Thermodynamics of GPCR activation
2015-08-08XuejunZhangYeZhouCanCaoNationalLaboratoryofMacromoleculesNationalCenterofProteinScienceBeijingInstituteofBiophysicsChineseAcademyofSciencesBeijing000China
Xuejun C.Zhang,Ye Zhou,Can CaoNational Laboratory of Macromolecules,National Center of Protein Science-Beijing,Institute of Biophysics,Chinese Academy of Sciences,Beijing 000,China
OPINION
Thermodynamics of GPCR activation
Xuejun C.Zhang1✉,Ye Zhou1,Can Cao11National Laboratory of Macromolecules,National Center of Protein Science-Beijing,Institute of Biophysics,Chinese Academy of Sciences,Beijing 100101,China
Received:10 December 2015/Accepted:6 January 2016/Published online:3 February 2016
INTRODUCTION
G-protein coupled receptors(GPCRs)are the largest family of signaling proteins that are responsible for information input from the extracellular environment. The plasma membrane in which GPCRs reside usually carries an electrostatic membrane potential(ΔΨ).This potential and its variations in some cell types are important for cellular functions,including GPCR signaling(Mahaut-Smith et al.2008;Zhang et al.2014).
The phenomenon of membrane potential-sensitivity signaling has been observed in many GPCRs,including the M2receptor(Ben-Chaim et al.2003),P2Y1(Gurung et al.2008),the α2A-adrenoceptor(Rinne et al.2013), the β1-adrenoceptor(Birk et al.2015),the dopamine D2receptor(Sahlholm et al.2008),and the histamine H3 receptor(Sahlholm et al.2012).The signaling capacity of a GPCR changes when the membrane potential is experimentally modulated.A question often raised is what and where is the voltage sensor?Researchers would like to know which amino acid residue(s)in the GPCR molecule is responsible for ΔΨ-sensitivity.
In general,a particular ligand-GPCR pair can be considered as a unique system that has different thermodynamic parameters and pharmacological properties from other ligand-GPCR combinations(Masuho et al. 2015).In real in vivo situations,such systems are necessarily coupled with each other and with upstream and downstream networks,and their thermodynamic parameters can be infuenced strongly by the environment,including ΔΨ.In the following report,we will frst discuss the thermodynamics of an isolated GPCR activation process according to the classical view of ligandreceptor equilibrium,and then attempt to address the above questions about ΔΨ-sensitivity.
THERMODYNAMICS
A GPCR molecule contains seven transmembrane(TM) helices(Palczewski et al.2000).Activation of the GPCR is associated with a conformational rearrangement of the 7-TM domain.Here,opening of the cytosolic side of the 7-TM domain facilitates interactions with downstream effectors such as G-proteins(Rasmussen et al. 2011).Roughly speaking,a typical GPCR possesses two major states:the ground(R)and active(R*)states (Zhang et al.2014;Lamichhane et al.2015).Although multiple active states have been proposed to explain socalled biased-signaling phenomena(Onaran et al.2014), the two-state model remains the cornerstone of the GPCR activation study:the free-energy differences and energy barriers between the multiple active states are usually signifcantly smaller than those between the ground and active states.It is important to note that multiple active states are not sequential steps in the activation process.Instead,they are thermodynamically parallel to and equilibrate with each other.Therefore, the two-state model is a reasonable approximation of GPCR activation and multiple activation states may be considered as a perturbation to the two-state model.
In the following discussion on the thermodynamics of GPCR activation,we will follow the conventions used in two-state transporters(see Appendix 3 in Zhang et al. (2015)).In particular,a negative free-energy term indicates that the corresponding step is thermodynamically favored.In principle,for each of the two states a GPCR may or may not bind with an agonist.Thus,there are four sub-states that are in thermodynamic equilibrium with each other(Scheme 1).In addition,we assume that there is no cooperativity among GPCR molecules(i.e.,the Hill coeffcient is one).At a given agonist concentration(denoted as[S],where‘‘S’’stands for substrate or agonist),the ratio of the probability ofthe GPCR in the ground state to that in the active state is defned as a partition function,f([S]).This function can be estimated based on experimental measurements,for example,using the single molecule FRET technique (Lamichhane et al.2015).Mathematically,it can be proved that three independent thermodynamic parameters are necessary and suffcient to describe the partition function of the four sub-states system.

Scheme 1 Thermodynamic equilibrium of a four-state GPCR-ligand system
In the absence of a bound agonist,there is a freeenergy term ΔGEassociated with the transition from the ground to active state,where the subscript‘‘E’’stands for‘‘elastic’’conformational energy stored in the active state.In principle,ΔGEcan be determined experimentally by measuring the partition function in the absence of an agonist(i.e.,ΔGE≡-RTln(f(0))).Thus,ΔGEdetermines the basal activity of the GPCR.In the absence of an agonist,if ΔGE=0,the GPCR would have a 50/50 chance of remaining in both the ground and active state,whereas if ΔGE/RT≪0,the GPCR would remain in the ground state most of the time.
Binding of an agonist,by defnition,promotes activation(Fig.1).The binding affnity of a given agonist towards its target GPCR depends on which state the receptor is in.In general,for a given GPCR-agonist pair, the dissociation constant in the ground state(termed Kd0)is different from that in the active state(Kd1)(see Scheme 1).The experimentally determined,apparent dissociation constant,Kd,app,is a probability-weighted average value of the two states(Zhang et al.2015).The affnity difference between the two states is associated with another important free-energy term,called the differential binding energy ΔGD(≡RTln(Kd1/Kd0)).In the case of ΔGD<0,this free-energy term functions as part of the activation driving energy(Fig.1).Similar to ΔGE,ΔGDcan be determined experimentally by measuring the partition function both in the absence of and at saturating concentrations ofthe agonist (i.e., ΔGD=RTln(f(∞)/f(0)))(Zhang et al.2015).Assuming that the GPCR-ligand system is in a thermodynamic equilibrium,the following three scenarios are of particular interest to GPCR functioning.(i)If ΔGD<0,the active state becomes thermodynamically more favorable than in the absence of the ligand,and the ligand is thus an agonist or partial agonist.(ii)If ΔGD=0,ligand binding will not change the distribution of states,and the ligand would be an antagonist.(iii)If ΔGD>0, ligand binding will stabilize the ground state,and the ligand would function as an inverse agonist.The freeenergy term ΔGD-ΔGEis directly related to the intrinsic effcacy(ε)of classical receptor theory(Onaran et al. 2014).Regarding the above-mentioned three independent parameters,they can be chosen from[S]/Kd0,[S]/ Kd1,f(0),f(∞),ΔGD,or ΔGE,which are of clear physical meaning.

Fig.1 Schematics of the energy landscape of GPCR activation, driven solely by agonist binding.Solid and dashed horizontal lines depict real and imaginary states,respectively.State‘‘R’’and‘‘R*’’stand for the ground and active states,respectively,and‘‘S’’stands for substrate(agonist).Tilted arrows depict transitions between neighboring states.Red arrows are associated with the chemical potential of agonist binding,where the sum of the chemical potential is zero to follow the frst and second laws of thermodynamics.Here,we assume that the system is in thermodynamic equilibrium;i.e.,the system does not consume energy from the environment.Therefore,the start and end points are identical
ADDITIONAL DRIVING ENERGY
The binding energy of agonists may vary signifcantly between receptors and between different agonists for the same receptor.In addition,there are usually energy barrier(s)(e.g.,ΔGf‡in Fig.2)between the ground and active states,which affect the kinetics of receptor activation.In the case that ΔGf‡is signifcantly larger thanthe amplitude of the substrate-loading energy,e.g., ΔGL(S)(=-RTln([S]/Kd0),where the subscript‘‘L’’stands for loading of the agonist),some other forms of driving energy must be provided for GPCR activation (Zhang et al.2013).In particular,in our recent hypothesis on the proton transfer-mediated activation mechanism of class-A GPCRs(Zhang et al.2014),an agonist binding-triggered proton release is proposed to drive the transition from the ground state to the active state(see the green line-marked transition in Fig.2).
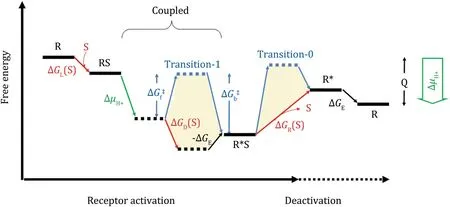
Fig.2 Schematics of the energy landscape of GPCR activation in the presence of proton transfer.Here,we assume that the free-energy associated with proton release(ΔμH+)only affects the kinetics of the activation process by overcoming the(forward)transition-state energy barrier,ΔGf‡.The starting and ending states are structurally and chemically identical,except that they differ by released heat,Q, that is converted from ΔμH+in one cycle of the GPCR activation.There are two transition-state energy barriers(termed transition-0 and -1)separating the ground and active states.The forward and backward energy barriers of transition-1 are labeled as ΔGf‡and ΔGb‡, respectively
MEMBRANE POTENTIAL
As a self-contained capacitor,the cellular membrane carries an electrostatic potential.Not only is this potential an important source of energy for the cell,it also affects the equilibrium conformations of all membrane proteins that carry electric charges.GPCRs are no exception.The conformation of the GPCR in a given state is the result of a balance between the ΔΨ-associated electrostatic force and the hydrophobic mismatch force from the lipid bilayer,as well as ligand binding(Zhang et al.2013).Such a balance can be analogous to,because of gravity,the weights and distribution of cargos in a boat,which affect the equilibrium position and the kinetic properties of the boat.Such an analogy may aid our understanding of the biased signaling of GPCR,in which different agonists may result in distinct spectrums of activation of downstream effectors(Onaran et al.2014;Masuho et al.2015).Moreover,the properties of a GPCR(e.g.,a set of parameters such as [S]/Kd0,[S]/Kd1,and-ΔGE)are ΔΨ dependent.For example,the sign of the electric charge(s)of the GPCR-agonist complex determines the direction of the overall movement of the complex in response to a change in ΔΨ (i.e.,ΔΔΨ).In particular,if it carries a positive charge, the complex will shift towards the extracellular side of the membrane upon de-polarization of the membrane potential(e.g.,from-90 mV to+60 mV).More detailed consequences of such a movement depend on the equilibrium conformations of the ground and active states at the new ΔΨ.In short,a voltage sensor may not be a localized region or the side chain of a particular amino acid of the GPCR molecule,rather the overall charge distribution of the protein may function as the voltage sensor.
POTENCY VERSUS THE EFFICACY EFFECT OF ΔΨ-SENSITIVITY
The membrane potential may affect the activation process of a GPCR in many ways.Experimentally,it has been shown that ΔΨ de-polarization may either deactivate or potentiate some GPCRs,and the effects may affect either potency(binding ability)or effcacy of the agonist.According to Le Chaˆtelier’s principle,a hydrophobic mismatch induced upon movement of a GPCR towards the cytosol(cytosolic movement)favors the opening of the cytosol side of the GPCR molecule, thus minimizing the exposed hydrophobic TM helices tothe cytosol.This is similar to what has been proposed for the major-facilitator superfamily transporters (Zhang et al.2015).Therefore,if an electrostatic interaction between the charge and ΔΔΨ results in a cytosolic movement,the GPCR is more likely to become active.In contrast,if the movement is towards the extracellular direction,deactivation of the GPCR will occur.In the latter case,if the energy gained from the charge movement is smaller than the transition-state energy barrier,its effect is likely to be local and minor, and only effects on effcacy(i.e.,activation of downstream effectors such as G-proteins or arrestins)may be observed.However,if the energy gain is large enough to overcome the transition-state energy barrier,the effect will be more global,and ligand potency may be affected.
In the case of a potency effect,de-polarization may reduce the affnity of the GPCR towards the agonist(Ben Chaim et al.2013).The corresponding deactivation process is often slower,consistent with a transition back to the ground state.Upon re-polarization,the activation curve appears to recover slowly.Saturated concentrations of the agonist may diminish the potency effect.A potency effect is exemplifed for the α2A-adrenoceptor binding with norepinephrine(Fig.3,left panel)(Rinne et al.2013).In contrast,in the case of the effcacy effect, de-polarization may decrease the binding of downstream effectors by directly adjusting the active GPCR to a sub-optimal conformation.Because of the nature of the electrostatic interaction,the effcacy effect is usually fast.Re-polarization often reverts the GPCR to the original active state quickly.An effcacy effect is exemplifed in the β1-adrenoceptor(Fig.3,right panel)(Birk et al.2015).

Fig.3 Typical potency and effcacy effects of ΔΔΨ,based on real data from(Rinne et al.2013)and(Birk et al.2015).Note that the left panel may be the result of both potency and effcacy effects, because the vertical axis represents the activation of downstream G-proteins,rather than a direct effect on agonist binding
In a real GPCR,both potency and effcacy effects of ΔΔΨ may function in combination,although the large potency effect is likely to overshadow the small effcacy effect.In addition,ΔΔΨ may also affect(even abolish) the proton transfer-mediated activation mechanism (Zhang et al.2014),thus changing the kinetics of GPCR activation.However,in some cases,a GPCR may carry no electric charges in the active state,and thus ΔΔΨ may not have a signifcant effect on such GPCR activation, which is most likely the situation for β2-AR(Birk et al. 2015).Taken together,thermodynamic discussion about GPCR activation places GPCR-mediated signal transduction on a more physically meaningful ground,and covers many interesting observations about GPCR activation under a unifed theoretical framework.
Acknowledgments This work was supported by‘‘973’’Program (2014CB910104),Chinese Academy of Sciences(XDB08020301), and National Natural Science Foundation of China(31470745).
Compliance with Ethical Standards
Confict of Interest Xuejun C.Zhang,Ye Zhou,and Can Cao declare that they have no confict of interest.
Human and Animal Rights and Informed Consent This article does not contain any studies with human or animal subjects performed by any of the authors.
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License(http:// creativecommons.org/licenses/by/4.0/),which permitsunrestricted use,distribution,and reproduction in any medium,provided you give appropriate credit to the original author(s)and the source,provide a link to the Creative Commons license,and indicate if changes were made.
Ben Chaim Y,Bochnik S,Parnas I,Parnas H(2013)Voltage affects the dissociation rate constant of the m2 muscarinic receptor. PLoS One 8:e74354
Ben-Chaim Y,Tour O,Dascal N,Parnas I,Parnas H(2003)The M2 muscarinic G-protein-coupled receptor is voltage-sensitive. J Biol Chem 278:22482-22491
Birk A,Rinne A,Bunemann M(2015)Membrane potential controls the effcacy of catecholamine-induced beta1-adrenoceptor activity.J Biol Chem 290:27311-27320
Gurung IS,Martinez-Pinna J,Mahaut-Smith MP(2008)Novel consequences of voltage-dependence to G-protein-coupled P2Y1 receptors.Br J Pharmacol 154:882-889
Lamichhane R,Liu JJ,Pljevaljcic G,White KL,van der Schans E, Katritch V,Stevens RC,Wuthrich K,Millar DP(2015)Singlemolecule view of basal activity and activation mechanisms of the G protein-coupled receptor beta2AR.Proc Natl Acad Sci USA 112:14254-14259
Mahaut-Smith MP,Martinez-Pinna J,Gurung IS(2008)A role for membrane potential in regulating GPCRs.Trends Pharmacol Sci 29:421-429
Masuho I,Ostrovskaya O,Kramer GM,Jones CD,Xie K,Martemyanov KA(2015)Distinct profles of functional discrimination among G proteins determine the actions of G proteincoupled receptors.Sci Signal 8:ra123
Onaran,Rajagopal S,Costa T(2014)What is biased effcacy? Defning the relationship between intrinsic effcacy and free energy coupling.Trends Pharmacol Sci 35:639
Palczewski K,Kumasaka T,Hori T,Behnke CA,Motoshima H,Fox BA,Le Trong I,Teller DC,Okada T,Stenkamp RE et al(2000)Crystal structure of rhodopsin:A G protein-coupled receptor. Science 289:739-745
Rasmussen SG,DeVree BT,Zou Y,Kruse AC,Chung KY,Kobilka TS, Thian FS,Chae PS,Pardon E,Calinski D et al(2011)Crystal structure of the beta2 adrenergic receptor-Gs protein complex.Nature 477:549-555
Rinne A,Birk A,Bunemann M(2013)Voltage regulates adrenergic receptor function.Proc Natl Acad Sci USA 110:1536-1541
Sahlholm K,Nilsson J,Marcellino D,Fuxe K,Arhem P(2008) Voltage-dependence of the human dopamine D2 receptor. Synapse 62:476-480
Sahlholm K,Nilsson J,Marcellino D,Fuxe K,Arhem P(2012) Voltage sensitivities and deactivation kinetics of histamine H(3) and H(4) receptors. Biochim Biophys Acta 1818:3081-3089
Zhang XC,Sun K,Zhang L,Li X,Cao C(2013)GPCR activation: protonation and membrane potential.Protein Cell 4:747-760
Zhang XC,Cao C,Zhou Y,Zhao Y(2014a)Proton transfer-mediated GPCR activation.Protein Cell 6:12-17
Zhang XC,Cao C,Zhou Y,Zhao Y(2014b)Proton transfer-mediated GPCR activation.Protein Cell 6:12-17
Zhang XC,Zhao Y,Heng J,Jiang D(2015)Energy coupling mechanisms of MFS transporters.Protein Sci 24:1560-1579
✉Correspondence:zhangc@ibp.ac.cn(X.C.Zhang),
杂志排行

Biophysics Reports的其它文章
- Implications for directionality of nanoscale forces in bacterial attachment
- Visualizing the Ensemble Structures of Protein Complexes Using Chemical Cross-Linking Coupled with Mass Spectrometry
- E2-2,a novel immunohistochemical marker for both human and monkey plasmacytoid dendritic cells
- Radiolig and saturation binding for quantitative analysis of ligand-receptor interactions
- Pseudomonas sp.LZ-Q continuously degrades phenanthrene under hypersaline and hyperalkaline condition in a membrane bioreactor system