顺序为:竹柳无性系1 >竹柳无性系2 >对照>竹柳无性系3;当光照强度大于500 μmol·m-2·s-1时,净光合速率大小顺序为:竹柳无性系1 >竹柳无性系3>竹柳无性系2 >对照,竹柳光合作用效率比‘109’柳高很多。
4.3 光响应线性回归拟合
在光照强度PAR <200 μmol·m-2·s-1时,对该段曲线进行线性回归拟合,并作出回归方程Pn=αPAR + Rd。当回归方程Pn=Pmax时,可以计算出光饱和点(LSP)数值;当回归方程Pn=0 时,可以计算出光补偿点(LCP)数值;回归方程斜率即为表观光合量子效率(AQY),结果见表2。

表2 光照强度线性回归拟合方程、LSP、LCP 和AQY 值Tab.2 Illumination intensity linear regression fitted equation,LSP、LCP and AQY value

?
从表2 中可以看出,模型所有系数Sig <0.01,说明方程通过F 检验,线性回归分析有效;R2数值均在0.99 以上,说明高度拟合,可以真实反映光响应曲线走势。光饱和点在482.02 ~648.55 μmol·m-2·s-1之间,竹柳无性系3 光饱和点为648.55 μmol·m-2·s-1,是模型中最大值;对照‘109’柳光饱和点为482.02 μmol·m-2·s-1,是模型中最小值。光补偿点在7.81 ~32.02 μmol·m-2·s-1之间,对照‘109’柳光补偿点为32.02 μmol·m-2·s-1,是模型中最大值;竹柳无性系2 光补偿点最小为7.81 μmol·m-2·s-1,是模型中最小值。表观光合量子效率试验材料与对照基本一致。
4.4 CO2 响应参数
利用直角双曲线修正模型对竹柳3 个无性系和对照‘109’柳实测环境CO2(Ci)数据进行拟合,经过迭代计算之后,各参数值见表3。
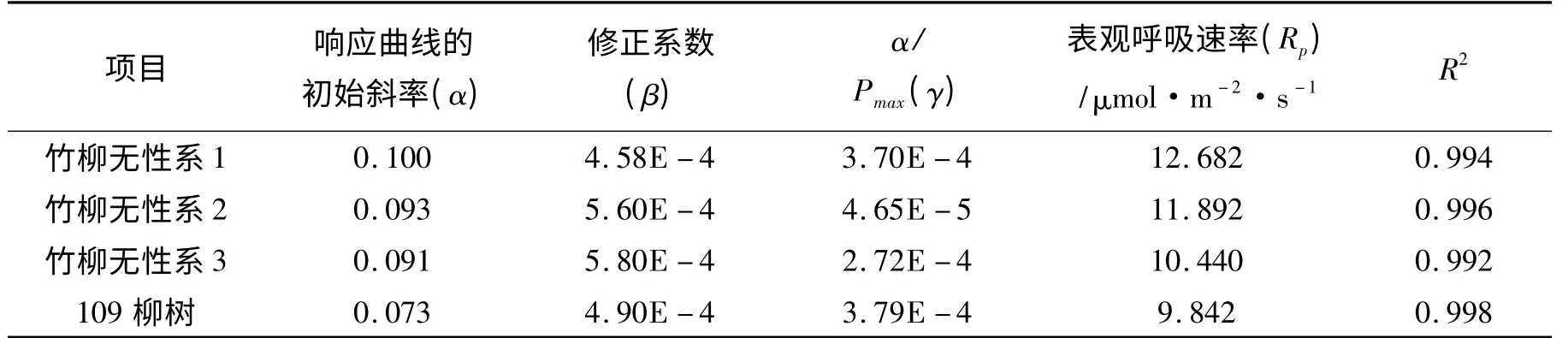
表3 直角双曲线修正模型参数SPSS 迭代值Tab.3 Rectangular hyperbola modified model parameters SPSS iteration
从表3 中可以看出,所有R2数值均在0.99 以上,说明拟合程度非常好,可以真实反映出CO2浓度(Ci)与叶片瞬时净光合速率(Pn)关系。表观呼吸速率(Rp)在 9.842 ~12.682 μmol·m-2·s-1之间,竹柳无性系1 表观呼吸速率为12.682 μmolCO2·m-2·s-1,为最大值;对照‘109’柳表观呼吸速率为9.842 μmolCO2·m-2·s-1,为最小值。
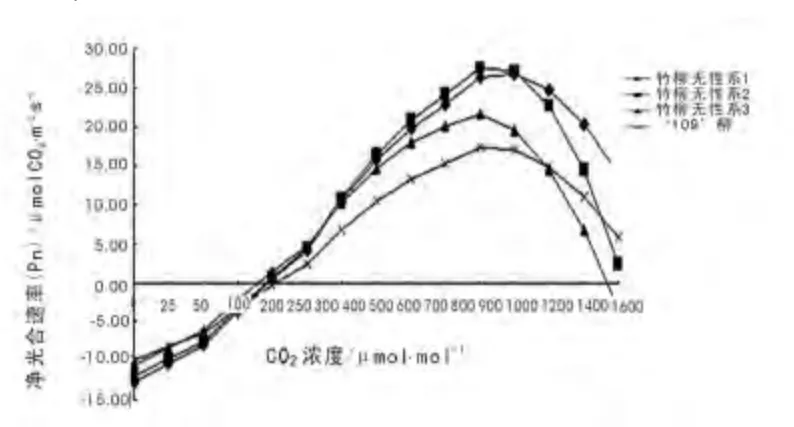
图2 竹柳无性系与‘109’柳的CO2 响应曲线Fig.2 The CO2 response curve of Salix‘Zhuliu’clone and Salix בChuibao109’
4.5 CO2 响应曲线
竹柳3 个无性系与对照‘109’柳的光响应曲线如图1 所示,X 坐标为CO2浓度(μmol·mol-1),Y 坐标为净光合速率(μmolCO2·m-2·s-1)。
从图2 中可以看出,竹柳与‘109’柳净光合速率随着CO2浓度上升而缓慢上升,达到最大值之后净光合速率随着CO2 浓度增加而快速下降。达到最大值时顺序为:竹柳无性系2>竹柳无性系1 >竹柳无性系3 >对照‘109’柳;竹柳无性系2 在CO2浓度为800 μmol·mol-1时最大值为27.70 μmolCO2·m-2·s-1,竹柳无性系1 在CO2浓度为1 000 μmol·mol-1时最大值为26.88 μmolCO2·m-2·s-1,竹柳无性系3 在CO2浓度为800 μmol·mol-1时最大值为21.61 μmolCO2·m-2·s-1,对照‘109’柳在CO2浓度为800 μmol·mol-1时最大值为17.40 μmolCO2·m-2·s-1。
4.6 CO2 线性回归拟合
在CO2浓度Ci<200 μmol·mol-1时,对该段曲线进行线性回归拟合,并做出回归方程Pn=αCi+Rp。当回归方程Pn=Pmax时,可以计算出CO2饱和点(CSP)数值;当回归方程Pn=0时,可以计算出CO2补偿点(CCP)数值;回归方程斜率即为羧化效率(CE),结果见表4。

表4 CO2 线性回归拟合方程、CSP、CCP 和CE 值Tab.4 CO2 linear regression fitted equation,CSP、CCP and CE value
从表4 中可以看出,模型所有系数Sig <0.01,说明方程通过F 检验,线性回归分析有效;R2的数值都在0.99 以上,说明高度拟合,可以真实反映光响应曲线的走势。CO2饱和点在417.13 ~479.54 μmol·mol-1之间,竹柳无性系2CO2饱和点为479.54 μmol·mol-1,是模型中最大值;竹柳无性系3CO2饱和点为417.13 μmol·mol-1,是模型中最小值。CO2补偿点在132.79 ~156.82 μmol·mol-1之间,对照‘109’柳CO2补偿点为156.82 μmol·mol-1,是模型中最大值,竹柳无性系3CO2补偿点为132.79 μmol·mol-1,是模型中最小值。羧化效率竹柳无性系1 最大,为0.085;‘109’柳最小,为0.061。

图3 竹柳无性系1 净光合速率(Pn)日变化Fig.3 The net photosynthetic rate diurnal variation of Salix‘Zhuliu’clone 1
4.7 净光合速率日变化
竹柳无性系1 净光合速率日变化曲线如图3 所示。净光合速率(Pn)呈现双峰模式,在11:00 出现最大值15.70 μmolCO2·m-2·s-1,11:00 ~13:00 出现光合午休现象数值逐步下降,13:00 ~15:00 再次上升,15:00 出现第二个峰值12.90 μmolCO2·m-2·s-1。从多项式趋势线上看,竹柳无性系1 从9:00 开始Pn值逐步趋于饱和状态直到15:00 结束。
竹柳无性系2 净光合速率日变化曲线如图4 所示,净光合速率(Pn)呈现双峰模式,在10:00出现第一个峰值18.47 μmolCO2·m-2·s-1,10:00 ~11:00 出现光合午休现象,数值逐步下降;11:00 ~13:00 再次上升,13:00 出现第二个峰值19.13 μmolCO2·m-2·s-1。从多项式趋势线上看,竹柳无性系1 从9:00 开始Pn值逐步趋于饱和状态直到15:00 时结束。

图 4 竹柳无性系2 净光合速率(Pn)日变化Fig.4 The net photosynthetic rate diurnal variation of Salix‘Zhuliu’clone 2

图5 竹柳无性系3 净光合速率(Pn)日变化Fig.5 The net photosynthetic rate diurnal variation of Salix‘Zhuliu’clone 3
竹柳无性系3 净光合速率日变化曲线如图5 所示,净光合速率(Pn)呈现三峰模式,在9:00出现第一个峰值16.47 μmolCO2·m-2·s-1,9:00 ~10:00 出现第一次光合午休现象,数值逐步下降;10:00 ~11:00 再次上升,11:00 出现第二个峰值16.57 μmolCO2·m-2·s-1,11:00 ~13:00出现第二次光合午休现象,数值逐步下降;13:00 ~15:00 再次上升,15:00 出现第三个峰值16.40 μmolCO2·m-2·s-1。从多项式趋势线上看,竹柳无性系1 从9:00 开始Pn值逐步趋于饱和状态直到15:00 结束。
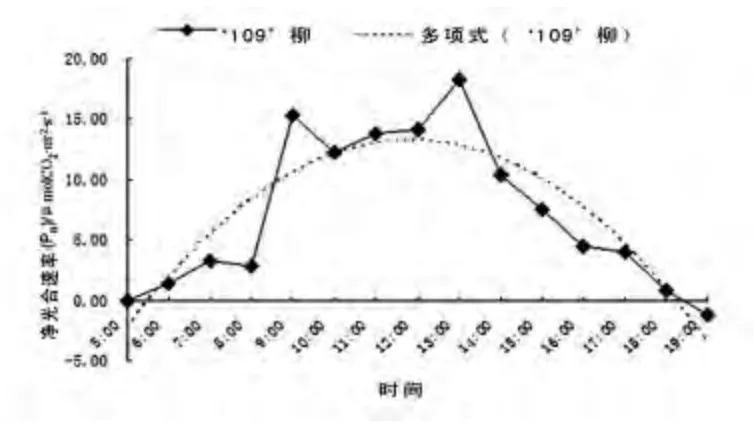
图6 ‘109’柳净光合速率(Pn)日变化Fig.6 The net photosynthetic rate diurnal variation of Salix בChuibao109’
对照‘109’柳净光合速率日变化曲线如图6 所示,净光合速率(Pn)呈现双峰模式,在9:00出现第一个峰值15.23 μmolCO2·m-2·s-1,9:00 ~10:00 出现第一次光合午休现象,数值逐步下降;10:00 ~13:00 再次上升,13:00 出现第二个峰值18.23 μmolCO2·m-2·s-1。从多项式趋势线上看,竹柳无性系1 从9:00 开始Pn值逐步趋于饱和状态直到14:00 结束。
竹柳3 个无性系与对照‘109’柳光合速率日变化峰值和趋饱和时间见表5,竹柳无性系3有3 个峰值,竹柳无性系2、竹柳无性系1 和对照‘109’柳只有2 个峰值。从峰值出现时间来看,竹柳无性系3 与对照‘109’柳第一峰值出现的时间最早,其次是竹柳无性系2,最后是竹柳无性系1;竹柳无性系3 第二峰值出现最早,竹柳无性系1 第二峰值出现最晚,竹柳无性系2 与对照‘109’柳第二峰值出现的时间居中。从趋饱和时间来看,竹柳3 个无性系趋饱和时间都是6 h,而对照‘109’柳趋饱和时间只有5 h。

表5 竹柳3 个无性系与‘109’柳的光合速率对照Tab.5 The photosynthetic rate comparison of three Salix‘Zhuliu’clone and Salix בChuibao109’
5 结论与讨论
光合作用光响应过程试验中,竹柳3 个无性系最大光合速率都明显大于对照‘109’柳最大光合速率,说明竹柳在强光作用下合成有效物质能力更强;而对照‘109’柳暗呼吸速率则明显大于竹柳3 个无性系,说明在维持植物正常生长发育过程中‘109’柳对物质和能量具有最高利用率,在整个生长过程中‘109’柳克服不良外界条件能力更强。从LSP、LCP 结果来看,竹柳3 个无性系光饱和点明显高于‘109’柳,而光补偿点却明显低于‘109’柳,可以说明竹柳不需要很强光照就开始合成干物质,在高光强条件下竹柳合成干物质能力也明显高于‘109’柳。
生长快的植物呼吸速率就高,生长慢的植物呼吸速率就低。光合作用CO2响应过程试验中,表 观 呼 吸 速 率(RP)在 9. 842 ~12.682 μmol·m-2·s-1之间,竹柳无性系1 为最大值,对照‘109’柳为最小值。竹柳与‘109’柳净光合速率随着CO2浓度上升而缓慢上升,达到最大值之后净光合速率随着CO2浓度增加而快速下降。达到最大值时顺序为:竹柳无性系2 >竹柳无性系1 >竹柳无性系3 >对照‘109’柳。竹柳3 个无性系呼吸速率都明显大于‘109’柳呼吸速率,说明竹柳生长速度比‘109’柳生长速度更快。CO2饱和点在417.13~479.54 μmol·mol-1之间,竹柳无性系2 是模型中最大值,竹柳无性系3 是模型中最小值。CO2补偿点在132.79 ~156.82 μmol·mol-1之间,对照‘109’柳是模型中最大值,竹柳无性系3 是模型中最小值。从CCP 的结果来看,竹柳3 个无性系明显低于‘109’柳值,说明竹柳光合效率更高。羧化效率竹柳无性系1 最大,‘109’柳最小。从CE 结果来看,竹柳3 个无性系都明显高于‘109’柳,说明竹柳光能利用率高于‘109’柳。
净光合速率(Pn)日变化试验中,竹柳无性系3 有3 个峰值,竹柳无性系2、竹柳无性系1和对照‘109’柳只有2 个峰值,竹柳3 个无性系的Pn趋饱和时间为6 h,‘109’柳的Pn趋饱和时间为5 h,也说明竹柳光合利用率明显高于‘109’柳。
在非线性回归模型选择上,光响应曲线和参数试用了比较常用的光合作用响应模型,包括指数模型、抛物线模型、直角双曲线模型、非直角双曲线模型和直角双曲线修正模型,最后确定光响应曲线和参数用非直角双曲线模型;CO2响应曲线和参数使用直角双曲线修正模型。在分析竹柳和‘109’柳光响应曲线和参数上,使用非直角双曲线模型拟合性最好,模型结果与实测数值基本一致;CO2响应曲线和参数上,使用直角双曲线修正模型拟合性最好,模型结果与实测数值也基本一致。
对竹柳3 个无性系和对照‘109’柳生长性状实测结果是:竹柳胸径、树高生长量明显大于‘109’柳,但是抗性却不如‘109’柳。从光合作用机理角度分析与实测结果完全一致。
[1]叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356-1361.
[2]陈根云,俞冠路,陈悦,等.光合作用对光和二氧化碳响应的观测方法探讨[J].植物生理与分子生物学学报,2006,32(6):691-696.
[3]刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J]. 中国农学通报,2005,21(8):76-79.
[4]叶子飘,高峻.丹参羧化效率在其CO2补偿点附近的变化[J].西北农林科技大学学报:自然科学版,2008,36(5):160-164.
[5]叶子飘,高峻.光响应和CO2响应新模型在丹参中的应用[J].西北农林科技大学学报:自然科学版,2009,37(1):129-134.
[6]叶子飘,王健林.植物光合-光响应模型的比较分析[J].井冈山学院学报:自然科学版,2009,30(4):9-13.
[7]高峻,孟平,吴斌,等.杏-丹参林药复合系统中丹参光合和蒸腾特性的研究[J].北京林业大学学报,2006,28(2):64-67.
[8]张弥,吴家兵,关德新,等.长白山阔叶红松林主要树种光合作用的光响应曲线[J].应用生态学报,2006,17(9):1575-1578.
[9]陆佩玲,于强,罗毅,等.冬小麦光合作用的光响应曲线的拟合[J].中国农业气象,2001,22(2):12-14.
[10]黄玉清,王晓英,陆树华,等.岩溶石漠化治理优良先锋植物种类光合、蒸腾及水分利用效率的初步研究[J].广西植物,2006,26(2):171-177.
[11]叶子飘,高峻.低光强时丹参叶片的光合速率与光强的关系[J].井冈山学院学报,2007,28(8):47-50.
[12]胡文海,胡雪华,曾建军,等.干旱胁迫对2 个辣椒品种光合特性的影响[J].华中农业大学学报,2008,27(6):776-781.
[13]夏江宝,张光灿,刘京涛,等.美国凌霄光合生理参数对水分与光照的响应[J].北京林业大学学报,2008,30(5):13-18.
[14]张淑勇,张光灿,陈建,等.土壤水分对五叶爬山虎光合与蒸腾作用的影响[J].中国水土保持科学,2006,4(4):62-66.
[15]张昆,万勇善,刘风珍,等.花生幼苗光合特性对弱光的响应[J].应用生态学报,2009,20(12):2989-2995.
[16]夏江宝,张光灿,刘刚,等.不同土壤水分条件下紫藤叶片生理参数的光响应[J].应用生态学报,2007,18(1):30-34.
[17]李德全,高辉远,孟庆伟.植物生理学[M].北京:中国农业科学技术出版社,2004.
[18]李小磊,张光灿,周泽福,等.黄土丘陵区不同土壤水分下核桃叶片水分利用效率的光响应[J].中国水土保持科学,2005,3(1):43-47.
[19]张光灿,刘霞,贺康宁.黄土半干旱区刺槐和侧柏林地土壤水分有效性及生产力分级研究[J].应用生态学报,2003,14(6):858-862.
[20]陈新军,张光灿,周泽福,等.黄土丘陵区紫丁香叶片气体交换参数的日变化及光响应[J].中国水土保持科学,2004,2(4):102-107.
