辽西走廊白狼山保护区植物区系及其与邻近山地的关系
2015-07-04张梅
张 梅
(辽东学院,辽宁丹东118003)
不同山地植物类群的性质、关系及分布格局一直是生态学和保护生物学研究的热点[1-3]。白狼山位于辽西建昌县境内,西处燕山余脉大(小)凌河的上游集水区,北靠内蒙古高原科尔沁沙地,东接辽东,南邻渤海辽东湾,同时又属半干旱地区的农林交错区,山体纺锤状呈西南至东北方向,主峰海拔1 140.2m,独特的地质地貌和气候水文条件孕育了较为丰富山地森林生态系统。保护区位于环渤海经济圈的北方,京津唐生态防护区的东北侧,丰富的森林资源成为辽西北一道天然生态屏障,有效阻挡内蒙干旱寒冷的西北强风及科尔沁沙地向环渤海地区扩展,良好植被对“环渤海经济区”和“京津唐生态圈”的生态保护有重要生态和社会效益[4]。关于本区以往多关注于花卉资源、生态评价等研究[5-7],而植物区系方面的研究未见报道。本研究对白狼山保护区种子植物区系组成、属的分布区类型及与其他山地的相似性进行分析,以揭示辽西地区与华北、华东植物区系的相似关系,为区域生物资源的可持续开发和生物多样性保护提供基础参考和科学依据。
1 自然概况
白狼山自然保护区(40°54′53.96″N,119°34′46.98″E)属燕山山脉最东侧余脉之松岭山系,是以暖温带天然落叶阔叶林、侧柏林和野生动植物种栖息地为主要保护对象的森林生态系统类型保护区。保护区总面积17 440hm2,属暖温带大陆性季风气候,年平均气温9 ℃,极端最低气温-23.9 ℃,极端最高气温40.7 ℃;年平均降水量为430 mm,春季雨量较小,多集中在6~8月,无霜期180d,年日照时数2 673h,以春季为多,夏季次之。土壤多为棕色森林土及褐土。保护区植被类型可以分为4个植被型,10个植被亚型,23个群系,主要有落叶阔叶林组成的地带性植被,还有天然侧柏(Biotaorientalis),油松(Pinustabulaeformis)等群落类型和其他杂木林群落,灌木群落有山杏(Prunnsmandshurica)、映山红(Rhododendronsimsii)和荆条(Vitex negundo)等,共同组成了保护区复杂和稳定的森林生态系统,其中,侧柏林群落是中国天然侧柏林面积最大的一处。本区处于华北、蒙古高原和长白植物区系的交汇带上,孕育了丰富的物种多样性,有国家级重点保护野生植物9种,国家级重点保护野生动物17种,是辽西干旱地区植被恢复与发展的一个优良种源基地和基因储存库。
2 研究方法
2.1 区系调查与分析
通过查阅白狼山保护区基本资料并外业调查,采用线路调查、定点辐射调查、特殊地区重点调查相结合进行植物调查和采集,GPS野外定位并以标志点为中心辐射状线路调查不同林型和生境,范围包括该区不同林型(针叶林、阔叶林和针阔叶混交林)、地貌(山顶、坡地和沟谷)和海拔生境(300~1 400 m),依据野外调查和植物鉴定等建立保护区植物数据库;运用植物区系地理学原理,研究植物科、属的区系地理成分,对本区种子植物区系分类群的组成、区系类型划分及其与相邻区系的关系进行了全面整理、定量统计和系统分析。同时选取其他11个代表山地植物属的分布型数据进行相似关系分析[8-12]。
2.2 区系相似关系
参照文献[13]计算植物区系谱的比率,计算公式为:FER=(FEi/T)×100,式中FEi为某区系第i个区系成分的分类群数(如属或种),(i=1,2,3,…,n),T为某区系各种地理成分的分类群总数,即:;聚类模型以属的地理成分为指标[14],求出分布区类型间距离矩阵,设有n个m维样本数据xij:i=1,2,…,n;j=1,2,…,m。计算各区系样本间标准欧氏距离:,把 每 个样本作为一类,将距离最近的两类归并为一个新类,采用系统聚类中组间均联法并作图,谱系图通过SPSS18.0软件获得;排序分析以主坐标法探讨各区系间的相似关系,排序图及方差计算通过PAST3.0软件获得。不同植物区系之间的相关系数,以99%的概率保证作为显著性检验指标,分析在SPSS18.0软件中完成。
3 结果与分析
3.1 植物区系多样性组成
通过外业调查结合资料统计建立植物区系数据库,本区共有野生种子植物101科394属790种(含变种、亚种和变型,不含栽培种),其中裸子植物有3科7属13 种,侧柏、油松、落叶松(Larixgmelinii)是构成针叶林的建群种。双子叶植物81科306属620种,单子叶植物17科81属157种(表1)。单种科和单属科反映了植物科进化过程中,新产生的科其属种尚未分化,还没有新衍生的属种;另为属种已经大量消亡的演化终极的科或是现存的残遗种类;单种科和单属科也反映了地区植物进化的历史和现状[15]。
3.1.1 科的多样性及区系组成 作为较高级分类单位的科反映物种间较广泛的亲缘关系,科的区系类型可揭示特定空间内植物区系特征的总体概况,探析特定植物区系与其它区系的联系以及植物区系起源的线索等。本区共有种子植物101科,其中既有进化水平较低的古老科,如麻黄科(Ephedraceae)、木兰科(Magnoliaceae)、马兜铃科(Aristolochi-aceae)等,也有植物进化中处于分化的关键类群,如虎耳草科(Saxifragaceae)、金缕梅科(Hamamelidaceae)等,还有高度进化、物种多样性分化十分复杂的菊科(Asteraceae)、禾本科(Gramineae)、兰科(Orchidaceae)等科。
表2显示,本区种子植物含41 种以上科有菊科、禾本科和蔷薇科(Rosaceae)、豆科(Leguminosae)、毛茛科(Ranunculaceae)共5科292种,分别占总科、种数的4.95%和36.96%,不足植物科数的5%,说明优势科在区系组成中具有重要作用;含2~5种的科有36科,只有1种的科有30个,如亚麻科(Linaceae)、蓝雪科(Plumbaginaceae)等;二者的比例多达65.34%,而种数只占16.08%,反映了本区种子植物中大科少,而小科较多的特点。根据吴征镒的世界种子植物科的分布区类型划分[16],白狼山种子植物科的分布型属于8个类型,其中世界广布和热带广布占68.31%,温带分布占有31.69%,以北温带分布最显著(表3)。因此,从植物科的水平来看,本区种子植物区系具有显明的温带色彩。
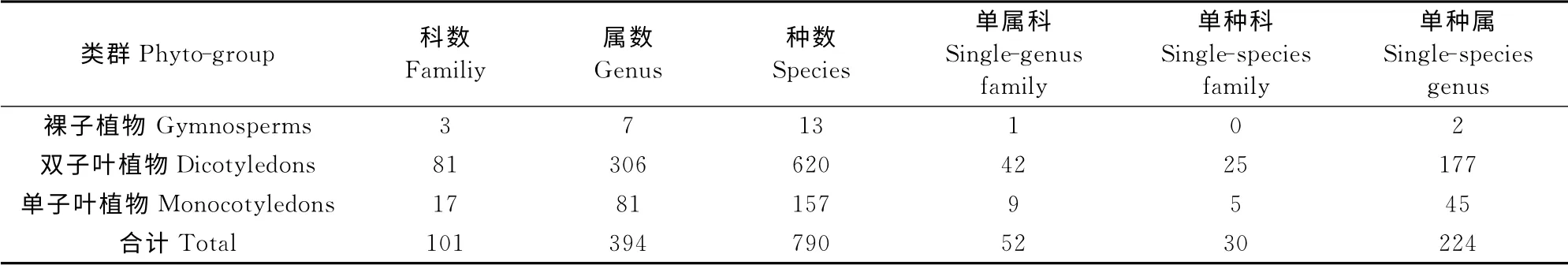
表1 白狼山野生种子植物组成Table1 Composition of plant biodiversity of seed plants in Bailang Mountain
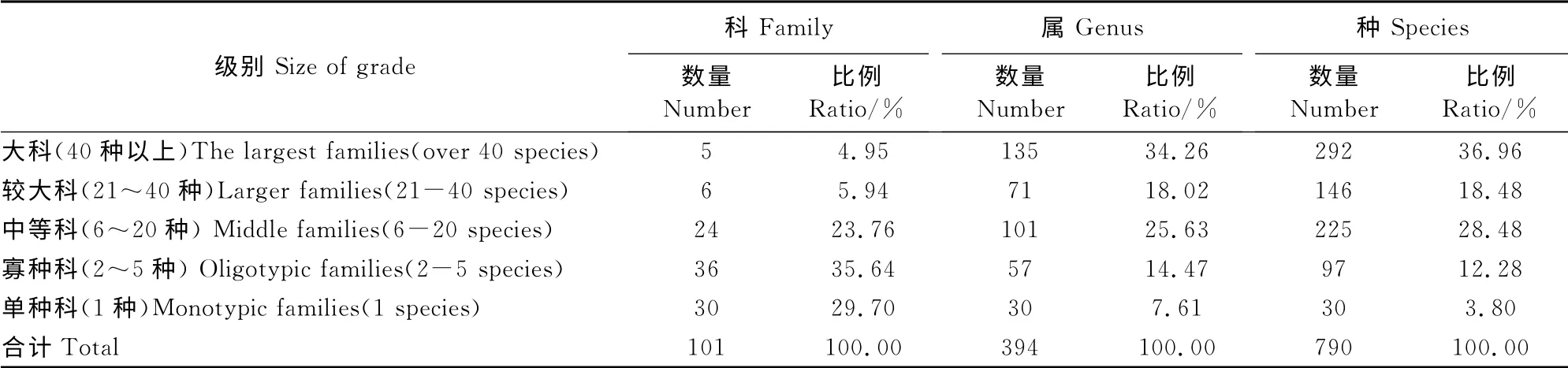
表2 白狼山种子植物科级别统计Table2 Size of families of the seed plants in Bailang Mountain

表3 白狼山种子植物科的区系分布区类型Table3 The areal-types of the families of the seed plants in Bailang Mountain
3.1.2 属的多样性组成 白狼山自然保护区内含11种以上的种子植物有蓼属(Polygonum)和堇菜属(Viola),中、小属十分丰富,含6~10种有20个属,含2~5种有148属389种,占总种数49.24%;单种属有224属,占56.85%(表4)。单种属和寡种属代表植物进化的两个方向:一个是新属建立,种属尚未分化;另一个是古老属演化终极,只有少数残遗种类。保护区内既有许多古老的残遗成分,又有一些年轻成分,反映了白狼山植物区系在进化水平上的多样性,其中桔梗属(Platycodon)、鹅肠菜属(Myosoton)、防风属(Saposhnikovia)等为世界单型属,说明植物区系分类上的孤立性和演化上的原始性。
3.2 属的区系分布类型组成
本区种子植物属的区系地理成分可划为15个分布区类型(表5),包括热带、亚热带分布共有58属,占白狼山总属数的17.27%,其中泛热带分布居首位,共有40属;其次是热带亚洲至热带非洲分布,共6属;旧世界热带分布有6属,多为草本植物和灌木,如 香 茶 菜 属(Rabdosia)、雨 久 花 属(Monochoria)、天门冬属(Asparagus)等;热带亚洲成分共有2属,苦荬菜属(Ixeris)等。温带性质分布共273属,占总属数81.30%,其中北温带成分居首位,共有116属,居该区系的第一位,许多是灌木和草本种类,木本属也比较丰富,是该地区植被的优势种和表征种,如蒙古栎(Quercusmongolica)、侧柏、紫椴(Tiliaamurensis)、核 桃 楸(Juglansmandshurica)、白桦(Betulaplatyphylla)、油松;东亚分布有25属,代表性的如猕猴桃属(Actinidia)、刺榆属(Hemiptelea)、溲疏属(Deutzia)等;该分布型的变型中,中国-日本变型占有绝大多数,表明该区系与华东、日本植物区系的关系更密切;东亚-北美间断分布有22 属,如珍珠梅属(Sorbaria)、扯根菜属(Penthorum)、胡枝子属(Lespedeza)等。中国特有成分仅有5属,都为残遗的古特有属,这与其远离中国特有属分布中心的地理位置是一致的。可见,本区植物区系地理成分复杂,种类丰富多样,东亚成分的起源过程占有重要地位,占有次要地位的热带成分表明本区区系与其的渊源和联系;以温带性质属为主,热带性质属也占有一定比例,区系应置于中国-日本森林植物亚区从暖温带向温带过渡的范围内。

表4 白狼山种子植物属级别统计Table4 Size of genera of the seed plants in Bailang Mountain

表5 白狼山种子植物区系属的分布区类型Table5 The areal-types of the genera of the seed plants in Bailang Mountain
3.3 白狼山与其它山地植物区系的相似关系
从东北、华北、华东等地区选取11个代表性山地,探讨白狼山在各山地植物区系中的位置关系。不同山地植物区系谱的相似程度规律性比较明显,长白山、帽儿山等热带成分(F2~F7)相对比例较低,而温带属(F8~F14)的比例相对较高,纬度较低的庐山和井冈山则与此相反。保护区山地植物属的分布区包含有14 个分布区类型,地理成分来源广泛,北温带分布成分(F8)明显高于其他地区,有明显的温带属性(表6)。
应用主坐标排序法得到12个山地种子植物属的分布区类型关系的排序图(图1,A),主坐标的特征根和方差贡献率是选择主坐标的依据。由表7可知,第一主坐标的方差贡献率为95.64%,第二主坐标的方差贡献率为2.19%,前两个主坐标的累积方差贡献率为97.83%,表示前两个主坐标,即影响区系分布的纬度、其他综合因子可表达全部信息的97.83%,意味着图2中只有2.17%的信息丢失。所以,选择前两个主坐标进行排序,较好显示了不同山体植物区系的相似性关系,可把12个山地种子植物属的分布区类型划分为3组(图1,A)。
聚类树状图反映了12个山地植物属的分布区类型之间的相似性关系(图1,B),其欧式距离在12的水平上可分为三类:第一类包括长白山和帽儿山,属东北植物区系;第二类包括白狼山、努鲁儿虎山、小五台山和东灵山、雾灵山先聚合一起,再与凤凰山、千山聚为一类,属华北植物区系;第三类为庐山、井冈山和铜钹山为一组,属华东植物区系。这与主坐标分组情况大致匹配。同时,相关分析表明各山地植物区系间相似程度,相关系数为0.473~0.999,白狼山与努鲁儿虎山(r=0.999,P<0.01)和东灵山(r=0.998,P<0.01)相关性较大,与井冈山(r=0.548,P<0.05)、铜钹山(r=0.654,P<0.05)相关性较小(表8),表明了与白狼山远近不同山地的相似性相对大小。
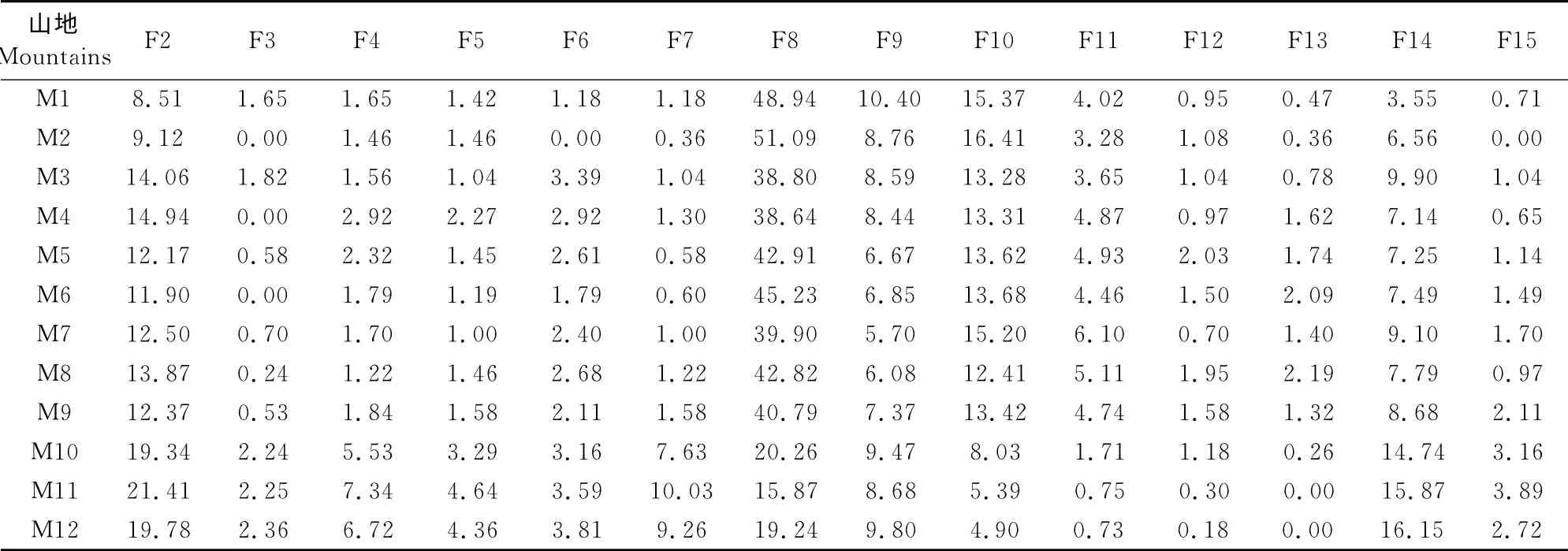
表6 白狼山等12个山地种子植物分布型谱Table6 Floristic spectrum of 12mountains flora in China

表7 区系谱前两个主坐标的方差统计率和累计方差贡献率Table7 Flora spectrum variance proportion and cumulative variance of the two principal coordinates
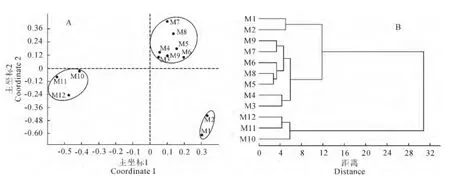
图1 白狼山等12个山地植物区系的主坐标排序图(A)和聚类树状图(B)Fig.1 PcoA ordination(A)and Cluster dendrogram(B)of 12mountains flora in China
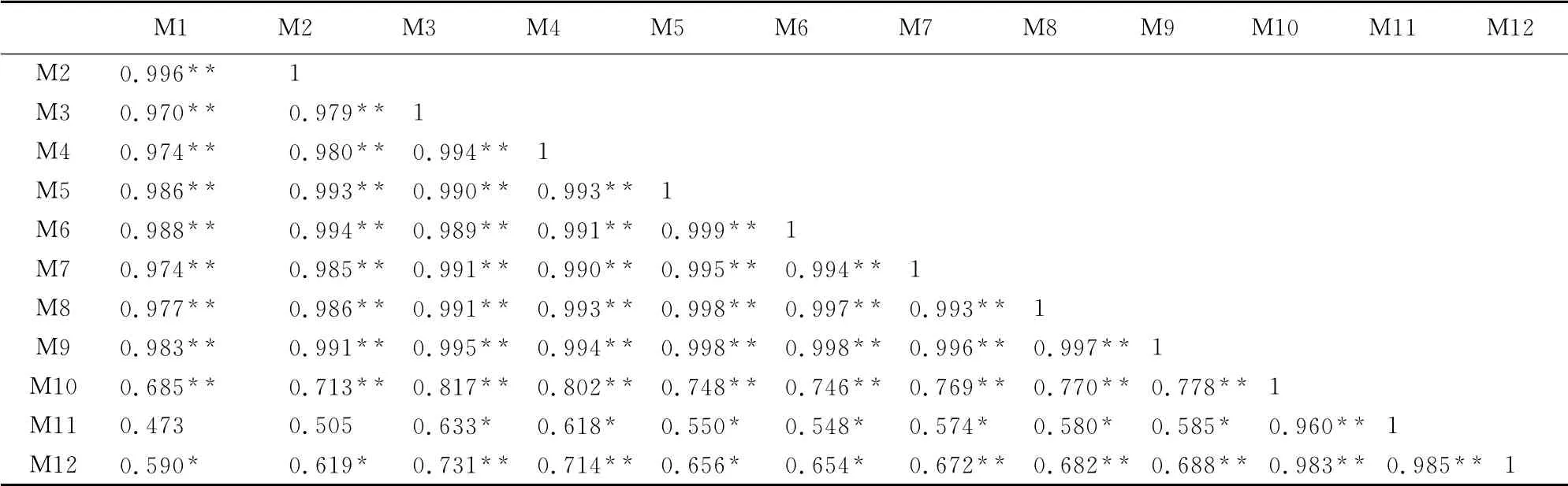
表8 白狼山等12个山地植物区系的相关性Table8 Floristic correlation matrix of 12mountains flora in China
4 讨 论
辽西低山丘陵是华北地区北端、内蒙古高原向辽河平原的过渡地带,也是植物地理的交汇区[12]。本研究表明:白狼山保护区种子植物较为丰富多样,共有野生种子植物101科394属790种,科的组成优势科现象明显,而属的组成中单种属和少种属共有372 属,占 种 子 植 物 总 属、种 数 的94.4% 和77.6%,这些属中如红豆杉属(Taxus)是典型第三纪孑遗植物,表现出植物区系演变的残遗性和古老性,是植物在进化和分化过程中对复杂生境条件适应的结果,与所处地质、地理及气候降水等有密切的关系。
在植物分类学上,属有相对稳定的形态特征和分布范围,在其进化过程中随地理环境的变化发生分异。因此,属能反映植物系统发育过程中的进化分化情况及地区性特征[17],这能反映植物区系在不同地域的类似性。研究表明:本区种子植物属的地理成分主要以温带成分总属数的78.83%,而热带成分仅占19.23%,呈现出明显的温带性质,在温带成分中以北温带分布、旧世界温带分布和东亚分布比例较高,这和在同纬度的华北山地努鲁儿虎山、雾灵山比较相似且都以温带成分为主,植物属分布类型比例相似性也接近[9]。
植物的分布不仅取决于现代生境条件,也受地质时期环境的影响[18]。华北地区地理环境的演迁以及燕山运动为主的造山运动,影响了本区植物分布及物种的形成。研究表明:白狼山与努鲁儿虎山、小五台山、东灵山及雾灵山相似关系最为密切,而与华东地区东部井冈山、庐山的关系较远。主坐标排序与聚类结果呈现出基本一致的组别划分,一定程度上反映了中国南北山地植物区系地理属性的接近程度及相似关系,支持白狼山区系划为华北平原山地亚地区较为合适。从大尺度生物地理格局分析,处于低纬度华东植物区系的庐山、井冈山和铜钹山的热带性质较为明显,而高纬度的东北植物区系的帽儿山、长白山的温带性质则相对显著。地理区域及生境因子的变化,不同山地水热环境条件的纬向差异,可能是影响南北植物区系地理相似关系的重要因素[19]。
[1] LE ROUX P C,LENOIR J,PELLISSIER L,etal.Horizontal,but not vertical,biotic interactions affect fine-scale plant distribution patterns in a low-energy system[J].Ecology,2013,94(3):671-682.
[2] HUY H,SHENG D Y,XIANG Y Z,etal.The environment,not space,dominantly structures the landscape patterns of the richness and composition of the tropical understory vegetation[J].PloSone,2013,8(11):e81308.
[3] CALABRESE J M,CERTAIN G,KRAAN C,etal.Stacking species distribution models and adjusting bias by linking them to macroecological models[J].GlobalEcologyandBiogeography,2014,23(1):99-112.
[4] WANG H SH(王荷生).华北植物区系地理[M].北京:科学出版社.1997.
[5] LI B(李 斌).Overall ecological evaluation of Liaoning Bailangshan Nature Reserve[J].ProtectionForestScienceandTechnology(防护林科技),2013(7):69-70(in Chinese).
[6] LI H SH(李怀生),LI D W(李德武).Survey and utilization of flower plants resource in Liaoning Bailangshan Nature Reserve[J].Journal ofHeilongjiangVocationalInstituteofEcologicalEngineering(黑龙江生态工程职业学院学报),2012,6:15(in Chinese).
[7] LI D W(李德武),LI H SH(李怀生).Investigation and analysis on spermatophyte of Liaoning Bailangshan Nature Reserve[J].Journalof HeilongjiangVocationalInstituteofEcologicalEngineering(黑龙江生态工程职业学院学报),2012,(5):3-5(in Chinese).
[8] MA Y X(马玉心),CAI T J(蔡体久),TAN X J(谭晓京).Comparison on floristic components between Liangshui Nature Reserve and Maoershan Nature Reserve[J].JournalofWuhanBotanicalResearch(武汉植物学研究),2007,25(1):51-56(in Chinese).
[9] MA G Q(马国青),JIANG L W(蒋丽伟).Composition and geographic elements of the flora in Nuluer Hushan National Nature Reserve,Liaoning of Northeastern China[J].JournalofBeijingForestryUniversity(北京林业大学学报),2012,34(4):136-141(in Chinese).
[10] ZHU L T(朱 立 涛),WEN G(文 光),WANG SH E(王 善 娥),etal.A comparison of the floras of Qian Mountain and adjacent mountains[J].JournalofTropicalandSubtropicalBotany(热带与亚热带植物学报),2007,15(1):77-81(in Chinese).
[11] YAN SH X(闫双喜),YANG Q SH(杨秋生),WANG P F(王鹏飞),etal.Study on the Flora Comparison of Partial Areas in China[J].JournalofWuhanBotanicalResearch(武汉植物学研究),2004,22(3):226-230(in Chinese).
[12] LIU P(刘 平),CHENG Q(程 群),LI B X(李宝祥),etal.Survey of mammalian resources in Yiwulv Mountain National Nature Reserve[J].ChineseJournalofWildlife(野生动物),2008,29(2):66-68(in Chinese).
[13] LIU L(刘 利),HUANG G H(黄国 辉),ZHANG M(张 梅),etal.Floristic compositions of spermatophyte and the similarity with other moutain land in Xianren Cave Natural Reserve in Liaoning Province[J].JournalofNortheastNormalUniversity(Natural Science Edition)(东北师大学报·自然科学版),2014,46(4):116-121(in Chinese).
[14] LIU L(刘 利),ZHANG M(张 梅),KANG F F(康峰峰).A comparison of the floras of Bashilazi Reserve and aduacent mountains[J].ActaBoreal.-Occident.Sin.(西北植物学报),2013,3:607-612(in Chinese).
[15] TONG Y(童 毅),JIAN SH G(简曙光),CHEN Q(陈 权),etal.Vascular plant diversity of the Paracel Islands,China[J].BiodiversityScience(生物多样性),2013,21(3):364-374(in Chinese).
[16] 吴征镒,孙 航,周浙昆,等.中国种子植物区系地理[M].北京:科学出版社,2011.
[17] WEI Z X(魏宗贤),SONG M ZH(宋满珍),NIU Y L(牛艳丽),etal.Flora and life-form of wild vine resources in the Lushan Mountain[J].JournalofZhejiangA&FUniversity(浙江农林大学学报),2013,30(4):505-510(in Chinese).
[18] CHEN SH B(陈圣宾),OUYANG ZH Y(欧阳志云),FANG Y(方 瑜),etal.Geographic patterns of endemic seed plant genera diversity in China[J].BiodiversityScience(生物多样性),2011,19(04):414-423(in Chinese).
[19] QIAN H,WANG S,HE J SH,etal.Phytogeographical analysis of seed plant genera in China[J].AnnalsofBotany,2006,98(5):1 073-1 084.
