云杉矮槲寄生内寄生系统的解剖学研究
2015-07-05朱宁波李学武才让旦周李积闰田呈明
朱宁波,陈 磊,白 云,李学武,才让旦周,李 涛,李积闰,田呈明*
(1 北京林业大学 林学院,北京100083;2 青海省林业厅森林病虫害防治检疫站,西宁810000;3 青海省门源县仙米林场,青海门源810300)
云杉矮槲寄生(Arceuthobium sichuanense)是 中国特有的一种多年生寄生性种子植物,属于槲寄生科(Viscaceae),油杉寄生属(Arceuthobium),寄生于青海云杉(Picea crassifolia)、紫果云杉(P.purpurea)、川 西 云 杉(P.likiangensis var.balfouriana)、西藏云杉(P.spinulosa)和青杄(P.wilsonii)等云杉属植物[1-3]。被寄生后,寄主植物发生生理和形态的改变[3-4],受侵染的寄主枝干部位抽发出大量下垂的扫帚状丛枝,树体再生能力和材质下降,严重时导致寄主死亡。近年来,仅在青海省云杉矮槲寄生害的发生面积近1万公顷,发病严重区域受害株率高达90%以上,严重威胁了当地的森林植被健康和生态环境安全[5-6]。
油杉寄生属的寄生植物通过其内寄生系统与寄主建立牢固的寄生关系,并吸取水分和营养物质[2]。其种子外具有一层粘性的半透明状的槲寄生素膜,用于种子粘附在寄主枝条上[2]。种子萌发过程中产生胚根,胚根接触到寄主枝条表面时停止伸长,开始形成固着器,随后在固着器的下方产生侵染钉,开始侵入寄主[7]。当成功侵入寄主组织后,矮槲寄生在寄主体内建立由皮层根和吸根构成的内寄生系统,进而与寄主建立牢固的寄生关系[8]。随后,矮槲寄生的侵染又可以分为局部侵染和系统侵染[8]。局部侵染时,矮槲寄生的内寄生系统和寄生芽均局限在侵染点附近的膨大区域[2,9]。进一步研究发现,所有的矮槲寄生在合适的寄主上均可以发生局部侵染[8]。而系统侵染时,内寄生系统可以向顶部扩展至寄主枝条内仅有初生生长的部分,甚至可以扩展至顶芽内,并随着寄主枝条的伸长而同步生长[8,10],但通常不会引起寄主枝条膨大[2]。目前,仅有A.douglasii经过内寄生系统的观察而证实可以进行系统侵染[10]。在其余种类中,根据所引起的症状,推测A.americanum、A.guatemalense、A.minutissimum、A.pusillum 和冷杉寄生(A.tibetense)可以引起系统侵染[2]。前期研究表明,云杉矮槲寄生可以引起寄主枝条膨大,形成扫帚状丛枝,同时也发现部分遭受云杉矮槲寄生侵染的枝条未发生膨大[11],因此,云杉矮槲寄生是否可以系统侵染仍需进一步研究证实。
由于目前对云杉矮槲寄生的研究仅限于其分类地位、形态特征、危害特点及其与环境条件的关系等[3-4,12-13],缺乏 对 其 侵 染 过 程 及 致 病 机 理 的 研 究,因而对云杉矮槲寄生的防治仍然处于探索阶段。尽管喷施植物生长调节剂(乙烯利)可以促进云杉矮槲寄生的花和果实的提前脱落,并且可以使其寄生芽脱落[14-15]。乙草胺等除草剂也能对云杉矮槲寄生的寄生芽具有良好的防除效果 。然而,由于药剂不能对云杉矮槲寄生内寄生系统造成明显伤害,因此难以彻底去除云杉矮槲寄生[17]。
本研究通过石蜡切片研究云杉矮槲寄生内寄生系统的发育和扩展的特点,并从组织水平上研究其与寄主互作的特点,有利于揭示云杉矮槲寄生的致病机制,为建立科学有效的防治方法提供参考。
1 材料和方法
1.1 材料采集
在青海省门源县仙米林场采集被云杉矮槲寄生侵染的青海云杉枝条作为样品,包括被侵染枝条的当年新生小枝、一年生和两年生小枝,并针对受侵染后寄主枝条的膨大部位以及与膨大部位相连的同年生非膨大部位进行采集。同时,采集云杉矮槲寄生种子萌发处于侵入阶段的枝条。所有材料均在现场切成小块进行固定、保存,以保证样品的质量。
1.2 方法
1.2.1 样品固定 将切好的小块新鲜枝条置于100mL安培瓶,用FAA 固定液(每100mL固定液中含70%乙醇溶液90mL,38%冰乙酸溶液5mL,38%甲醛溶液5 mL,另加入10 mL 丙三醇)固定,抽气后保存1 个月,备用。将固定好的样品使用70%、85%、95%和100%叔丁醇溶液进行梯度脱水。脱水后倒入已融化的石蜡,然后在60 ℃水浴锅中浸蜡1周,浸蜡完成后,石蜡包埋并修整蜡块备用。
1.2.2 切片的制备 莱卡RM2235石蜡切片机上切片,切片厚度为12μm,以明胶甘油溶液作粘贴剂,4%甲醛溶液作浮载剂。将粘贴有植物切片的载玻片放入60 ℃的烘箱中烘烤12h。
1.2.3 切片的染色和显微观察 用二甲苯和100%、95%、70%3个梯度的乙醇溶液对切片进行依次脱蜡,然后进行复水和染色。使用番红-固绿染色时,先用1%番红溶液(将1g番红先溶于少量蒸馏水中,然后加蒸馏水定容至100mL)染色20min后,用35%、50%、70%、80%4种梯度的乙醇溶液适当褪色,再用0.5%固绿溶液(将0.5g固绿先溶于少量95%乙醇中,然后加95%乙醇定容至100 mL)进行复染10s,然后用100%乙醇溶液褪色;使用甲苯胺蓝染色时,采用上述二甲苯和梯度乙醇溶液的方法脱蜡,复水后用0.05%甲苯胺蓝水溶液染色30min,然后用0.5%冰乙酸溶液褪色。最后使用二甲苯透明,中性树胶进行封片保存。使用莱卡DM1000显微镜进行观察,并使用Canon DS126231照相机拍照记录。
2 结果与分析
2.1 云杉矮槲寄生的侵入过程
云杉矮槲寄生的种子经弹射后,通过其表面槲寄生素(图1,A)粘附在寄主枝条和针叶基部(图1,B)。在云杉矮槲寄生的种子萌发过程中,胚根突破内果皮,随后趋向寄主枝条方向生长(图1,C)。
当云杉矮槲寄生的胚根接触到寄主枝条表面时,胚根顶端产生吸盘(图1,D),吸盘能够将胚根紧紧固着在寄主的枝条上。在吸盘的下部产生楔形的侵染钉(图1,D),侵染钉穿透寄主的表皮层侵入到寄主组织中。在侵染钉接触到寄主的表皮层时,表皮层会出现加厚现象,可能用于抵抗侵染钉的侵入(图1,C)。
侵染钉侵入寄主一段时间后,侵染点周围的寄主枝条逐渐膨大(图1,E),表明云杉矮槲寄生成功的侵入并和寄主建立了寄生关系。和相邻非膨大部位枝条(图1,G)相比,寄主膨大部位枝条内部的皮层细胞数量显著增加,但细胞大小无明显差异(图1,F)。
2.2 云杉矮槲寄生的内寄生系统
2.2.1 皮层根 侵染钉侵入到寄主的皮层后,定向的穿过寄主的皮层、韧皮部和形成层,侵入到寄主的木质部,形成云杉矮槲寄生的初生吸器(图2,A)。生长在皮层中的初生吸器,靠近寄主次生韧皮部的部分,其四周的细胞分裂产生云杉矮槲寄生的皮层根(图2,A)。从寄主枝条的横切面可以看出,随着皮层根细胞的分裂生长,皮层根逐渐形成一个以小细胞为核心周围被较大的细胞围绕的类似圆形的结构,其内部细胞逐渐木质化(图2,B)。从寄主枝条的纵切面可以看出,皮层根细胞排列规则,其细胞长度是宽度的2~4倍(图2,C)。在皮层根延伸的过程中,皮层根外围的细胞能够进行分裂,产生新的皮层根(图2,D),使得云杉矮槲寄生内寄生系统的皮层根结构更加复杂,在寄主皮层内形成庞大的皮层根网状系统(图2,E)。此外,还发现当在两年生和一年生枝条的节间能够观察到云杉矮槲寄生的寄生芽时(图2,F),其皮层根已经扩展到寄主的一年生枝条(图2,G),但在当年生枝条内没有发现皮层根。
在皮层根生长的过程中,皮层根的尖端细胞,特别是生长到寄主形成针叶部位的皮层根尖端细胞,可以沿着针叶生长的方向,穿过寄主皮层细胞间隙向寄主的表皮层方向生长,随后穿透寄主的表皮层,生长到寄主的外部,分化成云杉矮槲寄生的寄生芽(图3,A~E)。从寄主枝条的纵切面观察,皮层根生长方式类似于歪J形结构(图3,F),这表明每一段皮层根只有抽发产生一次寄生芽的能力。当皮层根顶端的细胞突破寄主表皮层时,表皮层也会出现加厚现象(图3,A、E)。
2.2.2 吸根 部分皮层根靠近(图4,A)或接触(图4,B)寄主的次生韧皮部时,皮层根下部的细胞通过分裂产生云杉矮槲寄生的吸根。吸根尖端的单细胞能够定向的穿透寄主的皮层、韧皮部和形成层侵入到寄主的木质部(图4,C),并沿着寄主的木射线方向通过挤压寄主的木射线细胞生长(图4,C~G)。吸根最深能够侵染到寄主的髓附近(图4,H)。观察被云杉矮槲寄生侵染的寄主枝条纵切面,在健康的木射线中寄主细胞是单层排列的,被侵染的木射线中吸根细胞和寄主木射线细胞混合生长,呈多层排列的现象。随着吸根的分裂生长,形成一个楔形结构(图4,A)。
3 讨 论
云杉矮槲寄生侵染寄主后,与寄主建立寄生关系,从寄主体内获取水分和营养物质[4],影响寄主的生理和形态[3-4],最终造成寄主树势衰弱甚至死亡。本研究结果表明,云杉矮槲寄生种子粘附在寄主枝条上,萌发过程中产生胚根,胚根接触寄主表面后产生吸盘,在吸盘下方产生侵染钉,通过侵染钉穿透寄主表皮层侵入到寄主内部,并继续向木质部延伸形成初生吸器;在皮层内的初生吸器产生皮层根,皮层根在寄主皮层内形成沿木射线向木质部扩展的吸根,同时,皮层根继续延伸直至穿透寄主表皮层形成寄生芽。该过程与已报道的其它矮槲寄生的侵染过程一致[2,7],同时发现在云杉矮槲寄生侵染钉及其皮层根顶端细胞接触寄主表皮层时,寄主的表皮层均出现加厚现象,表明寄主可能对云杉矮槲寄生的侵染产生防卫反应。
矮槲寄生侵染寄主后,在所引起的膨大和丛枝部位中,细胞分裂素、吲哚乙酸的含量增加很多[18-19],引起寄主体内激素的分布异常,从而使受侵染枝条丧失顶端优势,改变寄主对营养物质的分配[2]。本研究中,云杉矮槲寄生对寄主发生侵染时,在侵染点附近引起寄主皮层细胞数量显著增加,从而导致寄主枝条膨大,该现象也可能是由于青海云杉受到云杉矮槲寄生侵染后,枝条内部植物激素的含量发生变化所导致。然而,尚不能确定在这个过程中矮槲寄生是通过合成激素导致其含量的增加还是仅仅改变了寄主体内激素的分配[2],仍需继续深入研究证实。

图1 云杉矮槲寄生的侵染过程(C、D 为横切面)co.皮层;es.胚乳;hf.固着器;hp.寄主表皮层;pp.侵染钉;ra.胚;sx.次生木质部;A.带槲寄生素的种子,箭头表示胶状透明的槲寄生素;B.生长在针叶基部的云杉矮槲寄生种子;C.种子萌发产生胚,箭头表示寄主表皮层加厚;D.萌发的种子产生胚、固着器和侵染钉;E.在侵染点处膨大的枝条;F.枝条膨大处的寄主皮层;G.和膨大枝条同年生枝条非膨大处的寄主皮层Fig.1 The infection process of A.sichuanensein transverse section(C,D)co.Cortex;es.Endosperm;hf.Holdfast;hp.Host periderm;pp.Penetration peg;ra.Radical;sx.Secondary xylem;A.Seed with viscin at unlabeled arrow;B.Seed at the base of a needle;C.Germinated seed with radical,host periderm thickened at unlabeled arrow;D.Germinated seed with radical,holdfast and penetration peg;E.Swollen branch at the point of infection;F.The cortex in swollen branch;G.The cortex in non-swollen branch connected with swollen branch

图2 云杉矮槲寄生的皮层根横切面(B、G)和纵切面(C~E)b.皮层根;ph.初生吸器;x.皮层根的木质部;A.初生吸器在皮层内产生皮层根;B.皮层根产生木质部;C.皮层根细胞呈并排生长;D.皮层根通过分裂产生新的皮层根;E.皮层根大量分裂;F.一年生和两年生寄主枝条节间生长出云杉矮槲寄生寄生芽;G.一年生寄主枝条皮层内生长的皮层根Fig.2 Bark strands of A.sichuanensein transverse sections(B,G)or longitudinal sections(C,D,E)b.Bark strand;ph.Primary haustorium;x.Xylem of bark strand;A.Primary haustorium with bark strands formed around the host cortex;B.Bark strands with xylem;C.Bark strand to show tiered arrangement of cells;D.New bark strands formed by cells division of bark strands at unlabeled arrow;E.Many bark strands formed;F.Shoots of A.sichuanensein the internode between 1year-old branch and 2year-old branch at unlabeled arrow;G.Bark strands in the cortex of 1year-old branch
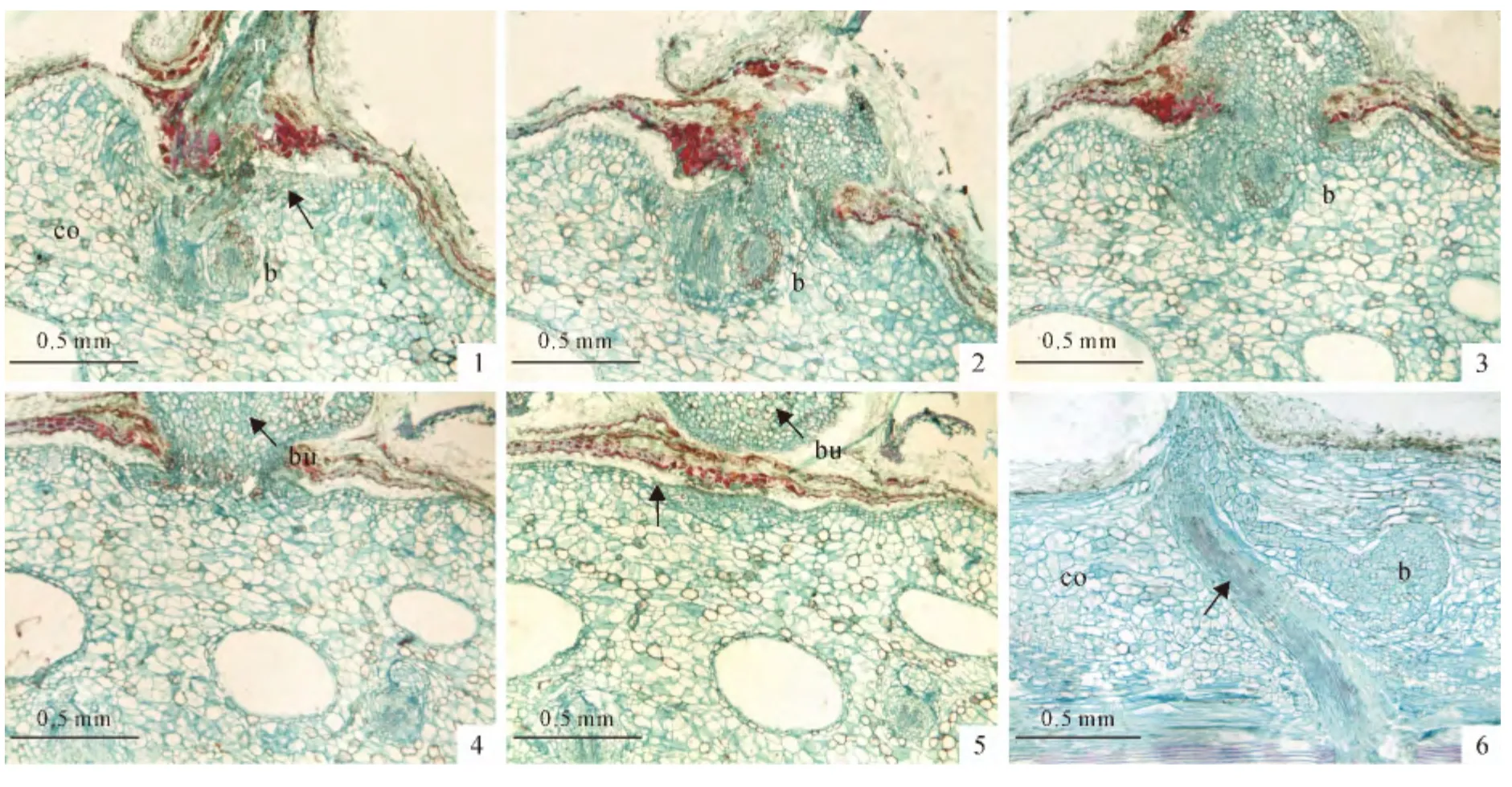
图3 云杉矮槲寄生寄生芽的产生过程(A~E为横切面;F为纵切面)b.皮层根;co.皮层;n.针叶;bu.寄生芽;A、E.未标记的箭头表示寄主的表皮层加厚;F.箭头表示皮层根沿着寄主针叶维管束方向生长Fig.3 Process of A.sichuanense shoots produced in transverse sections(A-E)or longitudinal section(F)b.Bark strand;co.Cortex;n.Needle;bu.bud;A,E.Host periderm thickened at unlabeled arrow;F.Bark strands extend along vascular of needle in the cortex at unlabeled arrow

图4 云杉矮槲寄生的吸根(A~D、H 为横切面;E、F为切向切面;G 为径向切面)b.皮层根;h.寄主细胞;ir.侵染射线;p.寄生物细胞;pi.髓;xr.木射线;s.吸根;sph.次生韧皮部;ur.非侵染射线;A、B.皮层根产生吸根;C、D.沿着寄主木射线方向生长的根吸;E.生长寄主木质部内的侵染射线和非侵染射线;F.侵染射线内的寄主和寄生物细胞;G.径向切面中寄主和寄生物的细胞;H.生长到寄主髓附近的吸根Fig.4 Sinkers of A.sichuanensein transverse sections(A-D,H)or tangential sections(E-F)or radial section(G)b.Bark strand;h.Host cells;ir.Infected ray;p.Paradise cells;pi.Pith;xr.Xylem ray;s.Sinker;sph.Secondary phloem;ur.Uninfected ray;A,B.Bark strand with sinker initiated;C,D.Sinker extended along the xylem ray;E.Infected and uninfected rays in xylem;F.Infected ray showing host and paradise cells;G.Paradise cells and host cells in radical section;H.Sinker extended into pith
矮槲寄生依赖由分布在寄主皮层内的皮层根和镶嵌在寄主木质部中的吸根组成的内寄生系统与寄主建立牢固的寄生关系[8]。本研究发现,当云杉矮槲寄生成功侵入寄主组织后,在靠近寄主次生韧皮部的初生吸器细胞分化形成皮层根,然后通过挤压寄主皮层细胞在寄主枝条中扩展,而且在外观无寄生芽的受侵染寄主枝条的一年生小枝内部,观察到云杉矮槲寄生内寄生系统的存在。这表明云杉矮槲寄生的内寄生系统可以在寄主枝条内部扩展,造成系统侵染。此外,A.douglasii的内寄生系统甚至可以扩展至寄主的顶芽内,随着寄主枝条的生长而扩展[10],而本研究并没有在当年生枝条中观察到云杉矮槲寄生的内寄生系统,因而有必要通过灵敏度更高的分子检测方法来检测寄主当年生枝条或顶芽中是否存在云杉矮槲寄生的内寄生系统。
云杉矮槲寄生内寄生系统的皮层根沿着寄主枝条扩展,可以向外穿透寄主表皮层,形成寄生芽。由此可见,对于矮槲寄生的防治,不仅要去除枝条外部的寄生芽,更要去除内寄生系统。目前在矮槲寄生害防治过程中所使用的药剂仅对寄生芽、花和果实具有去除效果[2,14-16,20],但由于内寄生系统与寄主组织形成一个嵌合整体,目前防治过程中所使用的药剂难以彻底杀死内寄生系统[2,17],同时在筛选矮槲寄生害生防菌的过程中已经关注对内寄生系统的影响 。因此,在防治过程中应研发具有内吸性和选择性的药剂,同时结合抚育间伐,对受侵染的枝条进行修枝处理,以减轻对寄主的危害。
此外,有报道称油杉寄生(A.chinense)的内寄生系统产生果胶质,使寄主细胞间的连接松散,细胞壁遭到破坏,从而便于其皮层根的生长蔓延[23]。本研究没有观察到云杉矮槲寄生的内寄生系统在扩展过程中对寄主细胞造成损伤,而是通过挤压皮层细胞和木射线细胞的方式进行扩展。矮槲寄生主要通过非共质体途径从寄主体内获取水分和营养物质[24-25],同时改变侵染部位的植物激素含量,从而影响 寄 主 的 形 态[2,18-19]。云 杉 矮 槲 寄 生 侵 染 是 否 对 寄主造成其它方面的损害仍需深入研究。
综上,本研究针对云杉矮槲寄生的侵染过程及内寄生系统进行了初步研究,研究结果揭示了云杉矮槲寄生内寄生系统的发育及在寄主内扩展特点,对进一步研究云杉矮槲寄生的致病机制以及建立科学有效的防治方法具有重要的参考意义。
[1] 吴征镒.中国植物志[M].北京:科学出版社,2003,5:240-245.
[2] HAWKSWORTH F G,WIENS D.Dwarf Mistletoes:Biology,Pathology,and Systematics[M].U.S.D.A.Forest Service Agriculture Handbook.709.Washington,DC,1996.
[3] GAO F M(高发明),CHEN L(陈 磊),TIAN CH M(田呈明),et al.Effect of Arceuthobium sichuanense infection on photosynthesis and transpiration of Picea wilsonii[J/OL].Acta Phytopathologica Sinica(植物病理学报),2015,45(1):14-21(in Chinese).
[4] XIA B,TIAN CH M,LUO Y Q,et al.The effects of Arceuthobium sichuanense infection on needles and current-year shoots of mature and young Qinghai spruce(Picea crassifolia)trees[J].Forest Pathology,2012,42(4):330-337.
[5] LI T(李 涛),MA M CH(马明呈),TAN J P(谭建萍).Damage situation and control Arceuthobium sichuanensein Xianmi forest area[J].Journal of Qinghai University(Nat.Sci.Edi.)(青海大学学报·自然科学版),2010,28(3):69-72(in Chinese).
[6] MA J H(马建海),HUAI W X(淮稳霞),ZHAO F Y(赵丰钰).Arceuthobium sichuanense,aparasitic plant attacking spruce in Qinghai Province[J].Forest Pest and Disease(中国森林病虫),2007,26(1):19-21(in Chinese).
[7] SCHARPF R F.Seed viability,germination and radicle growth of dwarf mistletoe in California[J].United States Forest Service Research Paper,Pacific Southwest Forest and Range Experiment Station,1970,59:18.
[8] KUIJT J.Morphological aspects of parasitism in the dwarf mistletoes(Arceuthobium)[J].University of California Publications in Botany,Berkeley,California,1960,30(5):337-436.
[9] SCHARPF R.Growth rate of the endophytic system of the dwarf mistletoe on Digger pine[J].Research Note.Pacific Southwest Forest and Range Exp,Stn..US Forest Service,1962,193:1-5.
[10] LYE D.Charting the isophasic endophyte of dwarf mistletoe Arceuthobium douglasii(Viscaceae)in host apical buds[J].Annals of Botany,2006,97(6):953-963.
[11] 周在豹.三江源云杉矮槲寄生生物学特性及防治研究[D].北京:北京林业大学,2011.
[12] SUN X L(孙秀玲),XU ZH CH(许志春),TIAN CH M(田呈明),et al.Spatiotemporal patterns of the seed rain of Arceuthobium sichuanense[J].Journal of Northwest Forestry University(西北林学院学报),2014,29(4):65-68(in Chinese).
[13] HU Y(胡 阳),TIAN CH M(田呈明),LI ZH Y(李周园),et al.The spatial distribution pattern of Arceuthobium sichuanense and its correlation with the environment in Qinghai Xianmi forestJ.Journal of Beijing Forestry University北京林业大学学报201436(1):102-108(in Chinese).
[14] ZHOU Z B(周在豹),XU ZH CH(许志春),TIAN CH M(田呈明),et al.Screening of the plant hormones causing the fruit preabscission of Arceuthobium sichuanense[J].Forest Pest and Disease(中国森林病虫),2007,26(3):39-41(in Chinese).
[15] XIA B(夏 博),TIAN CH M(田呈明),LUO Y Q(骆 有庆),et al.Flowering characteristics and chemical control of the buds of Arceuthobium sichuanense[J].Scientia Silvae Sinicae(林业科学),2010,46(4):98-102(in Chinese).
[16] ZHOU Z B(周在豹),XU ZH CH(许志春),TIAN CH M(田呈明),et al.Preliminary report of efficiency of herbicide on controlling Arceuthobium sichuanense[J].Plant Protection(植物保护),2014,4:41(in Chinese).
[17] 胡 阳.青海省云杉矮槲寄生发生的因素分析和防治研究[D].北京:北京林业大学,2013.
[18] PAPUET P J.Seasonal variation in cytokinin activity and content in two species of dwarf mistletoes and their hosts[J].Portland State University Dept of Biology,1979:864.
[19] LIVINGSTON W H,BRENNER M L,BLANCHETTE R A.Altered concentrations of abscisic acid,indole-3-acetic acid,and zeatin riboside associated eastern dwarf mistletoe infections on black spruce[J].General technical report RM Rocky Mountain Forest and Range Experiment Station,United States,Forest Service,1984:53-61.
[20] KAVOSI M R,FARIDI F,HAJIZADEH G.Effects of foliar application herbicides to control semi-parasitic plant Arceuthobium oxycedri[J].Bioscience,2012,4(2):76-80.
[21] SHAMOUN S F,RAMSFIELD T D,B.J VAN DER KAMP.Biological control approach for management of dwarf mistletoes[J].New Zealand Journal of Forestry Science,2003,33(3):373-384.
[22] RAMSFIELD T D,SHAMOUN S F,B.J VAN DER KAMP.Histopathology of the endophytic system and aerial shoots of Arceuthobium americanuminfected by Colletotrichum gloeosporioides[J].Botany,2011,90(1):43-49.
[23] YOU M P(游鸣佩),TONG J(童 俊).Study on biology of Arceuthobium chinense and its harm set to Keteleeria evelyniana[J].Journal of Southwest Forestry College(西南林学院学报),1987,1:38-46(in Chinese).
[24] ALOSI M C.The curious anatomy of Arceuthobiumin regards to host/parasite water relations and translocation in proceedings of the Twenty-sixth Annual Western International Forest Disease Work Conference[R].Tucson,AZ,1978.
[25] ALOSI M C,CALVIN C L.The ultrastructure of dwarf mistletoe(Arceuthobiumspp.)sinker cells in the region of the host secondary vasculature[J].Canadian Journal of Botany,1985,63(5):889-898.
