牛Wif1基因印记及甲基化调控印记的分子机制研究
2015-09-17吴茜红张明月杨文志吴国江李世杰
吴茜红,张明月,杨文志,吴国江,李世杰
(河北农业大学生命科学学院,保定071000)
基因组印记(Genomic imprinting)是一种非孟德尔遗传现象,是由于来自亲本双方的等位基因不对称表观遗传修饰而造成的,基因表达具有亲本特异性[1]。DNA甲基化是基因组印记的一种主要调控方式。基因组印记与生物进化、动物生长发育以及肿瘤发生都有密切关系。印记基因的研究主要集中在人类和鼠中,被鉴定的印记基因已经超过220个(http://igc.otago.ac.nz/home.html);而在牛中,由于缺少序列及多态性信息,被鉴定的印记基因只有20个。
Wnt抑制因子1(Wnt inhibitory factor 1,Wif1)是无翼信号通路(Wingless-type,Wnt)的胞外抑制蛋白,可直接和 Wnt蛋白结合,从而抑制Wnt信号的传导。Wnt信号通路具有传递生长刺激信号的作用,该通路的异常激活可引起细胞异常增殖、分化而导致肿瘤的发生[2]。Wif1基因首先在人的视网膜组织中发现,在人的胚胎滋养层中表现为一个父源表达的印记基因[3-4]。定位于人12号染色体。而在牛中,其印记状态和调控的分子机理还没有被研究。本研究在分析Wif1基因在牛不同组织中印记状态的基础上,对Wif1基因启动子区等位基因特异的甲基化状态进行了分析,以期为丰富牛基因组印记及揭示Wif1基因印记的调控分子机制提供依据。
1 材料与方法
1.1 试验材料
32头成年的荷斯坦奶牛(Holstein)的组织样本,包括心、肝、脾、肺、肾、骨骼肌、皮下脂肪等组织采自当地屠宰场,装入编号的采样袋中,立即投入液氮,-70℃冰箱保存,以备后续试验用。
1.2 RNA的提取及反转录
利用TRIzol试剂盒提取3头牛的心、肝、脾、肺、肾、肌肉和脂肪组织的总RNA,用无RNA酶的DNase-Ⅰ去除可能的DNA污染,-70℃保存备用。利用TransGen反转录试剂盒(TransGen,China)进行cDNA的合成。20μL反应体系中含有大约4μg的RNA,反转录反应程序按厂家提供的产品说明书进行,体系加好后轻轻混匀,65℃ 变性10min,打开RNA的二级结构,然后在每个反应体系中加入1μL的 EasyScriptTM RT/RI Enzyme Mix,42℃孵育30min,70℃保温15min,终止反应,cDNA于-20℃保存待用。
1.3 Wif1基因在不同组织中的表达谱分析
RT-PCR用来检测Wif1基因在牛组织中的表达模式。依照Wif1基因序列(GenBank accession No.NM_001075996.1),设计牛Wif1基因的特异性引 物 Wif-1F (5′-CCAACAAATGCCAGTG-3′)和 Wif-1R (5′-TCAAGCCAATGCCAAC-3′)用 于扩增812bp跨内含子的片段。利用GAPDH(GenBank accession No.BTU85042)跨内含子的引物 Gapdh-F(5′-GCACAGTCAAGGCAGAGAAC-3′)和 Gapdh-R(5′-GCGTGGACAGTGGTCATAAG-3′)扩增367bp大小的片段作为内参。反应体系为25μL:上下游引物(10μmol·L-1)各1μL,0.5μL cDNA 模板,10μL 双蒸水和12.5μL ES Tap MasterMix(CWBio,China)。扩增条件:94℃预变性5min;94℃变性30s,44℃退火30s,72℃延伸35s,35个循环;72℃总延伸10min。扩增产物用1%琼脂糖凝胶电泳进行检测,胶回收后送华大基因公司测序。
1.4 杂合子个体筛选
利用苯仿抽提法提取32头荷斯坦奶牛肝组织的 DNA。 引 物 Wif-2F (5′-CCCACCTGAATCCAAT-3′)和 Wif-2R (5′-TCAAGCCAATGCCAAC-3′)扩增667bp DNA片段,通过PCR产物直接测序法寻找SNP,筛选杂合子。除0.5μL DNA作为模板外,其他PCR反应体系同1.3RT-PCR。扩增条件:94℃预变性5min;94℃变性30s,44℃退火30s,72℃延伸30s,35个循环;72℃总延伸10min。PCR产物纯化后送华大基因公司测序。
1.5 Wif1基因的印记状态分析
被鉴定为杂合子的动物用于等位基因特异的表达状态分析。提取杂合子牛组织的RNA,反转录成cDNA、RNA提取和反转录程序同1.2,用得到的cDNA为模板,引物 Wif-1F和 Wif-1R扩增目的片段,将PCR产物胶回收后直接进行测序。
1.6 Wif1基因启动子区CpG岛预测及杂合子筛选
通过 MethPrimer 在 线 软 件 (http://www.urogene.org/methprimer),预测牛Wif1基因第一外显子及上游1kb序列上的CpG岛(Criteria used:Island size>100,GC Percent>50.0,Obs/Exp>0.6)。为了寻找启动子区的杂合位点,设计引物 Wif1-3F(5′-AACCCTCTGACCGTTGTG-3′)和 Wif1-3R (5′-GGCATTTGGGAAGAAGTAG-3′)扩增32头牛DNA。PCR反应体系为25μL:0.5μL DNA模板,上下游引物(10μmol·L-1)各1 μL,10μL 双蒸水和12.5μL ES Tap MasterMix(CWBio,China)。扩增条件:94℃预变性5min;94℃30s,52℃30s,72℃30s,共35个循环;72℃延伸10min。胶回收PCR产物后直接进行测序。得到的杂合子用于等位基因特异的甲基化分析。
1.7 亚硫酸氢盐处理、引物设计及PCR扩增
按照DNA甲基化试剂盒(Zymo,USA)说明书,将1.6中筛选到的杂合子牛的约500ng肺和肝组织DNA进行亚硫酸氢盐处理。利用在线软件(http://www.urogene.org/methprimer),针 对Wif1基因亚硫酸氢盐转换后的DNA序列设计甲基 化 特 异 性 引 物 (BSP)Wif1-MF(5′-TTGGTTAGAGGTAGCGTAAGT-3′)和 Wif1-MR (5′-AACCTCTTAACTCTAATACGAAT-3′)扩增Wif1基因启动子区410bp片段。反应体系为25μL:1μL亚硫酸氢盐转换后的DNA模板,上下游引物(10 μmol·L-1)各1μL,9.5μL双蒸水和12.5μL ES Tap MasterMix(CWBio,China)。扩增条件:94℃预变性5min;94℃30s,52℃30s,72℃30s,共35个循环;72℃延伸10min。第一轮PCR产物稀释100倍后,取1μL作为第二轮模板。第二轮PCR的引物、体系和扩增条件与第一轮相同。将第二轮PCR产物在1.5%琼脂糖凝胶中电泳检测。
1.8 克隆、DNA测序和数据分析
利用DNA 回收试剂盒(Cwbio,China)对1.7中第二轮扩增产物进行胶回收,纯化后的片段与pMD19-T载体连接,转化后涂板培养。每一个体各挑取18~22个单一的阳性克隆送华大科技公司测序。计算每一个体中mCpG占全部CpG的百分比。SPSS软件分析等位基因间甲基化水平的差异显著性。
2 结 果
2.1 Wif1基因在不同组织中的表达
RT-PCR用于分析Wif1基因在牛7个组织中的表达,包括心、肝、脾、肺、肾、肌肉和脂肪。引物Wif-1F和 Wif-1R扩增来自3头牛组织的RNA,GAPDH基因作为内参。结果在7个被检测的组织中,Wif1基因均扩增得到812bp大小的条带(图1),测序验证为目的产物,表明Wif1基因在被检测的组织中均表达。

图1 牛Wif1基因各个组织中的RT-PCR结果Fig.1 RT-PCR results of Wif1gene from different tissues of cattle
2.2 Wif1基因杂合子个体筛选
用位于Wif1基因外显子8上的引物 Wif-2F和Wif-2R扩增来自32头牛肝组织的DNA,得到667bp的DNA片段,PCR产物回收后直接测序(图2),发现一个A/C杂合位点,位于该基因的第1 759个核苷酸处(c.1759A>C),鉴定的杂合子牛用于印记状态分析。
2.3 Wif1基因印记状态分析
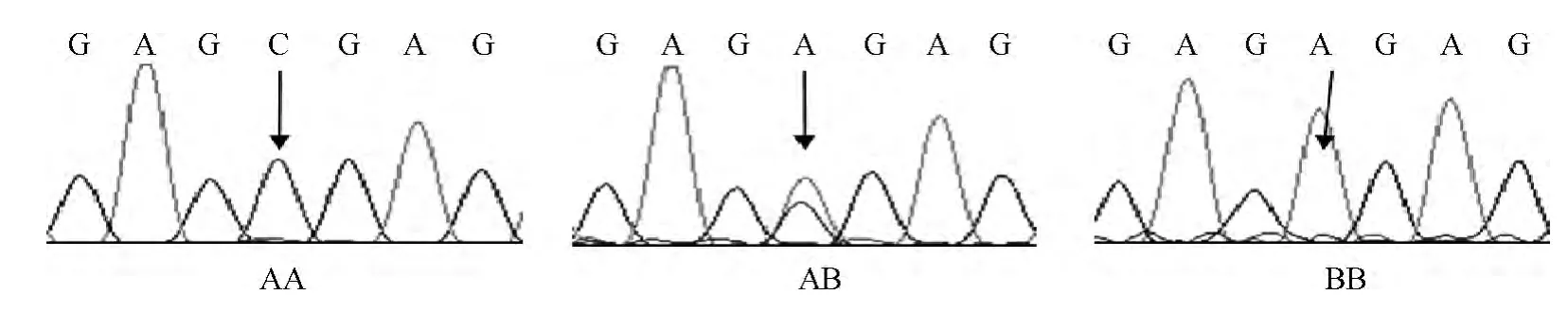
图2 牛DNA测序法确定Wif1基因的杂合子个体Fig.2 Identification of an SNP (A/C)in the bovine Wif1gene by sequencing
通过比较杂合子动物中g.1759A>C SNP位点在DNA PCR和RT-PCR产物直接测序的峰图来确定印记基因的表达状态。提取3头g.1 7 5 9A>C SNP位点为杂合的动物的组织RNA,RT-PCR产物直接测序,SNP位点附近峰图见图3,发现Wif1基因只有在肺中为单等位基因表达(单峰),说明Wif1基因在肺中是印记的;而在心、肝、脾、肾、肌肉、脂肪中为双等位基因表达(双峰),说明Wif1基因在检测的6个组织中是非印迹的。
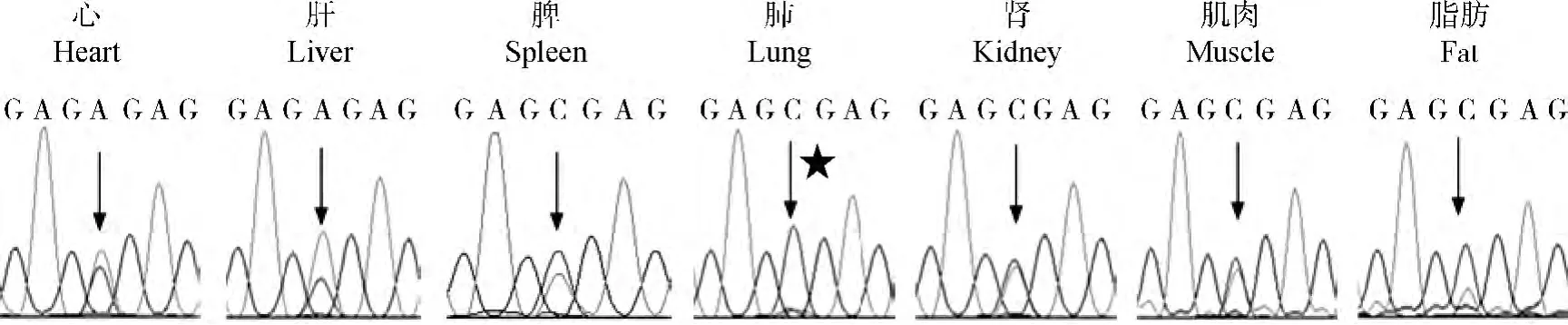
图3 测序法分析Wif1基因等位基因特异性的表达模式Fig.3 Allele-specific expression analysis of Wif1gene in cattle by sequencing directly
2.4 Wif1等位基因特异的甲基化状态分析
为了确定DNA甲基化在Wif1基因组织特异性印记中的可能作用,分析了Wif1基因启动子区在单等位基因表达的组织(肺)和双等位基因表达的组织(肝)中的等位基因特异的甲基化状态。用位于Wif1基因启动子区的引物 Wif-3F和 Wif-3R扩增来自32头牛肝组织的DNA,PCR产物回收后直接测序,发现一个A/G杂合位点,位于该基因的第115个核苷酸处(g.115A>G),g.115A>G SNP位点用于区分等位基因的两条链。用甲基化特异引物(Wif1-MF和 Wif1-MR)扩增包括g.115A>G在内的Wif1基因启动子416bp的区域,包括了29个CpG位点,软件预测每个CpG位点上可能存在的转录因子结合的位点,发现其中14个CpG位点(第1、2、3、4、5、6、7、8、10、11、15、18、24和25)上存在的7种转录因子结合的位点(图4),即ATF(TGACGT)、CREB(KWCGTCA)、GCF (SCGSSSC)、AP2(CCCMNSSS)、SP1 (GGCGGG)、HSV_IE_repeat(GCGGAA)、EIIaE-A(AAGGGCGC)。
分析3头杂合子牛(7号、25号和32号)肝和肺组织中等位基因特异的甲基化状态。根据在SNP位点的碱基命名来自亲本等位基因的两条链,分别为G-链和A-链(图5)。结果发现,在单等位基因表达的肺中,两条亲本链G-链和A-链的平均甲基化水平分别为38%和41%,差异不显著(P>0.05);在双等位基因表达的肝中,G-链(42%)和 A-链(38%)的平均甲基化水平也无显著差异(P>0.05)。进一步对肺和肝中每个CpG位点两条亲本链G-链和A-链之间的平均甲基化率进行差异显著性分析 (图6),发现在肺中有10个CpG位点A链与G链甲基化率存在显著差异,其中第2、7、13、17个CpG位点差异极显著(P<0.01),第1、6、9、19、23、29个CpG位点的差异显著(P<0.05);在双等位基因表达的肝中,有4个CpG位点A链与G链甲基化率存在显著差异,其中第7个CpG位点差异极显著(P<0.01),第13、23、和29个CpG位点差异显著(P<0.05)。与肝相比,肺中8个CpG位点(第1、2、6、9、17、19、23和29)两条亲本链甲基化率存在显著差异,其中3个CpG位点(第1、2和6)位于转录因子的结合位点上,由于单个CpG位点的甲基化变化会影响转录因子的结合,推测这3个CpG位点的甲基化可能参与调控Wif1基因的组织特异性印记。
3 讨 论
WIF1是近年来发现的肿瘤抑制因子,通过与Wnt蛋白直接结合抑制Wnt信号通路,从而抑制癌细胞的干性并且促使癌细胞老化。本研究分析了Wif1基因在牛7个组织中mRNA转录情况、印记状态以及启动子区DNA甲基化的状态。
Wif1基因的表达因物种、发育阶段而有差异。J.C.Hsieh等[3]在蟾蜍胚胎发育的神经胚期检测到了Wif1基因的mRNA,而在蟾蜍和斑马鱼的胚胎原肠胚期未能检测到Wif1基因的表达。Wif1基因在成年小鼠的心脏和肺脏中强表达,而在脑和眼中表达水平较低。D.D.Hunter等[5]发现在人的视网膜、软骨、肺和大脑内Wif1显著表达;Wif1分布在小鼠神经系统的很多不同区域,其中丘脑和嗅球内表达显著。C.Wissmann等[6]使用多克隆抗体的免疫组织化学分析显示,在前列腺、乳腺、肺和膀胱的正常上皮细胞核周细胞质Wif1高表达。本研究以RT-PCR方法分析了牛中Wif1基因在心、肝、脾、肺、肾、肌肉和脂肪组织中mRNA的转录情况,发现Wif1基因在7个被测组织中均表达。研究结果表明,Wif1基因在多个物种中广泛表达于多种组织中。

图4 甲基化分析的Wif1基因启动子区及预测的每个CpG位点上的转录因子Fig.4 The promoter region of Wif1gene analysed by bisulfite sequencing and the transcription factors predicted on each CpG sites
大部分印记基因在哺乳动物的胚胎和胎盘发育中起重要作用[7-10]。由于胎盘是最早印记的组织之一[11]。Wif1基因是在研究人胎盘肿瘤抑制基因时,被发现在人妊娠前3月胎盘滋养层细胞中表现为父源表达的印记基因[4]。关于Wif1基因在其他的物种和组织中的印记状态尚未报道。基于SNP位点的RT-PCR产物直接测序法是一种分析印记基因的有效方法,前期研究中应用此方法确定了Gtl2和Meg8在牛中的印记状态[12-13]。本研究发现Wif1基因在牛中的印记具有组织特异性,在被分析的7个组织中,Wif1基因只在牛的肺中是印记的,表现为单等位基因表达;而在心、肝、脾、肾、肌肉、脂肪中表现为双等位基因表达。表明Wif1基因的基因组印记具有物种特异性和组织特异性。

图5 3头杂合子牛(7、25和32号)肺和肝启动子区甲基化状态和甲基化水平Fig.5 Methylation level and status of promoter region in lung and liver tissues of 3heterozygous cattle(7,25,32)
DNA甲基化作为一种重要的表观遗传修饰方式,通过改变转录因子与DNA的结合,从而调控基因的组织特异表达、基因组印记和动物的正常发育[14-15]。Wif1基因启动子区域的甲基化调控Wif1基因的表达,大量的研究证明Wif1基因的启动子高甲基化与消化系统恶性肿瘤、鼻咽癌、膀胱癌、乳腺癌等恶性肿瘤有密切关系[16-22]。5-氮-2′-脱氧胞苷(5-aza-dC)是一种甲基化抑制剂,在 DNA 复制过程中,5-aza-dC与DNA甲基转移酶形成一种共价复合物,抑制甲基转移酶的甲基转移活性,从而实现去甲基化功能。在膀胱癌等细胞系中加入5-aza-dC,可使Wif1 基因的 表 达恢复正常[23-26]。本研究中,为了探究甲基化在Wif1基因的组织特异性印记中可能发挥的作用,分析了启动子区29个CpG位点的甲基化状态,发现肺中9个CpG位点的甲基化水平发生改变。对其转录因子结合位点的预测发现,该区域存在多种转录因子结合位点,这些转录因子调控着基因转录和转录表达水平的高低。如CREB是一种与细胞周期调控及记忆形成有关的核转录增强因子[27-28]。Sp1是广泛和系统的生长调控因子之一,它参与了细胞生长和分化,对肿瘤的促进作用与其参与众多癌基因、抑癌基因、细胞周期调控分子、生长相关信号转导通路分子的调节密切相关[29]。转录因子 AP-2家族是一类DNA 结合蛋白,以二聚体形式结合到DNA丰富GC的元件上,在基因表达调控、细胞分化、胚胎发育及肿瘤发生中起着极为重要的作用。由于单个CpG的甲基化可能会影响转录因子的结合,所以差异甲基化的CpG位点可能参与了Wif1基因的组织特异性印记。

图6 肺和肝中每个CpG位点的甲基化率Fig.6 Methylation rate of each CpG site in lung and liver tissues
(References):
[1]KHATIB H,ZAITOUN I,KIM E S.Comparative analysis of sequence characteristics of imprinted genes in human,mouse and cattle[J].MammGenome,2007,18(6-7):538-547.
[2]POLAKIS P.Wnt signaling and cancer[J].Genes Dev,2000,14(15):1837-1851.
[3]HSIEH J C,KODJACHIAN L,REBBERT M L,et al.A new secreted protein that binds to Wnt proteins and inhibits their activities[J].Nature,1999,398(6726):431-436.
[4]GUILLERET I,OSERHELD M C,BRAUNSCHWEIG R,et al.Imprinting of tumor-suppressor genes in human placenta[J].Epigenetics,2009,4(1):62-68.
[5]HUNTER D D,ZHANG M,FERGUSON J W,et al.The extracellular matrix component WIF-1is expressed during,and can modulate,retinal development[J].MolCellNeurosci,2004,27(4):477-488.
[6]WISSMANN C,WILD P J,KAISER S,et al.WIF1,a component of the Wnt pathway,is down-regulated in prostate,breast,lung,and bladder cancer[J].J Pathol,2003,201(2):204-212.
[7]YOUNG L E,SINCLAIR K D,WILMUT I.Large offspring syndrome in cattle and sheep[J].RevRepord,1998,3(3):155-163.
[8]HILL J R,BURGHARDT R C,JONES K,et al.Evidence for placental abnormality as the major cause of mortality in first-trimester somatic cell cloned bovine fetuses[J].BiolReprod,2000,63(6):1787-1794.
[9]FARIN C E,FRAIN P W,PIEDRAHITA J A.Development of fetuses from invitro-produced and cloned bovine embryos[J].JAnimSci,2004,82:E53-62.
[10]ZAITOUN I,KHATIB H.Comparative genomic imprinting and expression analysis of six cattle genes[J].JAnimSci,2008,86(1):25-32.
[11]MAEDA N,HAYASHIZAKI Y.Genome-wide survey of imprinted genes[J].CytogenetGenomeRes,2006,113(1-4):144-152.
[12]WANG J C,LI D J,DU W H,et al.Genomic imprinting of GTL2in somatic cell nuclear transfer cattle[J].ProgBiochemBiophys,2008,35(10):1183-1187.
[13]HOU X H,LI D J,SU H,et al.Molecular cloning,expression,and imprinting status of maternally expressed gene 8 (Meg8)in dairy cattle[J].Genetika,2011,47(8):1120-1125.
[14]SHIOTA K,KOGO Y,OHGANE J,et al.Epigenetic marks by DNA methylation specific to stem,germ and somatic cells in mice[J].GenesCells,2002,7(9):961-969.
[15]LORINCZ M C,DICKERSON D R,SCHMITT M,et al.Intragenic DNA methylation alters chromatin structure and elongation efficiency in mammalian cells[J].NatStructMolBiol,2004,11(11):1068-1075.
[16]DING Z,QIAN Y B,ZHU LX,et al.Promoter methylation and mRNA expression of DKK-3and WIF-1in hepatocellular carcinoma[J].WorldJGastroenterol,2009,15(21):2595-2601.
[17]HUANG L,LI M X,WANG L,et al.Prognostic value of Wnt inhibitory factor-1expression in hepatocellular carcinoma that is independent of gene methylation[J].TumourBiol,2011,32(1):233-240.
[18]FENDRI A,KHABR A,HADRI-GUIGA,et al.Epigenetic alteration of the Wnt inhibitory factor-1promoter is common and occurs in advanced stage of Tunisian nasopharyngeal carcinoma[J].CancerInvest,2010,28(9):896-903.
[19]LEE B B,LEE E J,JUNG E H,et al.Aberrant methylation of APC,MGMT,RASSF2A,and Wif-1genes in plasma as a biomarker for early detection of colorectal cancer[J].ClinCancerRes,2009,15(19):6185-6191.
[20]YOSHINO M,SUZUKI M,TIAN L,et al.Promoter hypermethylation of the p16and Wif-1genes as an independent prognostic marker in stage IA non-small cell lung cancers[J].IntJOncol,2009,35(5):1201-1209.
[21]KAWAKAMI K,HIRATA H,YAMAMURA S,et al.Functional significance of Wnt inhibitory factor-1 gene in kidney cancer[J].CancerRes,2009,69(22):8603-8610.
[22]URAKAMI S,SHIINA H,ENOKIDA H,et al.Epigenetic inactivation of Wnt inhibitory factor-1plays an important role in bladder cancer through aberrant canonical Wnt/beta-catenin signaling pathway[J].ClinCancerRes,2006,12(2):383-391.
[23]GAO Z,XU Z,HUNG M S,et al.Promoter demethylation of WIF-1by epigallocatechin-3- gallate in lung cancer cells[J].AnticancerRes,2009,29(6):2025-2030.
[24]BATRA S,SHI Y,KUCHENBECKER K M,et al.Wnt inhibitory factor-1,a Wnt antagonist,is silenced by promoter hypermethylation in malignant pleural mesothelioma[J].BiochemBiophysResCommun.2006,342(4):1228-1232.
[25]LIN YC,YOU L,XU Z,et al.Wnt signaling activation and WIF-1silencing in nasopharyngeal cancer cell lines[J].BiochemBiophysResCommun,2006,341(2):635-640.
[26]MAZIERES J,HE B,YOU L,et al.Wnt inhibitory factor-1is silenced by promoter hypermethylation in human lung cancer[J].CancerRes,2004,64(14):4717-4720.
[27]ZHANG C,WANG P.Progress in the reulation of transcription factor CREB on cell cycle[J].ChinJ CellBiol,2011,33(5):577-583.
[28]余瑞元,王燕峰,徐长法.CREB研究进展[J].中国生物工程杂志,2003,23(1):39-42.YU R Y,WANG Y F,XU C F.Progress in the studies on CREB[J].JournalofChineseBiotechnology,2003,23(1):39-42.(in Chinese)
[29]白 雪,邓 红.转录因子Sp1与肿瘤关系研究的新进展[J].浙江大学学报(医学版),2010,39(2):215-220.BAI X,DENG H.Research progress on relationship between transcription factor SP1and tumor[J].JournalofZhejiangUniversity(MedicalScience),2010,39(2):215-220.(in Chinese)
