哮喘相关的miR-201-3p靶基因的预测及生物信息学分析
2015-08-07周萍萍吴露依黄丹丹靳文杰马智达姜玉新
周萍萍,尹 康,吴露依,黄丹丹,靳文杰,马智达,姜玉新
(皖南医学院 1.生理学教研室;2.临床医学院,安徽 芜湖 241002)
支气管哮喘(简称哮喘)是以持续气道炎症、气道高反应性和气道重塑为主要特征的慢性呼吸道疾病,症状主要表现为反复发作性喘息、胸闷、咳嗽,其发病率高达29.1% ~32.2%[1]。哮喘发病受遗传及环境双重因素影响,发病机制主要是CD4+T细胞亚群Th1/Th2失衡,特别是偏向于Th2分化所致,其他CD4+T细胞亚群Th17及调节性T细胞(Treg)也参与了该病发生[3]。其病理特征为气道嗜酸粒细胞、T细胞、肥大细胞、中性粒细胞等长期浸润,异常的Th2细胞应答及其所分泌细胞因子产生的级联效应,最终导致气道慢性炎症。Th2细胞分泌的IL-4、IL-5、IL-9和IL-13与嗜酸粒细胞浸润、气道高反应性、高水平 IgE等密切相关[2-3]。哮喘气道的慢性炎症及重塑最终可导致不可逆性气流受限及对激素治疗的无反应[4]。
miRNA是一类长约为19~25个核苷酸的小分子RNA。主要通过结合靶基因mRNA的3'端非翻译区(3'-untranslated region,UTR),参与对靶基因的转录后调节[5]。近年来发现,某些miRNAs如miR-126、miR-145、let-7等可以调节哮喘气道炎症,分别表现出促炎、抗炎或对炎症的双重作用。通过调控这些miRNAs的表达以减轻哮喘的气道炎症,或将成为哮喘治疗的新方法及研究新热点。前期实验中,本课题组按文献[6]报道的方法成功构建了过敏性哮喘小鼠模型,制备了哮喘模型小鼠的单个脾细胞后,用免疫磁珠分选了其CD4+T细胞,提取其总RNA并用microRNA芯片检测了miRNA表达谱。与对照组相比,哮喘模型小鼠的CD4+T细胞中共有12种miRNAs表达升高,如miR-201-3p等(结果尚未发表)。该结果提示,这些miRNAs可能参与了哮喘的发病过程,尤其是对CD4+T细胞的调控作用。本研究拟通过生物信息学方法预测miR-201-3p的潜在靶基因,并对这些靶基因进行Gene Ontology(GO)基因分类以及KEGG信号通路富集,以期探究miR-201-3p在哮喘发病过程中对CD4+T细胞可能的调控作用。
1 材料与方法
1.1 靶基因预测 分别用miRanda(http://www.microrna.org/)、miRDB(http://www.mirdb.org/)、miRWalk(http://www.umm.uni-heidelberg.de/)和Targetscan(http://www.targetscan.org/)4 个数据库在线预测miR-201-3p的靶基因,并对获得的靶基因在4个数据库中的分布情况构建Venn图。
1.2 靶基因的GO分类 对1.1中获得的靶基因,剔除仅出现于2个或2个以下数据库的靶基因,剩下的靶基因用于后续分析。运用软件Cytoscape(2.8.1 版)中的功能插件 BiNGO(2.44 版)进行 GO分类并绘图。选择物种“Mus musculus”,以Benjamini&Hochberg's FDR correction进行多重检验,P<0.01为显著性阈值。
1.3 靶基因的KEGG信号通路分析 利用DAVID v6.7 网站(http://david.abcc.ncifcrf.gov/)中的KEGG(Kyoto encyclopedia of Genes and Genomes)数据库,以“Mus musculus”为研究背景,P <0.05为显著性阈值,对用于GO分类的靶基因进行信号通路富集分析。
2 结果
2.1 预测靶基因 在 miRanda、miRDB、miRWalk和Targetscan 4个数据库中共预测获得5201个潜在靶基因,这些靶基因在这4个数据库的分布如图1所示。由图1可知,在3个及3个以上数据库同时出现的靶点共计1082个靶基因。

图1 miR-201-3p靶基因在 miRanda、miRDB、miRWalk和Targetscan数据库中的分布Fig 1 The distribution of target genes of miR-201-3p in miRanda,miRDB,miRWalk and Targetscan databases
2.2 Cytoscape的GO 分析
2.2.1 生物学进程分析 对获得的miR-201-3p的1082个靶基因进行GO分类,结果表明:共有963个靶基因涉及245条生物学进程(图2,P<0.01),其中生物学进程中包含的靶基因数量排在前4位的分别为细胞进程(cellular process,62.5%)、生物学调节(biological regulation,45.8%)、生物学进程的调节(regulation of biological process,43.7%)和细胞进程的调节(regulation of cellular process,41.1%)。其他的生物学进程见表1(仅列出前20项)。
2.2.2 分子功能分析 对1082个靶基因进行分子功能分析的结果表明,这些靶基因的功能包括结合(binding)、蛋白结合(protein binding)、催化活性(catalytic activity)等,这些分子功能的关联关系见图3。
2.3 KEGG信号通路 对1082个靶基因进行KEGG信号通路富集的结果表明:这些靶基因隶属于30条信号通路(表2),其中与炎症相关的信号通路主要包括 MAPK、Wnt、趋化因子、TGF-β、T 细胞受体、胰岛素信号、VEGF信号通路等(P<0.01)。其中,有27个靶基因出现于 MAPK信号通路中,如MAP3K7、TRAF2、MAPKAPK2、GNA12 和 MAX 等(图4);16个靶基因出现于Wnt信号通路中,如APC2、PPP2R5D、WNT3A 和 F2D5等(图5);而T 细胞受体信号通路中富集了 PDK1、TRP53和RAF1等12个基因(图6)。

表1 miR-201-3p靶基因的生物学进程结果Tab 1 Biological process of predicted target genes of miR-201-3p
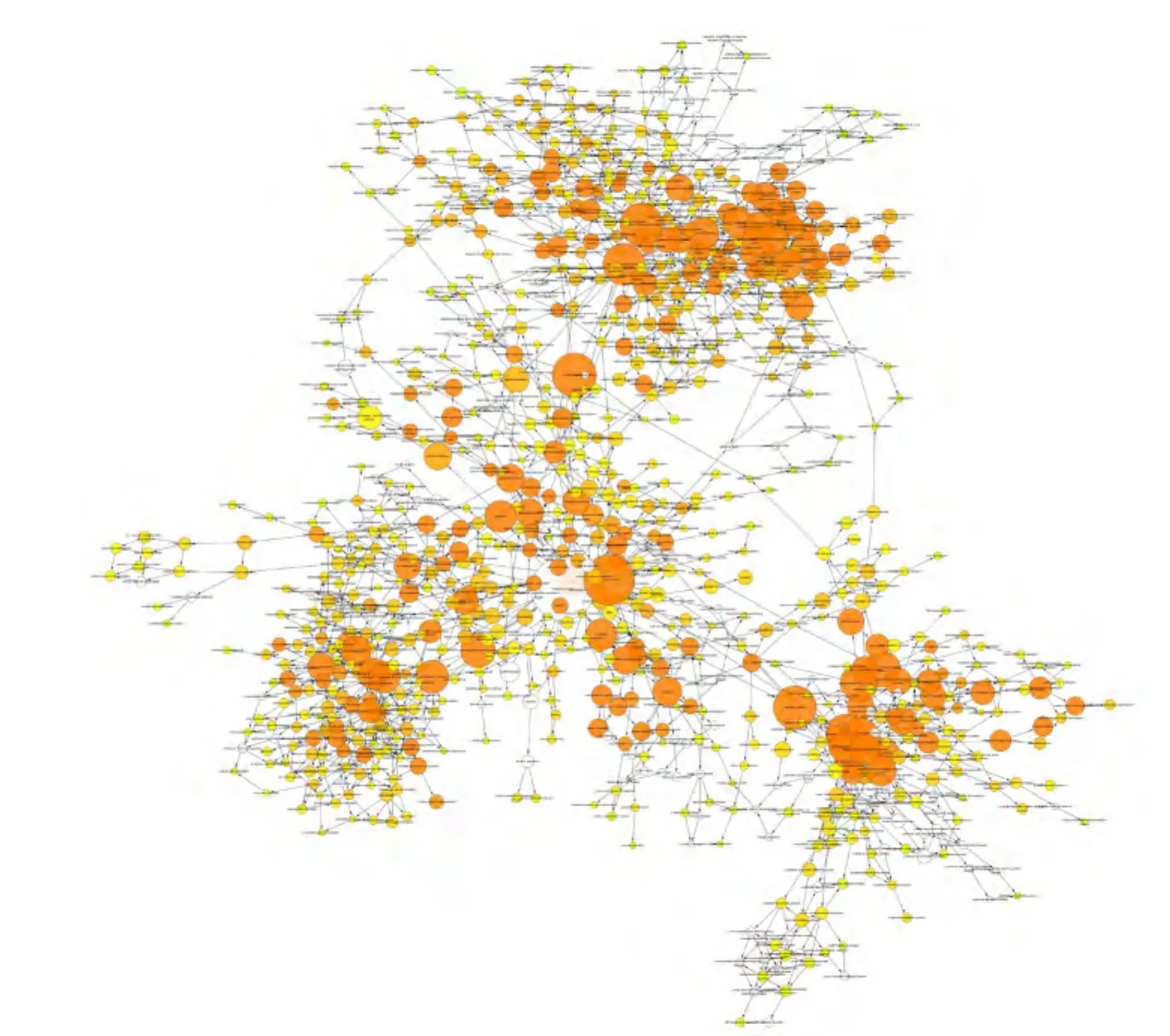
图2 miR-201-3p靶基因的生物学进程图Fig 2 Biological process of the predicted target genes of miR-201-3p

图3 miR-201-3p靶基因的分子功能图Fig 3 Molecular function of the predicted target genes of miR-201-3p
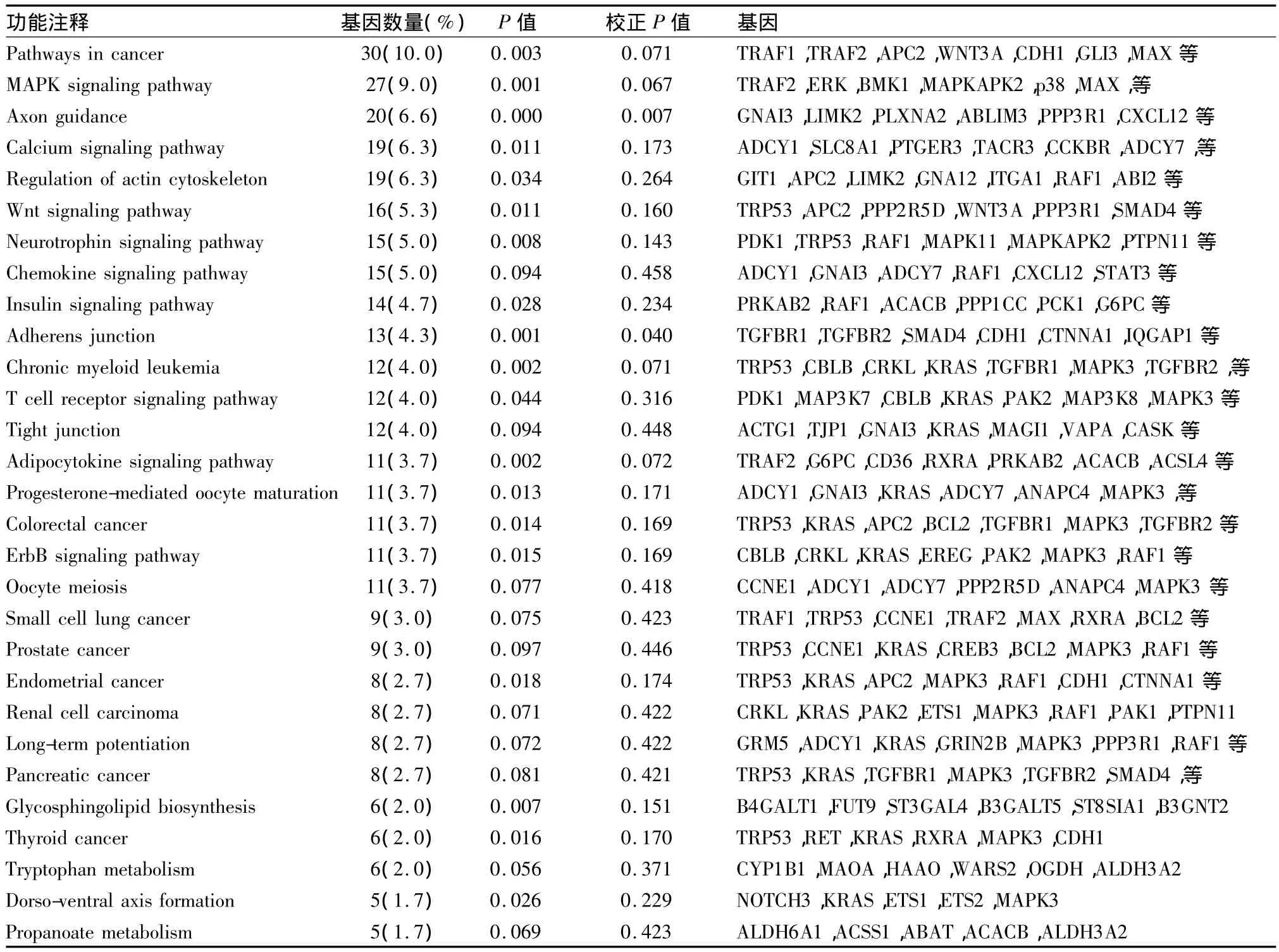
表2 miR-201-3p预测靶基因的KEGG信号通路富集分析结果Tab 2 Pathway enrichment of the predicted target genes of miR-201-3p
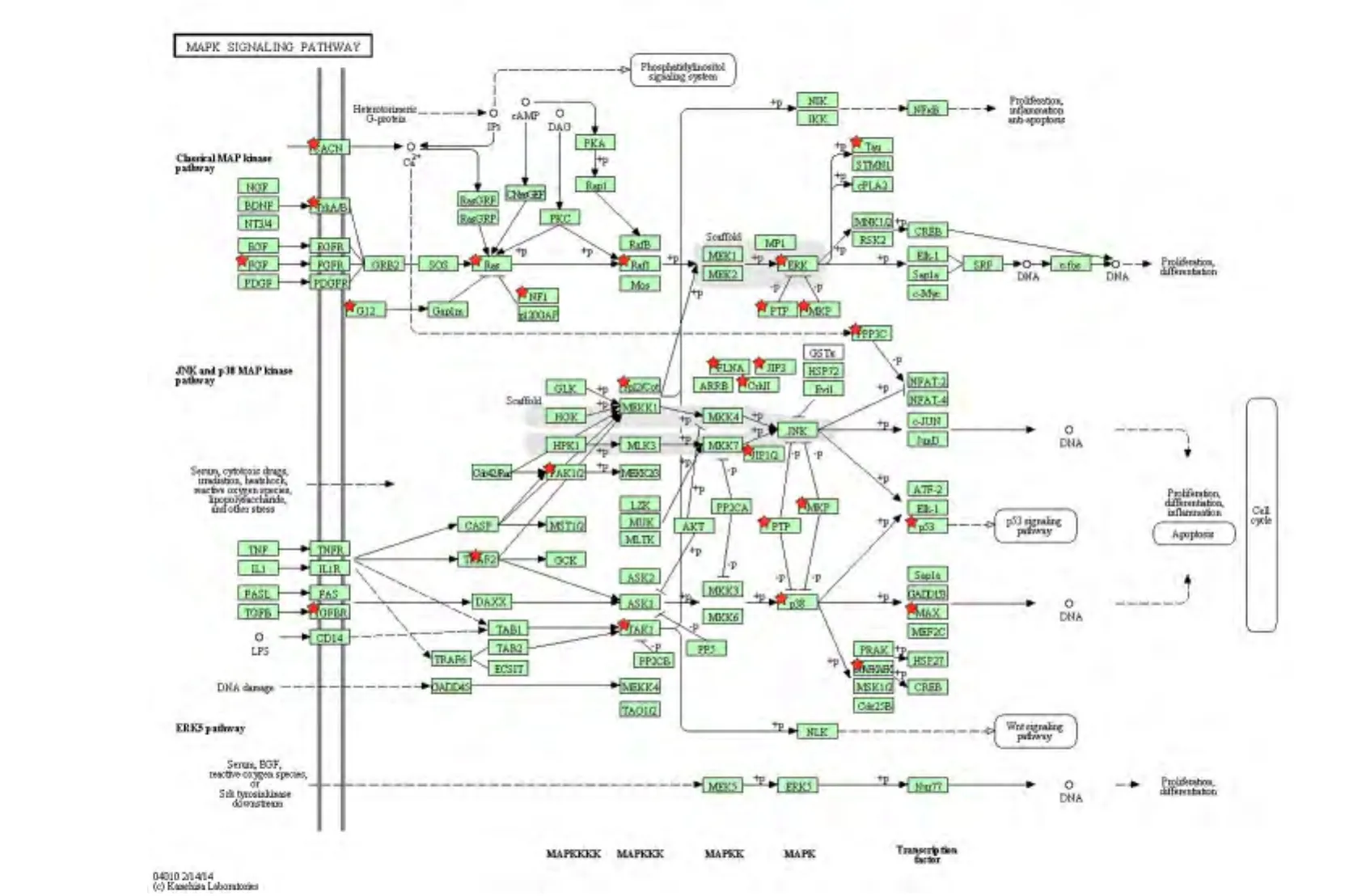
图4 miR-201-3p靶基因在MAPK信号通路图中的分布Fig 4 Distribution of the predicted target genes of miR-201-3p in MAPK signaling pathway
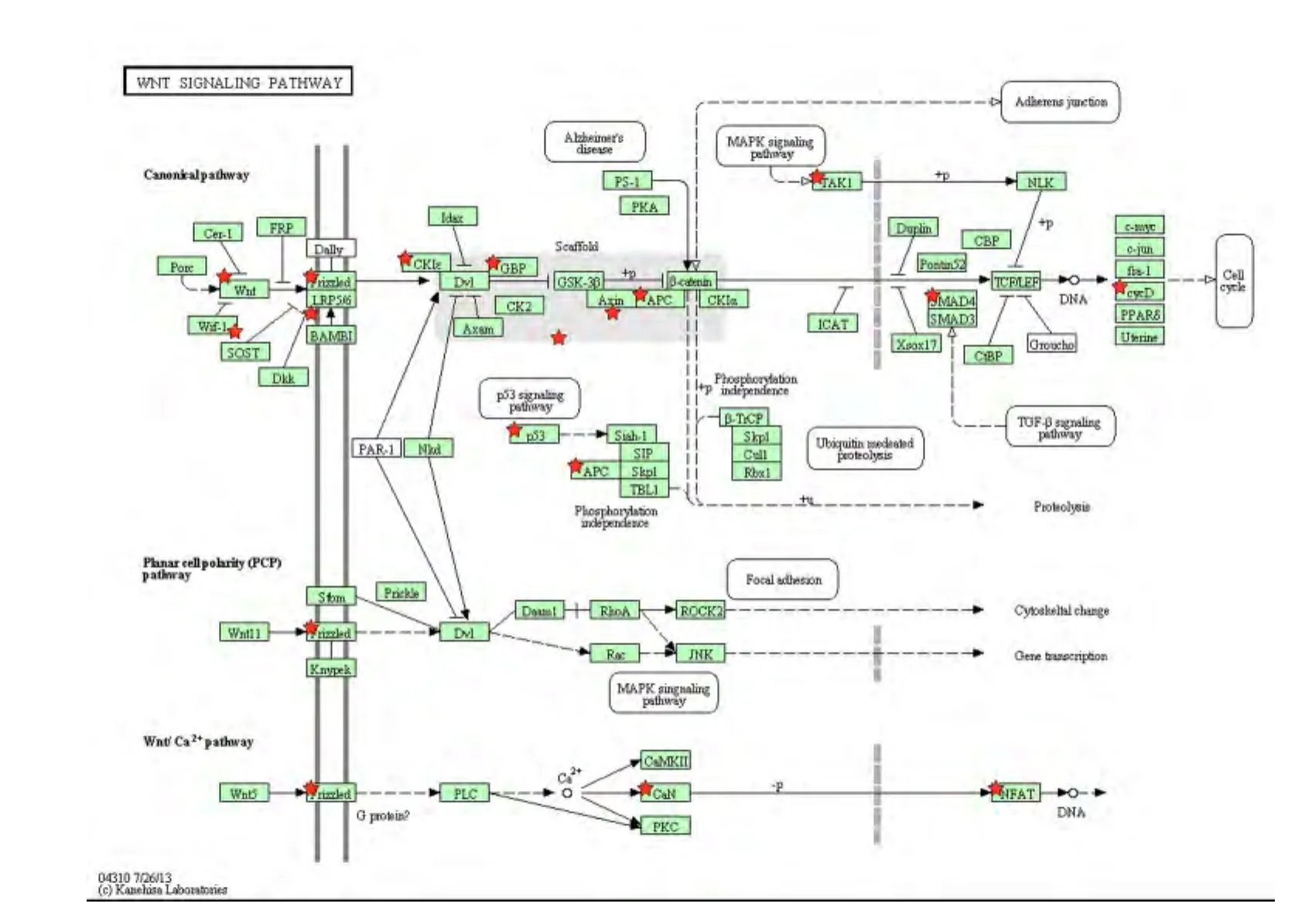
图5 miR-201-3p靶基因在WNT信号通路图中的分布Fig 5 Distribution of the predicted target genes of miR-201-3p in WNT signaling pathway
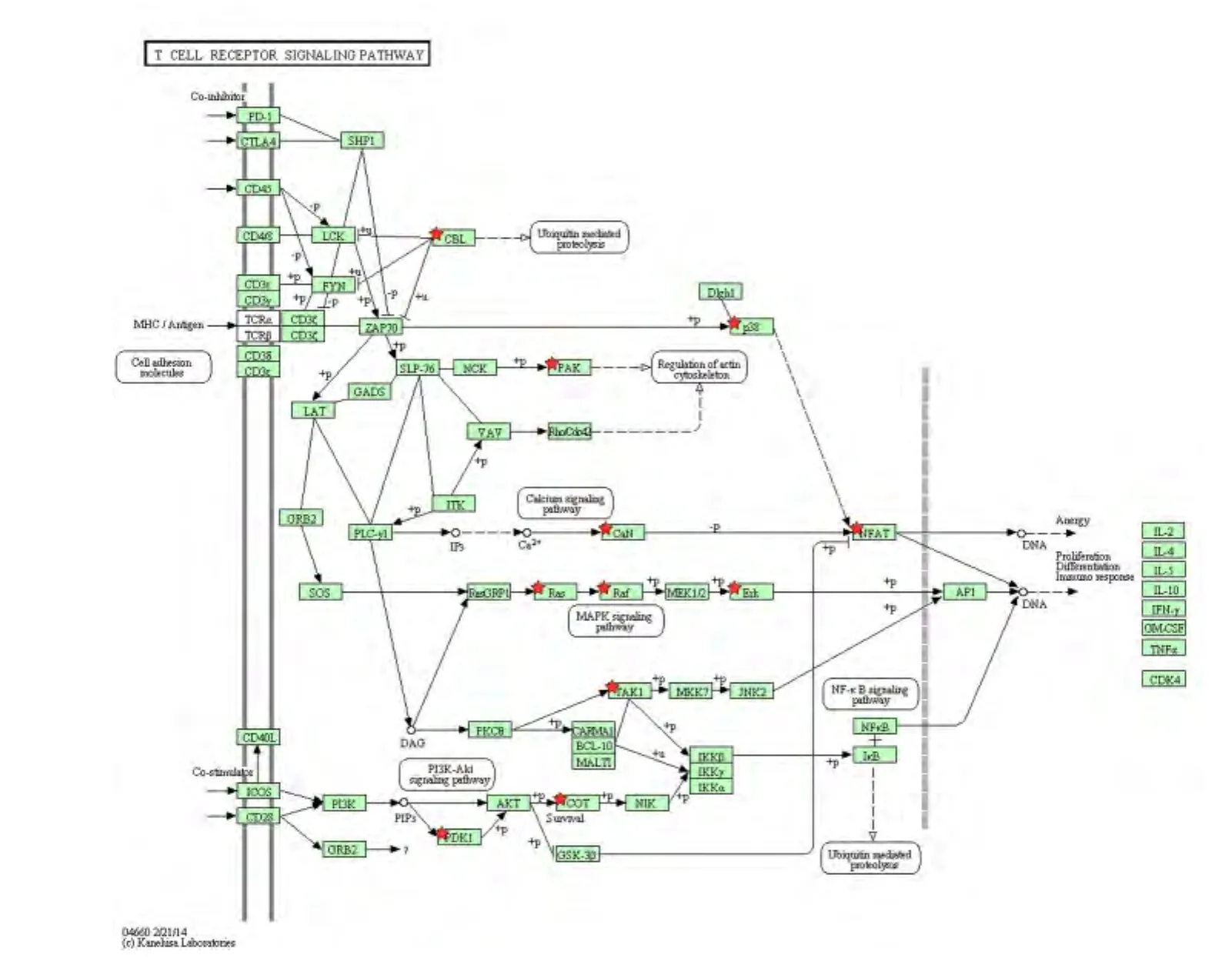
图6 miR-201-3p靶基因在T细胞受体信号通路图中的分布Fig 6 Distribution of the predicted target genes of miR-201-3p in T cell receptor signaling pathway
3 讨论
哮喘为临床常见疾病,其发病率逐年升高[1]。miRNA在哮喘的发生发展中发挥重要的作用。Moschos等[7]发现,细菌脂多糖(LPS)诱导了炎性肺组织中miRNA表达谱的改变。miRNAs在CD4+T细胞分化发育过程中起着重要的调控作用,如miR-155可抑制转录因子c-Maf的表达,下调miR-155可促进IL-4、IL-5和IL-13等Th2型细胞因子分泌,同时降低了 B 和 T 细胞反应[8]。Zhang等[9]研究表明过敏性哮喘患者外周血CD4+T细胞中miR-155表达下调,其表达水平与哮喘严重程度相关,提示miRNA调控CD4+T细胞分化在哮喘中发挥重要作用。前期实验发现,miR-201-3p在哮喘模型的小鼠脾CD4+T细胞中表达升高,提示该miRNA可能参与了哮喘发病过程中CD4+T细胞功能的调节。
miRNA通过对靶基因进行转录后的表达调控,在多种生理、病理过程中发挥重要作用。Tomankova等[10]研究表明,miRNA在哮喘、肺癌、肺纤维化等肺部疾病中表达异常,其可通过多条信号转导通路参与气道重塑等病理生理过程。本研究应用miRanda、miRDB、miRWalk 和 Targetscan 4 个数据库对miR-201-3p的潜在靶基因预测后,共获得了5201个潜在靶基因,其中有1082个靶基因同时出现在3个和3个以上数据库。进一步对这1082个靶基因进行GO分类后,发现这些靶基因的分子功能主要是蛋白结合、催化活性等。KEGG信号通路富集结果发现,这些靶基因主要存在于与炎症有关的信号通路如 MAPK、Wnt、趋化因子、TGF-β、胰岛素信号、T细胞受体信号通路(T cell receptor signaling pathway)、VEGF信号通路等。
本研究获得的T细胞受体信号通路基因包括PDK1、MAP3K7、CBLB、KRAS、PAK2 等。Park 等[11]将T细胞的PDK1基因敲除后发现TCR-CD28信号无法诱导活化NF-kB或蛋白激酶C-θ的磷酸化,说明PDK1对整合TCR和CD28之间的信号至关重要。KRAS原癌基因是细胞增殖的重要调节因子[12],研究表明,let-7(一种 miRNA)可通过结合KRAS的3'UTR进而抑制KRAS的表达,从而抑制非小细胞肺癌的发生[13]。miR-201-3p的靶基因PDK1和KRAS在哮喘发病过程中是否通过该信号通路发挥作用有待于进一步研究。
经典的MAPK通路(MAP kinase pathway)主要包括细胞外信号调节激酶(ERK)信号通路、c-Jun N端激酶(JNK)/应激活化蛋白(SAPK)信号通路、p38 MAPK信号通路和ERK5/大丝裂素活化蛋白激酶(BMK1)信号通路,参与了细胞运动、凋亡、分化及生长增殖等多种生理过程。研究证实p38 MAPK和ERK是烟草烟雾诱发气道炎症和肺气肿的重要信号通路[14]。Shin等研究发现褪黑素可减弱ERK通路磷酸化从而有效抑制由LPS或吸烟导致的气道炎症[15]。Wan等证实分泌球蛋白(SCGB32A)可限制ERK和JNK通路的磷酸化,从而减轻由LPS所致的气道炎症[16]。这些MARK信号通路既高度特异又存在一定的交叉。p38和 BMK1在 TNF-α诱导 cjun转录的调控中具有协同作用。p38和BMK1可分别上调MEF2A和MEF2D的转录活性。TNF-α通过激活p38和BMK1,使MEF2A和MEF2D磷酸化,从而诱导MEF2A/MEF2D异源二聚体形成,这在TNF-α诱导的c-jun基因转录表达调控中起着重要的作用[17]。本研究发现,TRAF2、ERK、BMK1、MAPKAPK2、MAP3K、MAX等27个分子富集于MAPK通路(图4),这说明miR-201-3p可能通过该通路参与了哮喘发病过程中对CD4+T细胞的调节作用。
正常生理状态下,干细胞中β-catenin主要通过启动经典的 Wnt/β-catenin通路激活下游基因[18];而在低氧时,低氧诱导因子1α(HIF-1α)也促进βcatenin活性以调节 Wnt/β-catenin信号通路[19]。应用DNA微阵列方法发现在支气管哮喘患者外周血中的Wnt 5a显著升高[20]。本研究也发现有16个靶基因富集于 Wnt/β-catenin信号通路(图5),提示miR-201-3p可能通过该信号通路诱发哮喘的发生。
总之,本研究预测了miR-201-3p的靶基因及可能涉及的信号通路,这些信号通路在以往研究中已被证实参与了气道炎症与气道重塑等病理生理过程。但miR-201-3p是否在炎症和气道重塑中也通过这些通路进行调节尚需进一步的实验验证。
[1]Reddel HK,Hurd SS,Fitzgerald JM.World Asthma Day.GINA 2014:a global asthma strategy for a global problem[J].Int J Tuberc Lung Dis,2014,18(5):505 -506.
[2]Robinson D,Hamid Q,Bentley A,et al.Activation of CD4+T cells,increased TH2-type cytokine mRNA expression,and eosinophil recruitment in bronchoalveolar lavage after allergen inhalation challenge in patients with atopic asthma[J].J Allergy Clin Immunol,1993,92(2):313 - 324.
[3]Wills-Karp M.Immunologic basis of antigen-induced airway hyperresponsiveness[J].Annu Rev Immunol,1999,17:255 -281.
[4]Seguin RM,Ferrari N.Emerging oligonucleotide therapies for asthma and chronic obstructive pulmonary disease[J].Expert Opin Investig Drugs,2009,18(10):1505 -1517.
[5]Li DF,Tian J,Guo X,et al.Induction of microRNA-24 by HIF-1 protects against ischemic injury in rat cardiomyocytes[J].Physiol Res,2012,61(6):555 -565.
[6]姜玉新,马玉成,李朝品.尘螨Ⅱ类改组变应原对哮喘小鼠免疫治疗的效果[J].山东大学学报:医学版,2012,50(10):50-55.
[7]Moschos SA,Williams AE,Perry MM,et al.Expression profiling in vivo demonstrates rapid changes in lung microRNA levels following lipopolysaccharide-induced inflammation but not in the anti-inflammatory action of glucocorticoids[J].BMC Genomics,2007,8:240.
[8]Rodriguez A,Vigorito E,Clare S,et al.Requirement of bic/microRNA-155 for normal immune function[J].Science,2007,316(5824):608-611.
[9]张莺莺,钟民,张梦莹,等.过敏性哮喘患者外周血CD4+T细胞中miR-155的表达及临床意义[J].细胞与分子免疫学杂志,2012,28(5):540 -543.
[10]Tomankova T,Petrek M,Kriegova E.Involvement of microRNAs in physiological and pathological processes in the lung[J].Respir Res,2010,11:159.
[11]Park SG,Schulze-Luehrman J,Hayden MS,et al.The kinase PDK1 integrates T cell antigen receptor and CD28 coreceptor signaling to induce NF-kappaB and activate T cells[J].Nat Immunol,2009,10(2):158-166.
[12]Karnoub AE,Weinberg RA.Ras oncogenes:split personalities[J].Nat Rev Mol Cell Biol,2008,9(7):517 - 531.
[13]Chin LJ,Ratner E,Leng S,et al.A SNP in a let-7 microRNA complementary site in the KRAS 3'untranslated region increases nonsmall cell lung cancer risk[J].Cancer Res,2008,68(20):8535-8540.
[14]Gu W,Song L,Li XM,et al.Mesenchymal stem cells alleviate airway inflammation and emphysema in COPD through down-regulation of cyclooxygenase-2 via p38 and ERK MAPK pathways[J].Sci Rep,2015,5:8733.
[15]Shin IS,Shin NR,Park JW,et al.Melatonin attenuates neutrophil inflammation and mucus secretion in cigarette smoke-induced chronic obstructive pulmonary diseases via the suppression of Erk-Sp1 signaling[J].J Pineal Res,2015,58(1):50 -60.
[16]Wang X,Tanino Y,Sato S,et al.Secretoglobin 3A2 Attenuates Lipopolysaccharide-Induced Inflammation Through Inhibition of ERK and JNK Pathways in Bronchial Epithelial Cells[J].Inflammation,2015,38(2):828 -834.
[17]赵明,刘亚伟,刘爱华,等.细胞信号转导分子在TNF-α诱导cjun基因表达中的作用[J].生物化学与生物物理学报,2000,32(3):258-264.
[18]Voronkov A,Krauss S.Wnt/beta-catenin signaling and small molecule inhibitors[J].Curr Pharm Des,2013,19(4):634 -664.
[19]Mazumdar J,O'brien WT,Johnson RS,et al.O2regulates stem cells through Wnt/beta-catenin signalling[J].Nat Cell Biol,2010,12(10):1007-1013.
[20]Van Scoyk M,Randall J,Sergew A,et al.Wnt signaling pathway and lung disease[J].Transl Res,2008,151(4):175 -180.
