秦岭佛坪自然保护区短柄枹栎种群的数量特征
2015-07-04黄雅昆王得祥张洪武郭廷栋胡有宁
黄雅昆,王得祥*,张洪武,2,郭廷栋,胡有宁
(1 西北农林科技大学 林学院,陕西杨陵712100;2 甘肃省武威市凉州区特色林果产业中心,甘肃武威733000)
种群是生态系统的基本组成部分,也是森林生态学研究的一个重要层次[1]。种群动态作为种群生态学研究的核心,可以反映种群在时空上的数量变化规律。种群数量特征通常包括植物种群的年龄结构、生命表和存活曲线及谱分析等研究内容,不仅能够反映种群的现实状况,重建过去的干扰历史,还能够预测未来的发展趋势[2-8],为种群保护及恢复措施的制定提供科学依据。
短柄枹栎(Quercusglanduliferavar.brevipetiolata)属壳斗科栎属,是亚热带及暖温带山地常见的落叶乔木树种,尤其在长江中下游一带广泛分布,其分布范围北自山东、河南,南至贵州、四川、广东,东起安徽、江西、浙江,西达陕西、甘肃等地,一般位于在海拔600~2 000 m 的地段[9]。在佛坪自然保护区,短柄枹栎林主要分布于海拔1 300~1 700 m,其下限连栓皮栎林,大体处在锐齿槲栎林带内,但上限较锐齿槲栎林低,是栓皮栎和锐齿槲栎林之间的过渡带群落[10]。近年来,关于短柄枹栎林的研究主要集中在种群遗传多样性[11-13]、病虫害防治[14]、物种组成[15]、群落结构[10,15]及群落分类[10]等方面,对其种群数量特征方面的研究较少[16]。
短柄枹栎林多分布于山坡中上部的阳坡或山脊两侧,且坡度较大(多在35°~50°之间),这些地段土层较薄,土壤水分、养分易流失,植被稀疏,其上分布的短柄枹栎林具有防风固沙、保持水土、涵养水源等重要的生态作用,短柄枹栎种群的稳定性直接影响到其生态功能的发挥。此外,短柄枹栎林下的优势种之一——巴山木竹(Bashaniafargesii),是大熊猫冬季食物的主要来源[17-18],短柄枹栎种群的发展及动态可能对大熊猫的数量及分布造成一定的影响。短柄枹栎种群的数量特征研究无疑是分析其稳定性及发展趋势的基础工作,为此,本研究以佛坪短柄枹栎种群为研究对象,在样地调查的基础上,利用“空间代替时间”的方法,编制短柄枹栎种群静态生命表,探讨了短柄枹栎种群的数量动态,同时应用谱分析方法揭示其种群生长的周期性变化,旨在为佛坪自然保护区短柄枹栎种群的保护提供基础资料。
1 研究区自然概况
陕西佛坪国家级自然保护区(107°41′~107°55′E,33°32′~33°43′N)位于秦岭中段南坡的陕西省佛坪县岳坝乡境内,属亚热带向暖温带过渡地带,总面积29 240hm2。地势西北高,东南低,海拔900~2 904 m,全年平均气温11.4~13℃,年降水量1 000mm 左右,土壤由低山至亚高山逐渐过渡,依次为黄棕壤(1 500 m 以下)、棕壤(1 500~2 300 m)、暗棕壤(2 300~2 700m)和草甸土(2 700m 以上)[19]。保护区内植被类型以森林为主,表现出显著的垂直分布格局,自下而上依次为落叶阔叶林和针叶林,海拔2 600m 以上分布有斑块状亚高山灌丛和草甸。短柄枹栎林主要分布于海拔1 300~1 700m 的阳坡和山脊两侧,群落层次结构简单清晰,可分为乔木层、灌木层和草本层。乔木层盖度为80%,除短柄枹栎外尚有栓皮栎(Quercusvariabilis)、板栗(Castaneamollissima)、茅栗(Castanea seguinii)、锐齿槲栎(Q.alienavar.acutiserrata)、山杨(Populusdavidiana)、漆 树(Toxicodendron vernicifluum)、青蛙皮槭(Acergrosseri)等。灌木层盖度变化较大,一般为20%~90%,视生境而定,主要种类有美丽胡枝子(Lespedezaformosa)、巴山木竹、无梗越桔(Vacciniumhenryi)、阔叶荚蒾(Viburnumlobophyllum)、短枝六道木(Abeliaengleriana)等。草本层总盖度为10%~35%,常见种类有大披针薹草(Carexlanceolata)、吉祥草(Reineckia triandra)、疏花野青茅(Deyeuxiaarundinaceavar.laxiflora)、麦冬(Ophiopogonjaponicus)、陕西蛾眉蕨(Lunathyriumgiraldii)、鸭儿芹(Cryptotaeniajaponica)、暗绿蒿(Artemisiaatrovirens)、鹿蹄草(Pyrolacalliantha)、羽叶长柄山蚂 蝗(Podocarpiumoldhami)、透骨草(Phrymaleptostachya)等[20]。
2 研究方法
2.1 样地调查
在研究区海拔为1 300~1 700 m 范围内的山脊两侧及上坡位选取具有代表性的短柄枹栎林,建立面积为20m×25m 的样地共计8块,样地概况见表1。以5m×5m 为基本单元,用相邻格子法进行每木检尺,记录样地内全部乔木树种的胸径、树高、枝下高、冠幅等,并采用网格进行单株定位,灌木和草本层记录植物种名、株(丛)数、高度、盖度等。
2.2 数据处理
2.2.1 种群年龄结构划分 采用“空间代替时间”法,以径级结构代替年龄结构。根据短柄枹栎生活史特点及调查资料,将短柄枹栎种群划分为11个径级,每级间隔5cm,即第1径级为0~5cm,对应第Ⅰ龄级,第2径级为5~10cm,对应第Ⅱ龄级,以此类推[3]。

表1 样地概况Table 1 Main characteristics of selected sites
2.2.2 编制静态生命表 短柄枹栎生命表主要包括:x为年龄级;ax为x龄级开始时的个体存活数目;lx为x龄级开始时的标准化存活个体数;dx为x到x+1到龄级间的标准化死亡数;qx为x到x+1龄级间的死亡率;Lx为x到x+1龄级间的平均存活个体数;Tx为x龄级及以上各龄级的存活个体总数;ex为进入x龄级个体的平均生命期望;Kx为种群消失率,Sx为x到x+1龄级间的存活率[7,14]。生命表中的各项都可通过野外调查的ax计算获得,计算关系如下:

同时,为了更好地分析短柄枹栎种群的动态,引入生存分析中的4个函数:生存函数S(t)、累计死亡率函数F(t)、死亡密度函数f(t)和危险率函数λ(t)[6-7],计算公式如下:

2.2.3 谱分析 谱分析是研究植物种群周期性年龄更替的重要数学工具。短柄枹栎种群天然更新过程的动态是通过不同龄级的株数分布波动表现的,因此,对短柄枹栎种群进行数量动态的谱分析。谱分析是Fourier级数的展开。Fourier证明过复杂的周期现象可以由不同振幅和相应的谐波组成,写成正弦波形式为:

式中,A0为周期变化的平均,确定了种群波动的基线;Ak(k=1,2,3,…,p;p=n/2)为各谐波的振幅,反映各周期作用的大小;ωk为各谐波的频率;θk为各谐波的相角;Nt为t时刻种群大小[5-6]。
将种群各年龄个体分布视为一个时间系列t,以Xt表示t年龄序列时个体数;n为系列总长度;p=n/2为谐波的总个体数;T为正弦波的基本周期即时间系列t的最长周期。根据以下公式能够计算出ak,bk,Ak:

3 结果与分析
3.1 短柄枹栎种群年龄级结构
佛坪短柄枹栎种群可划分为11个龄级(图1),所调查个体总数为818,其中Ⅰ龄级的个体数量为544,占总数的66.50%,Ⅱ~Ⅶ龄级的个体数为260,占总数的31.78%,Ⅷ~Ⅺ龄级的个体数为14占总数的1.72%。可见,佛坪自然保护区的短柄枹栎种群以中幼林为主体,老龄个体较少,龄级结构呈倒J型,属增长型种群。
3.2 短柄枹栎种群数量动态
3.2.1 静态生命表 静态生命表反映的是多个世代重叠的年龄动态历程中的一个特定时间,并不是对同生群全部生活史的追踪[6],在静态生命表编制过程中可能会出现死亡率为负的情况。本研究采用匀滑技术[21]对统计数据ax(表2)进行处理。

图1 佛坪短柄枹栎种群年龄结构Fig.1 The age structure of Q.glandulifera var.brevipetiolata population in Foping
根据调查资料,在第Ⅱ龄级和第Ⅷ龄级的数据发生波动,分别小于第Ⅲ龄级和第Ⅸ龄级的存活数。根据静态生命表的假设,年龄组合稳定,各年龄的比例不变。由此,确定2个区段:第Ⅱ~Ⅵ龄级和第Ⅶ~Ⅺ龄级,分别计算2个区段存活数的累积:

由表1可以看出:(1)短柄枹栎种群死亡率、亏损率与危险率的变化趋势基本相似。种群在第Ⅰ龄级的死亡率最高,第Ⅱ~Ⅴ龄级相对稳定,第Ⅵ龄级出现第二个峰值,并在第Ⅶ龄级以后又逐渐上升。说明短柄枹栎种群幼苗死亡率较高,这与第Ⅰ龄级的种群消失率最大相一致。(2)短柄枹栎的种群生命期望在第Ⅱ龄级时达到最大,随后逐渐减小并伴随波动。说明短柄枹栎在第Ⅱ龄级的生存能力最强。(3)短柄枹栎种群的最高生命期望值紧随最高死亡率的龄级之后,这说明在自然筛选和种内竞争之后,存活下来的个体具有较高的期望寿命。

表2 佛坪短柄枹栎种群静态生命表Table 2 The time-specific life table of Quercus glandulifera var.brevipetiolata population in Foping
3.2.2 存活曲线和死亡率曲线 根据佛坪短柄枹栎种群静态生命表,以龄级为横坐标,存活个体数的对数Inlx为纵坐标绘制存活曲线(图2)。由图2可以看出,短柄枹栎种群存活率在第Ⅰ龄级显著下降,第Ⅱ~Ⅴ龄级呈相对平稳的下降趋势,从第Ⅵ龄级开始表现为明显下降。曲线最大斜率出现在第Ⅰ龄级,表明短柄枹栎早期死亡数较高;第Ⅵ龄级处明显下降可能是随个体增长,种内和种间竞争加剧的结果。
Deevey将存活曲线分为3种类型:Ⅰ型存活曲线呈凸型,种群在达到生理寿命之前只有少数个体死亡,大部分个体都能活到生理寿命,即在生命末期死亡率才升高;Ⅱ型存活曲线呈对角线型,种群死亡率在各发育阶段大致相等;Ⅲ型存活曲线呈凹型,幼体死亡率很高,只有极少数个体能够活到生理寿命。总体上看,短柄枹栎种群存活曲线介于Deevey-Ⅱ型和Deevey-Ⅲ型之间。Hett和Loucks在检验估算的存活状况是符合Deevey-Ⅱ型曲线还是Deevey-Ⅲ型曲线时,采用2 种数学模型进行检验,即指数函数Nx=N0e-b用以描述DeeveyⅡ型存活曲线,幂函数式Nx=N0x-b描述Deevey-Ⅲ型存活曲线[22]。本研究采用上述2种模型对短柄枹栎种群存活曲线类型进行检验,建立其相应模型:
指数函数拟合结果:Nx=8.842 6e-0.1748x,F=43.957,R=0.911

图2 佛坪短柄枹栎种群存活曲线Fig.2 Survivorship curve for Q.glandulifera var.brevipetiolata population in Foping
幂函数拟合结果:Nx=9.2152x-0.6852,F=16.247,R=0.802
由于短柄枹栎种群指数模型的检验值及相关系数值均大于幂函数的检验值及相关函数值,可以认为短柄枹栎种群存活曲线更趋近于Deevey-Ⅱ型,说明佛坪短柄枹栎种群目前处于相对稳定的状态。
死亡率曲线能够反映种群死亡率的动态变化,以龄级为横坐标,死亡率和危险率为纵坐标绘制短柄枹栎的种群死亡率及种群消失率曲线(图3)。由图3可见,短柄枹栎种群在第Ⅰ龄级时的死亡率最高,说明低龄级的短柄枹栎死亡率很高,环境筛选强度大,只有少数幼苗能够通过筛选进入第Ⅱ龄级;从第Ⅱ~Ⅴ龄级,死亡率比较稳定,随年龄的增长,营养发育过渡阶段有一定的筛选强度;随后在第Ⅵ龄级出现第二个峰值,可能是由于随个体不断生长,对环境的需求也不断增加,种内和种间竞争加剧,导致死亡率升高;第Ⅶ~Ⅷ龄级死亡率低于第Ⅵ龄级并保持稳定,第Ⅵ龄级后存活下来的个体,生存环境得到改善,竞争强度降低,使种群死亡率维持在较低水平;第Ⅸ~Ⅺ龄级死亡率又陡然升高,种群内个体大量死亡,说明此时已到达短柄枹栎的生理年龄。种群消失率曲线的趋势与死亡率曲线基本相似。
3.3 短柄枹栎种群周期性变化规律
谱分析过程同样以“空间代替时间”的方法,用径级代替年龄级,数据长度n即为龄级数(n=11),基波的基本周期年限为n×5=55(5为龄级间隔)。由于调查数据各龄级个体数相差较大,在计算时采用对数化处理,即以X′t=ln(Xt+1)代换公式中的Xt。利用谱分析的公式计算各种情况下各波形的振幅Ak(k=1,2,3,…,p;p=n/2),结果见表3,其中A1为基波,A2~A6为各个谐波,每个谐波的周期分别为基本周期的1/2,1/3,1/4,…,1/p,Ak值的大小差异反映了各周期作用大小的差别。
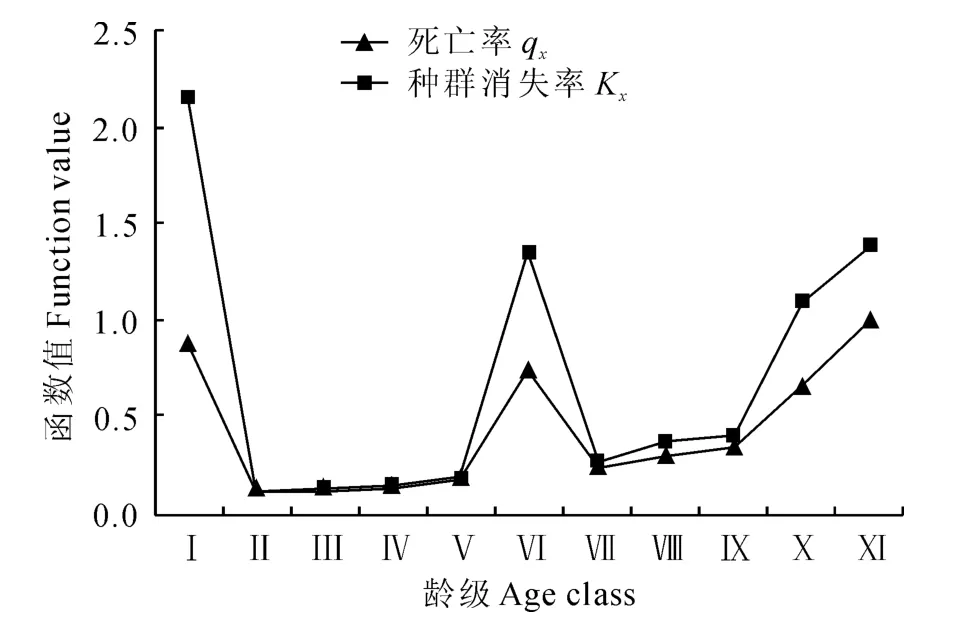
图3 佛坪短柄枹栎种群死亡率(q x)及种群消失率(K x)曲线Fig.3 Mortality curve and killing power curve of Q.glandulifera var.brevipetiolata population in Foping

表3 佛坪短柄枹栎种群周期性波动Table 3 Periodic fluctuation of Q.glandulifera var.brevipetiolata population in Foping
基波表现了基本周期的波动,其周期长度为种群本身所固有,由种群波动特性决定[2]。由表3可见,Ak值中A1为最大,这说明短柄枹栎生命周期的生物学特性对其种群动态变化的影响起主导作用。由于所调查的时间系列长度不足以表现出基本周期,无法准确得到基波的周期长度,但其存在性是肯定的。此外,A3处的波动表明除基波的影响外,短柄枹栎种群动态还存在小周期波动,此处波动的周期为基本周期的1/3,对应的空间序列胸径约为18 cm,对应第Ⅳ龄级,与短柄枹栎林木高生长进入主林层的阶段相吻合,可能是林木产生分化时的数量波动。
4 讨 论
本研究发现短柄枹栎种群呈稳定增长趋势,支持岳明等[10]对佛坪短柄枹栎林的研究结论。种群死亡率极值分别出现在第Ⅰ龄级和第Ⅵ龄级。第Ⅰ龄级处的峰值说明短柄枹栎幼苗的死亡率较高,这与许多学者对栎类的研究结果相似[3,23-24]。与大多数栎类树种一样,短柄枹栎有较强的萌蘖更新能力[9],这一方面有利于其种群的发展,另一方面也使幼苗分布较为密集,增加了个体间的竞争。此外,山脊日照强烈,气候干燥,风力强劲,土层瘠薄,林下腐殖质聚集少,一定程度上限制了幼苗的存活率。因此,这一现象可能是幼苗的密度制约性死亡与严酷环境条件共同作用的结果[24]。第Ⅵ龄级处峰值产生的原因可能是随种群的不断发育,个体对环境资源的需求增加,种内种间竞争加剧,致使一部分个体死亡,以维持种群内部的稳定。
而金则新对浙江天台山落叶阔叶林优势种群的研究显示,短柄枹栎林内天然更新困难,幼苗幼树极少,种群呈衰退趋势[16],与本研究结果存在差异,这可能是取样环境不同引起的。由于短柄枹栎适应性强,水平分布或垂直分布范围均较大,因此各地短柄枹栎林的立地条件也有较大差异[9]。生于土层较厚的山坡或沟谷两侧,林分生长发育好,但短柄枹栎喜光的特性使其在郁闭的林冠下难以完成更新。相比之下,处于山脊,立地条件较差,林分稀疏,种子萌发和幼苗形成能够获得充足的光照,天然更新的得以实现。可见,种群的发展趋势与其所处的立地条件有关,可能进一步影响到整个群落演替方向,对不同立地条件下同一树种种群动态的对比研究或许能够揭示群落的演替机制。
伍业钢等[25]首次将谱分析应用于阔叶红松林的演替及天然更新过程的研究,认为红松天然更新过程呈周期波浪式发展,并将这种现象作为稳定植物群落中优势种的特征。谱分析的结果揭示出短柄枹栎种群动态存在着周期性,基本周期较长,表明短柄枹栎种群天然更新需要较长的时间;而大周期内的小周期则说明短柄枹栎种群本身在结构上可能存在内调节机制,使其能够有效配置资源,以利于种群的发展。可见,谱分析的结果反映了短柄枹栎种群结构的复杂性和种群发展的相对稳定性。
综上所述,虽然短柄枹栎在幼苗到幼树的发育过程中存在一定的障碍,但整体而言,并未对其种群的发展产生消极影响。同时,通过种内竞争,存活下来个体能够获得足够的环境资源,使种群得以持续发展。可见,佛坪短柄枹栎种群目前处于稳定增长状态。此外,植物种群数量特征是个体生存能力与外界环境相互作用的结果,对种群进行保护、恢复和发展的同时应结合环境条件综合考量。
[1]KIMMINS J P.Forest ecology:a foundation for sustainable management[M].Prentice-Hall Inc.,1997:367.
[2]LIN Y Y(蔺雨阳),WANG D X(王得祥),LEI R D(雷瑞德),etal.Study on the characteristics ofPinustabulaeformisCarr.population on the western section of Qinling Mountains[J].ResearchofSoilandWaterConservation(水土保持学报),2009,16(4):180-185(in Chinese).
[3]TIAN L(田 丽),WANG X A(王孝安).Population dynamics analysis of tree speciesQuercusliaotungensisin Malan forest region on Loess Plateau[J].ActaAgriculturaeJiangxi(江西农业学报),2011,2(2):39-42(in Chinese).
[4]WU CH ZH(吴承祯),HONG W(洪 伟),XIE J SH(谢金寿),etal.Life table analysis ofTsugalongibracteatapopulation[J].Chinese JournalofAppliedEcology(应用生态学报),2000,11(3):333-336(in Chinese).
[5]LI X K(李先琨),SU Z M(苏宗明),XIANG W SH(向悟生),etal.Study on the structure and spatial pattern of the endangered plant population ofAbiesyuanbaoshanensis[J].ActaEcologicaSinica(生态学报),2002,22(12):2 246-2 253(in Chinese).
[6]BI X L(毕晓丽),HONG W(洪 伟),WU CH ZH(吴承祯),etal.Population statistics analysis ofPinustaiwanensis[J].ScientiaSilvae Sinicae(林业科学),2002,38(1):61-67(in Chinese).
[7]LI L(李 玲),ZHANG G F(张光富),WANG R(王 锐),etal.Life table of naturalGinkgobilobapopulation in Tianmu Mountain Nature Reserve[J].ChineseJournalofEcology(生态学杂志),2011,30(1):53-58(in Chinese).
[8]LI J J(李进军),XUE B R(薛斌瑞),CHAI Z ZH(柴宗政).Quantity dynamics and distribution pattern forHaloxylonammodendronpopulation of the desert-oasis ecotone in Minqin County of Gansu Province,China[J].JournalofNortheastForestryUniversity(东北林业大学学报),2013,41(8):27-31(in Chinese).
[9]吴中伦,徐化成,管中天.中国森林[M].北京:中国林业出版社,1998:1 211-1 212.
[10]YUE M(岳 明),REN Y(任 毅),GU T Q(辜天琪),etal.A preliminary study ofQuercusglanduliferavar.brevipetiolataforest in Foping National Reserve[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2001,21(1):146-153(in Chinese).
[11]CAO Q(曹 倩),LIU Y F(刘义飞),HUANG H W(黄宏文),etal.Restoration of genetic diversity ofQuercusglanduliferavar.brevipetiolatapopulation after artificial logging[J].JournalofTropicalandSubtropicalBotany(热带亚热带植物学报),2014,22(1):68-76(in Chinese).
[12]GU Q P(顾奇萍),JIN Z X(金则新),LI J M(李钧敏),etal.Genetic diversity ofQuercusserratavar.brevipetiolatapopulations at different succession stages[J].JournalofNorthwestForestryUniversity(西北林学院学报),2008,23(1):18-22(in Chinese).
[13]JIN Z X(金则新),LI J M(李钧敏),DING L Y(丁丽亚),etal.Genetic diversity ofQuercusglanduliferavar.brevipetiolatapopulations revealed by ISSR markers[J].BulletinofBotanicalResearch(植物研究),2007,27(5):559-563,621(in Chinese).
[14]LIU H(刘 华).Preliminary investigation on the pests ofQuercusserrataThunb.var.brevipetiolata(A.DC.)Nakai in Jinsi Valley[J].ShaanxiForestScienceandTechnology(陕西林业科技),2013,(3):50-53(in Chinese).
[15]SHA Y Y(沙迎迎),YUAN ZH L(袁志良),ZHU X L(朱学灵),etal.The species composition and community structure of 1haQuercus serratavar.brevipetiolataforest in Baotianman National Nature Reserve[J].JournalofHenanAgriculturalSciences(河南农业科学),2012,41(8):148-153(in Chinese).
[16]JIN Z X(金则新).Dominant population structure and trend of deciduous broad-leaved forest in the Tiantai Mountains of Zhejiang[J].JournalofZhejiangForestryCollege(浙江林学院学报),2001,18(3):245-251(in Chinese).
[17]YONG Y G(雍严格),ZHANG J(张 坚),ZHANG SH N(张陕宁).The distribution and number of the giant panda in the Foping reservation[J].ActaTheriologicaSinica(兽类学报),1993,13(4):245-250(in Chinese).
[18]YONG Y G(雍严格),WANG K W(王宽武),WANG T J(汪铁军).Giant panda’s moving habit in Foping[J].ActaTheriologicaSinica(兽类学报),1994,14(1):9-14(in Chinese).
[19]王宇超.秦岭大熊猫主要栖息地植物群落特征及与生境对应关系分析[D].陕西杨陵:西北农林科技大学,2012.
[20]YUE M(岳 明),REN Y(任 毅),DANG G D(党高弟),etal.Species diversity of higher plant communities in Foping National Reserve[J].ChineseBiodiversity(生物多样性),1999,7(4):263-269(in Chinese).
[21]江 洪.云杉种群生态学[M].北京:中国林业出版社,1992:11-12.
[22]SILVERTOWN J W,祝 宁.植物种群生态学导论[M].哈尔滨:东北林业大学出版社,1987,54:87.
[23]YU F(于 飞),WANG D X(王得祥),SHI X X(史晓晓),etal.Species composition and regeneration characteristics of main woody plant seedlings in a pine-oak mixed forest in Qinling Mountains[J].ActaBot.Boreal.-Occident.Sin.(西北植物学报),2013,33(3):592-598(in Chinese).
[24]BELDEN A C,PALLARDY S G.Successional trends and apparentAcersaccharumregeneration failure in an oak-hickory forest in central Missouri,USA[J].PlantEcology,2009,204(2):305-322.
[25]WU Y G(伍业钢),HAN J X(韩进轩).A spectral analysis of the population dynamics of Korean pine in the mixed broad-leavedPinus koraiensisforest[J].ChineseJournalofEcology(生态学杂志),1988,7(1):19-23(in Chinese).
