弱光环境对罗勒幼苗叶片生长和叶绿素荧光特性的影响
2015-08-24林晗婧张秀丽孙广玉东北林业大学生命科学学院哈尔滨150040
林晗婧,田 野,张秀丽,孙广玉(东北林业大学生命科学学院,哈尔滨150040)
弱光环境对罗勒幼苗叶片生长和叶绿素荧光特性的影响
林晗婧,田野,张秀丽,孙广玉
(东北林业大学生命科学学院,哈尔滨150040)
以花色素苷含量不同的紫罗勒和大叶罗勒为对象,分别对其进行弱光和自然光处理30d后进行生长参数、色素含量和叶绿素荧光参数的测定,结果显示:弱光导致两种罗勒幼苗叶片比叶面积、叶面积、叶宽、叶长和株高显著增加,幼苗根长,地上、地下生物量和总生物量明显降低。经过弱光处理的叶片花色苷与总叶绿素比和花色苷含量均降低。总叶绿素含量、叶绿素a含量、叶绿素b含量和叶绿素a/b比值较自然光明显升高。在弱光下,两种罗勒幼苗叶片失活PSⅡ反应中心的热耗散量子产额(ФNF)降低,而基本的荧光量子产额和热耗散的量子产额(Фf,D)升高。说明,花色素苷可能在光破坏防御上发挥了作用。
罗勒;弱光;叶绿素荧光;花色素苷
Abstract∶Based onOcimum basilicumL.seedlings with different content of anthocyanins,this article studied on the growth parameters,pigment content and chlorophyll fluorescence parameters dealed with lowlight intensity and nature light intensity for 30 days.Finally,the specific leaf area,leaf area,blade width,leaf length,plant height of leaves ofOcimum basilicumL.seedlings increased significantly,however root length,aboveground biomass,underground biomass,total biomass of leaves decreased significantly with low light intensity.The content of anthocyanins and the ratio of anthocyanins and total chlorophyll ofOcimum basilicumL.seedlings with low light intensity was lower than the one with nature light intensity,however the content of total chlorophyll,chlorophyll a,chlorophyll b,the ratio ofchlorophyll a and b ofOcimum basilicumL.seedlings with lowlight intensity was higher. Dealed with lowlight intensity,the quantum yield of thermal dissipation in nonfunctional PSⅡ(ФNF)decreased,the basic quantum yield of fluorescence and thermal dissipation(Фf,D)increased in leaves ofOcimum basilicumL. seedlings.The result showed that anthocyanins mayhave the effect on the defense oflight damage.
Key words∶OcimumbasilicumL;lowlight density;chlorophyll fluorescence;anthocyanin
罗勒(Ocimum basilicum L.)为唇形科罗勒属一年生草本植物,原产于南洋群岛及非洲等地,遍布全球[1]。罗勒形态上呈扇形植株,枝繁叶茂,紫花成串,适宜盆栽,由于其具有能香化、净化空气、驱蚊等功效,而被誉为“天然香水瓶”[2]。由于罗勒是一种值得推广的天然保健、药食同源的资源植物,因而近年来市场需求不断增大,食用罗勒栽培面积亦呈增长趋势[3]。紫罗勒是罗勒的变种之一,其叶片富含大量的花色素苷,该类物质属于天然色素类抗氧化剂,可被应用于食品以及药品等领域[4],其主要分布于幼嫩叶片和衰老叶片中。影响花色素苷合成的因素主要有光、温度、水分以及叶片生长状态等[5],其中光强[6]直接决定了叶片中的色素含量。为此,本试验以紫罗勒和大叶罗勒幼苗为试验材料,利用叶绿素荧光技术研究了弱光条件下花色素苷对罗勒幼苗叶片光能吸收、利用和分配的影响,为揭示花色素苷如何在植物叶片光合作用中发挥效果提供参考。
1 材料与方法
1.1材料
试样紫罗勒叶片均为紫色;丁香罗勒叶片均为绿色。6月初播种,将种子均匀地种植到直径12cm、高15cm的培养钵中,每钵种植5粒种子,培养基质采用充分混匀的草炭土与蛭石,比例定为1∶1(v/v)。在温度25/23℃(光/暗)、正午最大光照强度1 200μmol·m-2· s-1、光周期12/12h(光/暗),75%左右相对湿度的温室中培养,定期进行苗期管理并浇水,待幼苗出土长至约10cm时,间苗处理,每钵保留生长相对一致的幼苗1株。
1.2处理方法
试验设计:一组为弱光处理,用市售透光率为50%的黑色遮阳网固定在铁架四周,每个品种罗勒15盆于铁架下;另一组为自然光处理,每个品种15盆于温室中,试验材料处理30d后进行生长参数、色素含量和叶绿素荧光参数的测定。
1.3测定项目和方法
叶面积的测量方法为称量纸质量法。用玻璃板把叶片压平,在白纸上画出叶片轮廓,剪下称取质量,并测量其叶宽和叶长。剪下1cm2纸,称出单位面积纸的质量,叶片面积公式为:叶片面积(cm2)=纸质量/单位面积纸质量(cm2),将叶片放入铝盒中杀青(105℃,30min)、烘干(60℃,30h)至恒重后称其质量即为干质量,比叶面积(cm2/g)=叶面积/干质量。测定其各待测株的株高后收获每培养钵内的植株,洗净根系表面的培养基质后测定根系长度,用吸水纸吸干根系表面的水分后,分别测定地上和地下部分的鲜重,然后分别将地上部和地下部放铝盒内杀青(105℃,30min)、烘干(60℃,30h)至恒重后称其生物量,计算平均每株的地上部生物量和地下部生物量、根冠比,其中根冠比=地下部生物量/地上部生物量。
根据无水乙醇法[7]测定叶片中叶绿素a、叶绿素b以及叶绿素总量含量,并参照Pirie[8]的方法测定叶片中花色素苷的含量。最后,计算花色素苷与叶绿素总量的比值。
根据Hu[9]的方法测定叶绿素荧光参数,并按张会慧[10]法分别计算出PSⅡ反应中心吸收光能的去向。
1.4数据处理和统计方法
本试验运用Excel和DPS软件对数据进行统计分析,所有试验数据均平均三次,结果表示为值±标准差(SE),并通过Design expert 8.0软件对比分析不同数据组间的差异。
2 结果与分析
2.1弱光对罗勒幼苗生长特性影响
由表1可以看出,弱光导致两种罗勒幼苗叶片比叶面积、叶面积、叶宽、叶长和株高均增加,其中大叶罗勒较紫罗勒增加幅度大,而两种罗勒幼苗根长、地上生物量、地下生物量和总生物量均明显降低,大叶罗勒幼苗叶片较紫罗勒降低幅度较小。在弱光下,紫罗勒幼苗叶片含水量较自然光无显著变化而大叶罗勒幼苗叶片是自然光下叶片含水量的2.3倍;紫罗勒幼苗根冠比较自然光显著降低而大叶罗勒幼苗变化不显著。
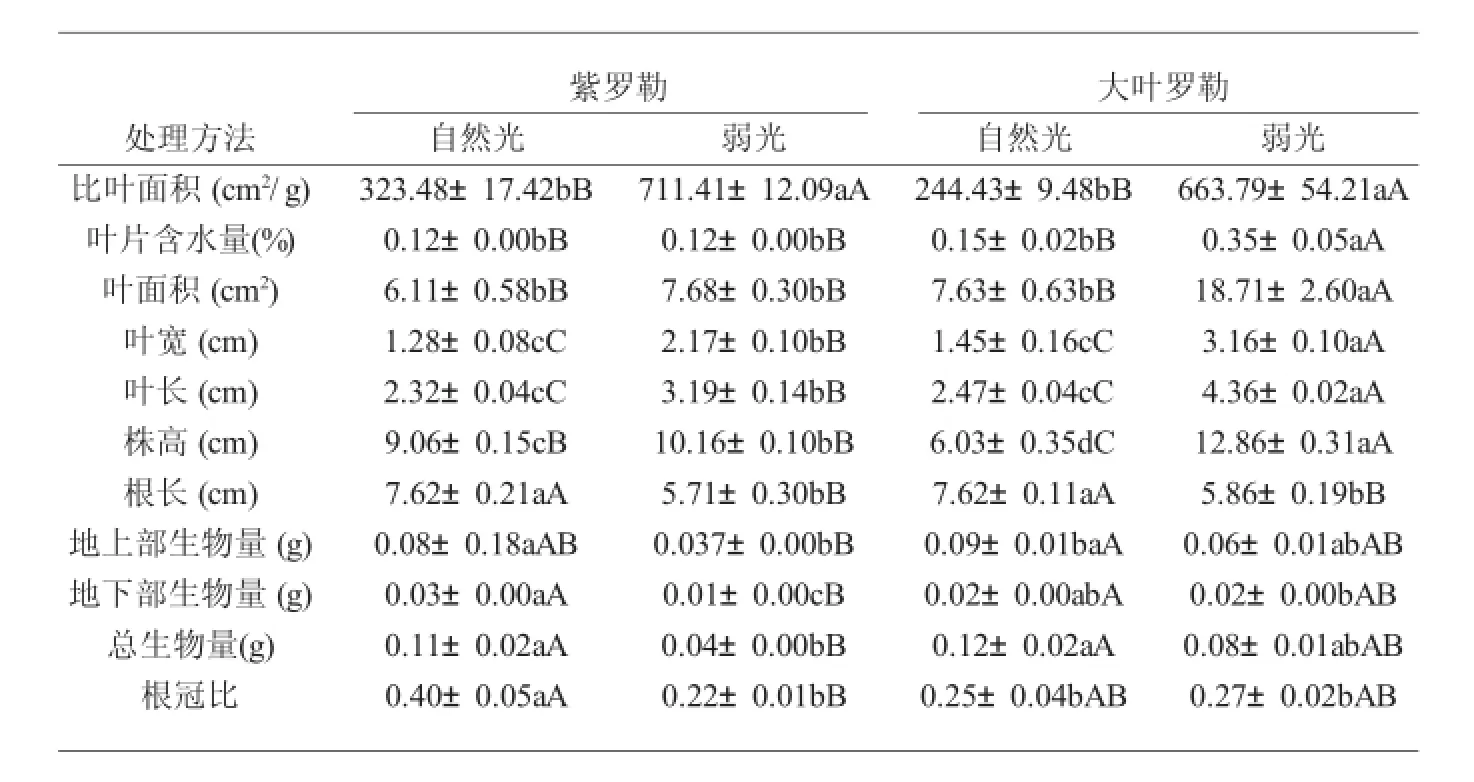
表1 弱光对罗勒幼苗生长特性的影响Tab.1 Effects of growth characteristics in leaves ofOcimum basilicumL.seedlings under low light intensity
2.2弱光对罗勒幼苗叶片叶绿素和花色素苷的影响
由表2可以看出,弱光导致紫罗勒幼苗叶片花色素苷含量和花色素苷与总叶绿素比值明显降低,但大叶罗勒变化不显著(P<0.05)。在弱光下,两种罗勒幼苗叶片总叶绿素含量、叶绿素a含量、叶绿素b含量和叶绿素a/b比值较自然光明显升高,并且大叶罗勒幼苗叶片总叶绿素含量、叶绿素a含量和叶绿素b含量分别为自然光的1.8倍、2倍和1.9倍。这与其他彩叶植物在弱光下形态变化、叶片光合色素及花色素苷含量变化一致。
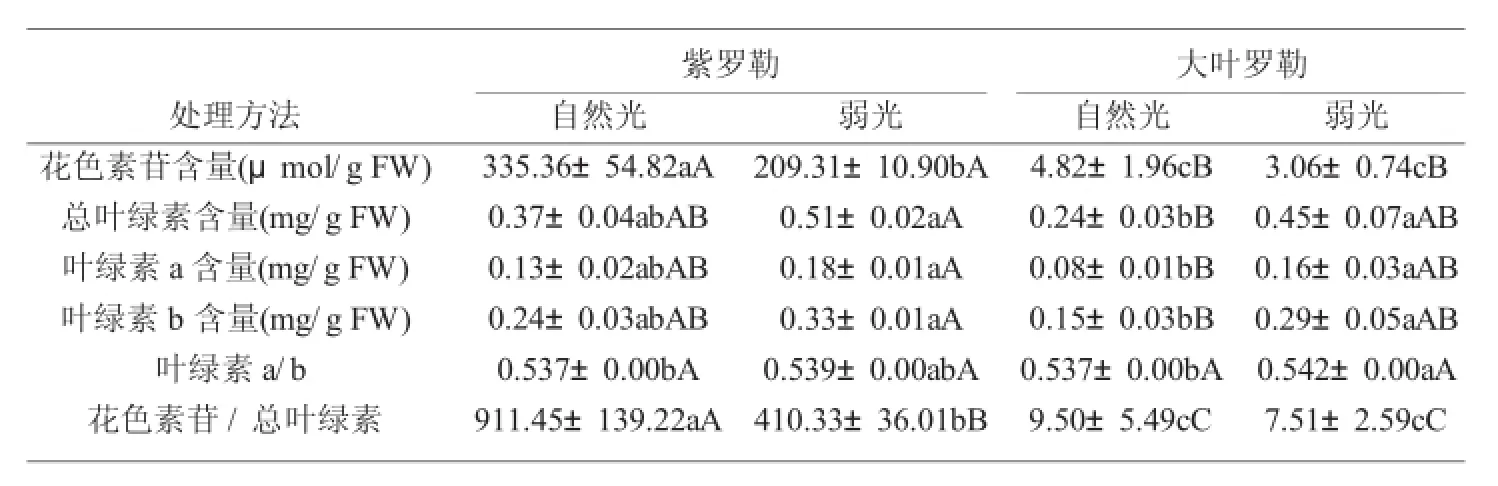
表2 弱光对罗勒幼苗叶片叶绿素和花色素苷的影响Tab.2 Effects of chlorophyll and anthocyanin contents in leaves ofOcimum basilicumL.seedlings under low light intensity
2.3弱光对罗勒幼苗叶片Fo、Fm、Fv/Fm和Fv/Fo的影响
由图1可以看出,在弱光下,紫罗勒幼苗叶片初始荧光(Fo)较自然光显著升高而大叶罗勒幼苗叶片降低;两种罗勒幼苗叶片最大荧光(Fm)较自然光均明显升高,但紫罗勒幼苗叶片升高幅度较大叶罗勒幼苗增大;大叶罗勒幼苗叶片最大光化学效率(Fv/Fm)和潜在光化学效率(Fv/Fo)较自然光均显著升高,但紫罗勒幼苗叶片Fv/Fm和Fv/Fo变化不显著。
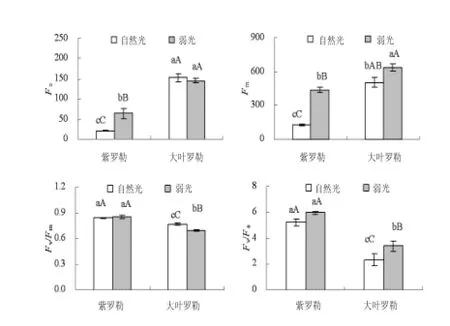
图1 弱光对罗勒幼苗叶片Fo、Fm、Fv/Fm和ETR的影响Fig.1 Effects of Fo,Fm,Fv/Fm and ETR in leaves ofOcimum basilicumL.seedlings under low light intensity
2.4弱光对罗勒幼苗叶片PSⅡ反应中心吸收光能分配参数的影响
由图2可以看出,紫罗勒幼苗叶片热耗散量子产额(ФNF)和用于光化学反应的量子产额(ФPSⅡ)大于大叶罗勒叶片,而基本的荧光量子产额和热耗散的量子产额(Фf,D)小于大叶罗勒叶片。弱光处理使紫罗勒叶片用于光化学反应的量子产额(ФPSⅡ)降低,依赖于类囊体膜两侧质子梯度和叶黄素循环的量子产额(ФNPQ)升高,而大叶罗勒叶片情况恰恰相反,(ФPSⅡ)升高而(ФNPQ)降低,在弱光下,两种罗勒幼苗叶片失活PSⅡ反应中心的热耗散量子产额(ФNF)降低,而(Фf,D)较自然光升高。
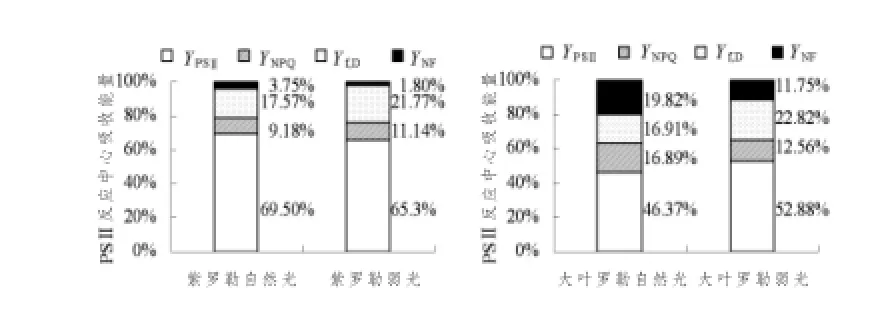
图2 弱光对罗勒幼苗叶片PSⅡ反应中心吸收光能分配参数的影响Fig.2 Effects of PSⅡenergy allocation pathways in leaves of Ocimum basilicumL.seedlings under low light intensity
3 讨论与结论
本试验中弱光导致两种罗勒幼苗叶片比叶面积、叶面积、叶宽、叶长和株高均增加,与之前对苹果、桃、黄瓜[11]、葡萄等的研究结果一致,该结果表明,一方面植物不同器官在形态发生方面对光的敏感性不同,另一方面植物能够根据生长环境光照条件进行主动适应。光合效率指标是以叶绿素荧光指数Fv/Fm表示的PSⅡ的光化学效率,因此,从花色素苷对罗勒叶片叶绿素荧光特性和能量分配角度分析,弱光下,紫罗勒幼苗叶片Fv/Fm和Fv/Fo变化不明显,这说明弱光并没有导致叶片PSⅡ反应中心的光化学活性降低;而大叶罗勒幼苗叶片Fv/Fm和Fv/Fo显著升高,这是由于叶片叶绿素含量升高,叶片捕光能力增强,但并未产生过剩的光能。弱光条件下大叶罗勒幼苗叶片Fo降低是由于PSⅡ反应中心的热耗散增加[12],而紫罗勒幼苗叶片Fo升高表示PSⅡ反应中心受到破坏或可逆失活[13]。本试验发现,弱光导致紫罗勒叶片用于光化学反应的量子产额(ФPSⅡ)降低,而大叶罗勒叶片ФPSⅡ升高,这可能是由于紫罗勒叶片中花色素苷的存在衰减或者反射部分光能,导致紫罗勒幼苗叶片光化学效率降低,而大叶罗勒幼苗叶片中花色素苷含量相对较少[14]。在弱光下,两种罗勒幼苗叶片失活PSⅡ反应中心的热耗散量子产额(ФNF)较自然光降低,基本的荧光量子产额和热耗散的量子产额(Фf,D)较自然光升高。增加了
热耗散所消耗的能量,从而提高了有活性的反应中心,防止反应中心的失活、降解。而紫罗勒幼苗叶片依赖于类囊体膜两侧质子梯度和叶黄素循环的量子产额(ФNPQ)升高,而大叶罗勒叶片ФNPQ降低。这说明在弱光下,紫罗勒幼苗叶片通过叶黄素循环以及热耗散等来维持反应中心的正常功能,而大叶罗勒幼苗叶片主要通过热耗散来维持反应中心的正常功能。通过以上结果可以看出,花色素苷可以保护紫罗勒幼苗叶片免受弱光胁迫,花色素苷可能在光破坏防御上发挥了作用。
[1]周爱芬.罗勒的光合特性研究[J].丽水学院学报,2011,33(2):42-44.
[2]吴冬乾,夏月明,朱玉萍.罗勒的实用栽培技术[J].上海蔬菜,2005,(02):1002-1469.
[3]祝丽香.罗勒的研究与开发应用[J].北方园艺,2005,(01):15-16.
[4]孙丽华,江月仙,王巧懿.天然抗氧化剂原花青素的保健功能及其应用[J].食品研究与开发,2004,(02):109-112.
[5]Jin S K,Byung H L,So H K,et al.Responses to environmental and chemical signals for anthocyanin biosynthesis in non-chlorophyllous corn(Zea mays L.)leaf[J].Journal of Plant Biology,February,2006,49(1)∶16-25.
[6]Oren-Shamir M.Does anthocyanin degradation play a significant role in determining pigment concentrationinplants?[J].Plant Science,2009,177(4)∶310-360.
[7]占丽平,鲁剑巍,杨娟,等.施肥对黑麦草生长和产量的影响[J].草业科学,2011,28(2):260-265.
[8]Pirie A,Mullins M G.Changes in anthocyanin and phenolics content of grapevine leaf and fruit tissues treated with sucrose,nitrate,and sbscisic acid[J]. Plant Physiology,1976,58(4)∶468-472.
[9]Hu Y B,Sun G Y,Wang X C.Induction characteristics andresponse of photosynthetic quantum conversion to changes in irradiance in mulberry plants[J].Journal of Plant Physiology,2007,164(8)∶959-968.
[10]张会慧,张秀丽,胡彦波,等.碱性盐胁迫对桑树幼苗叶片叶绿素荧光和激发能分配的影响[J].经济林研究,2012,30(1):6-12.
[11]战吉成,王利军,黄卫东.弱光环境下葡萄叶片的生长及其在强光下的光合特性[J].中国农业大学学报ISTIC PKU,2002,7(3):1007-4333.
[12]Demmig-Adams B,Adams W W III.Photoprotection and other responses of plants to high light stress[J].Annual Review of Plant Physiology and Plant Molecular Biology,1992,(43)∶599-626.
[13]李天来,刘玉凤,宋礼毓.夜间亚低温处理及其恢复对番茄叶片光抑制的影响[J].园艺学报,2008,(07):1003-1010.
[14]田野,张会慧,王娟,等.遮荫对紫叶李幼苗叶片花色素苷含量和光合能量分配的影响[J].中南林业科技大学学报,2013,(06):64-69.
Effects of growth characteristics and chlorophyll fluorescence characteristics in leaves ofOcimum basilicumL.seedlings under low light intensity
LINHan-jing,TIANYe,ZHANGXiu-li,SUNGuang-yu
(College of Life Science,Northeast ForestryUniversity,Harbin 150040,China)
S636.9
A
1674-8646(2015)02-0004-04
2014-11-05
黑龙江省科技攻关重大项目(GA09B201-02);黑龙江省自然科学基金重点项目(ZD201105)
林晗婧(1989-),女,黑龙江哈尔滨人,在读硕士,主要从事植物生理生态学研究。
孙广玉(1963-),男,黑龙江巴彦人,教授,博士研究生导师,主要从事植物生理生态学研究,e-mail∶sungy@vip.Sina.com
