高寒草甸植物碳氮组成及其稳定同位素特征
2015-07-04全小龙乔有明段中华梁海东
全小龙,乔有明,段中华,梁海东
(青海大学 生态环境工程学院,西宁810016)
碳(C)、氮(N)是植物必需的养分元素,在植物各项生命活动中发挥着重要作用,植物碳、氮也是土壤有机碳的主要来源,两者在生态系统中的储量、循环与草甸生态系统功能,以及系统的维持、发展和稳定有着密不可分的关系[1-2]。因此,维护生态系统中碳氮平衡对于整个高原生态环境具有十分重要的作用。
黄河源区高寒草甸是青藏高原的典型植被类型之一,在人类活动直接影响或同气候环境协同作用影响下,用于冬春草场的高寒草甸发生了显著的逆行演替,种类组成和群落结的变化以及伴随的土地退化造成植物和土壤碳、氮流失,植物碳的变化进一步影响了土壤有机碳储量。由于这一地区植被组成和土壤有机碳的空间变异大,不同学者对同一问题的研究常常因为地点的差异会得到不一致的结果,而且以往的研究绝大多数采用的是传统的生态学和土壤学方法。长期以来,利用δ13C 值 与δ15N 值 重建古气候、古环境和古生态过程,Robertson等[3]通过分析树轮不同组分的δ13C 值,发现其木质素和纤维素中δ13C 的高低变化同生长季的降水,温度和湿度等综合因素的变化相关,可作为过去气候的测定指标。沈亚婷等[4]对云南省曲靖地区发生植被演替的山地研究显示,δ13C 在C3植被的短期演替过程中具有很好的辨识力。近年来,利用植物稳定同位素技术研究植物光合型及养分、水分利用等受到了极大的重视,在生态学领域得到了广泛的应用[5-7]。但是国内植物稳定同位素研究起步较晚,且集中在森林和农田生态系统中,而对占陆地总面积约1/3的草原生态系统研究较少[8-9]。特别是缺乏对不同植物碳、氮含量以及稳定氮同位素的研究。因此利用稳定同位素的原位标记特性及其可以在长时间跨度和大空间范围检测的优势,分析高寒草甸植物碳氮组成和稳定同位素特征对研究植被演替和土壤有机碳有潜在的意义。
本研究以黄河源区典型高寒草甸为例,对区内主要植物、退化草甸(未退化、轻度退化和严重退化)以及人工改良高寒草地进行碳、氮组成及其稳定同位素特征进行分析,判断植物光合类型,探讨稳定碳氮同位素丰富度对草地植被演替的响应及预判草地的退化趋势。
1 研究区概况
黄河源区处于青藏高原腹地,介于33°27′~35°11′N,98°06′~100°13′E 之间,海拔介于3 730~4 375m。气候寒冷,年平均温度-4 ℃,年平均降水309mm,年日照时数2 400~2 800h,太阳总辐射量达6 500 MJ/m2[10-11]。植被类型以高山嵩草草甸为主,土壤类型为高山草甸土,根据马玉寿等[12]对该地区退化高寒草地的等级划分,并结合张金屯[13]对草地退化程度的划分标准确定样地类型。未退化草甸海拔4 048~4 072m,平均4 060m,以莎草科、禾本科为绝对优势物种,杂草较少、物种分布均匀,总盖度达90%以上,基本无秃斑地;轻度退化草甸海拔3 951~4 047m,平均3 991m,以禾草为优势种,嵩草属植物较多,呈密丛状,物种分布不均匀,总盖度达80%~90%,草地秃斑地占15%~20%;严重退化草甸海拔3 741~4 070 m,平均3 951m,以阔叶杂类草为主,禾草为次,豆科、菊科和莎草科植物偶见,毒草比例相对较大,总盖度为在80%以下,秃斑地面积占30%~50%左右;人工草地海拔3 736~4 104m,平均3 928m,以垂穗披碱草(Elymusnutans)为主,混播有中华羊茅(Festuca sinensis)和冷地早熟禾(Poacrymophila,少量的杂类草,无秃斑地。
2 样品采集及分析
58种主要植物样品采自于2012年和2013年植物生长茂盛季节(7~8月),所采植物样品均为正在生长的植物,每一种植物样品均由6~8株不同的个体混合而成。退化草甸及人工改良草地样品采于2013年8月初,采样涉及青海省果洛藏族自治州玛沁县大武镇、格多牧委会和达日县窝赛乡。在样点内随机选取1m×1m 样方进行采样,将框内地上植物(不含根系)全部剪下装袋作为一个样品,共32个样点,其中包括未退化草甸(植被盖度≥90%)5个,轻度退化草甸(80%≤植被盖度<90%)5个,严重退化草甸(植被盖度<80%,包括中度退化和重度退化草甸)13个,人工改良草地9个。实验室内将样品洗净,70 ℃恒温箱中烘干48h至恒重,再粉碎过100目筛筛制成供试样品。
样品测定在中国农业科学院农业环境与可持续发展研究所农业环境稳定同位素实验室完成。通过自动进样器进入元素分析仪(vario PYRO cube,德国Elementar公司)经燃烧与还原转化为纯净的CO2和N2气体,CO2再经过稀释器稀释,最后进入稳定同位素质谱仪(IsoPrime100,英国Isoprime公司)进行检测。每个样品重复测定3次。运用Excel 2010进行数据分析和制图。
3 结果与分析
3.1 高寒草甸主要植物碳氮组成及其稳定同位素特征
3.1.1 高寒草甸主要植物碳氮组成 由表1可知,本研究中采集的58种高寒草甸主要植物分属于22科、42属。其中菊科植物最多为13种,莎草科和禾本科各有6种,龙胆科、唇形科、毛茛科和蔷薇科均为4种,玄参科3种,豆科2种,其余科属均为1种植物。
本区植物碳元素含量在28.64%~51.55%变化(表1),平均值为43.12%,其中唇形科密花香薷(Elsholtziadensa)含量最低,而同为唇形科的黄花粘毛鼠尾草(Salviaroborowskii)含量最高,且超过2/3的植物集中于40%~50%;氮元素含量介于0.89%~4.04%,平均值为2.04%,其中禾本科垂穗披碱草(Elymusnutans)含量最低,而胡颓子科西藏沙棘(Hippophaethibetana)含量最高,有37 种植物氮含量处于1.5%~2.5%。
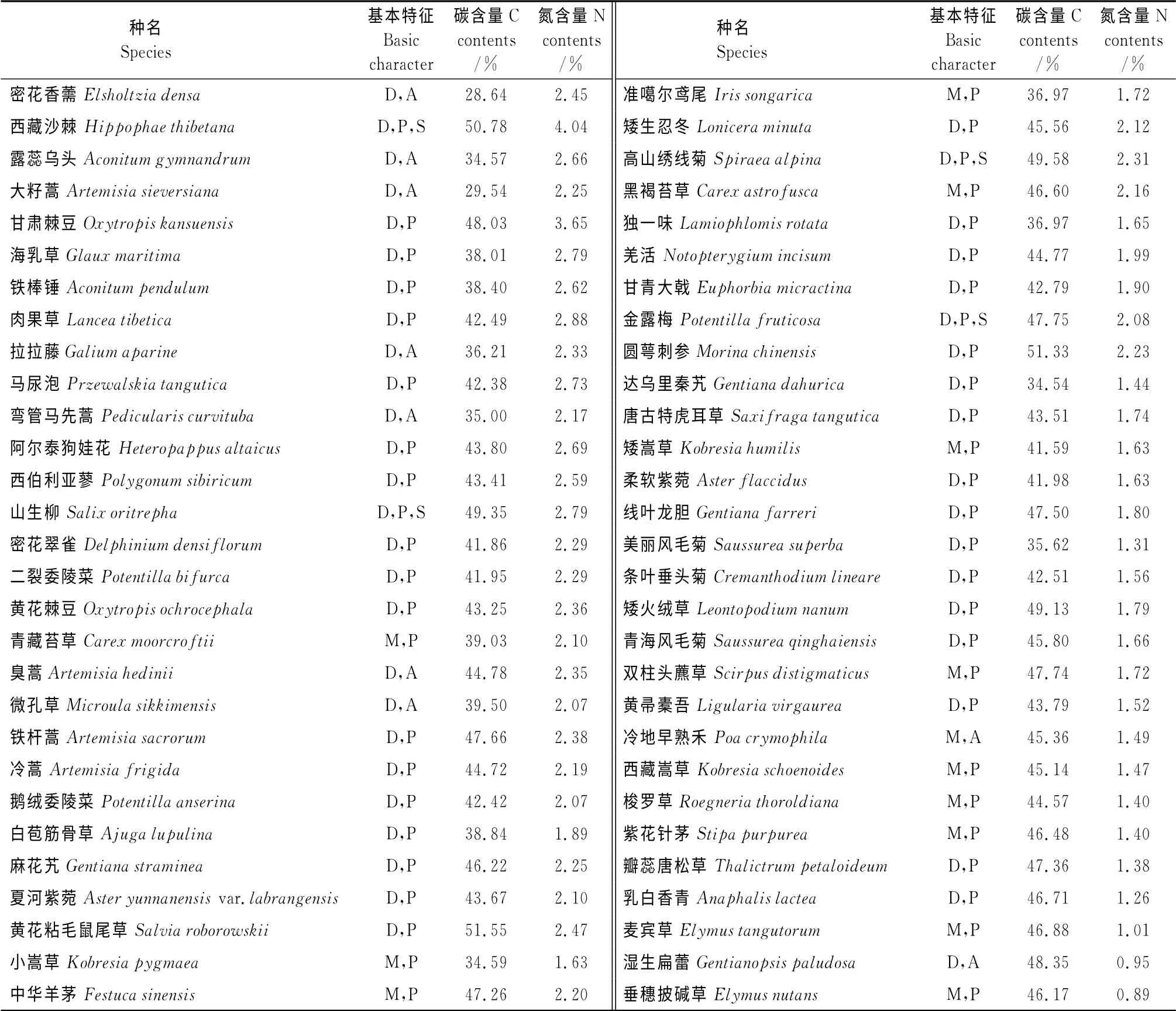
表1 植物碳氮元素组成Table1 Carbon and nitrogen composition of plants
3.1.2 高寒草甸主要植物稳定同位素特征 表2显示,本区内植物测定植物的稳定碳同位素比值变化范围介于-29.50‰ ~-24.69‰,平均为-26.98‰。稳定氮同位素比值介于-4.57‰~8.32‰,平均值为0.47‰。有80%的植物δ15N 值集中于-1.81‰~2.89‰。
3.2 不同退化高寒草甸植物碳氮组成及其稳定同位素特征
3.2.1 不同退化高寒草甸植物碳氮组成 由表3可以看出,本区未退化、轻度退化、严重退化草甸和人工草地的C 元素平均含量依次为43.18%、42.18%、39.68%和45.54%,N 元素平均含量依次为2.30%、2.13%、2.10%和2.28%(表3)。未退化草甸和人工草甸碳氮含量相对偏高,而严重退化草甸碳氮含量在4种类型中均最低。植物碳氮含量受草地退化程度影响,而当人工修复后植物碳氮含量有所增加。
3.2.2 不同退化高寒草甸植物稳定碳氮同位素特征 由表3可知,不同退化高寒草甸δ13C 值主要集中在-27.00‰至-26.00‰间,且严重退化草甸中δ13C值最低,其次为轻度退化草甸与人工改良草地较为相近,而未退化草甸δ13C 值处于最高水平。表明植物δ13C值随草地退化程度加剧而逐渐降低。表3表明,不同退化高寒草甸植物δ15N 值主要集中在-1.00‰~1.00‰。δ15N 值大小顺序依次为轻度退化草甸(2.76‰)>人工草地(0.32‰)>严重退化草甸(0.26‰)>未退化草甸(-0.63‰)。表明δ15N值没有随草地退化程度加剧而逐渐降低的变化趋势。不同退化高寒草甸δ13C 与δ15N的含量关系分布如图1所示。
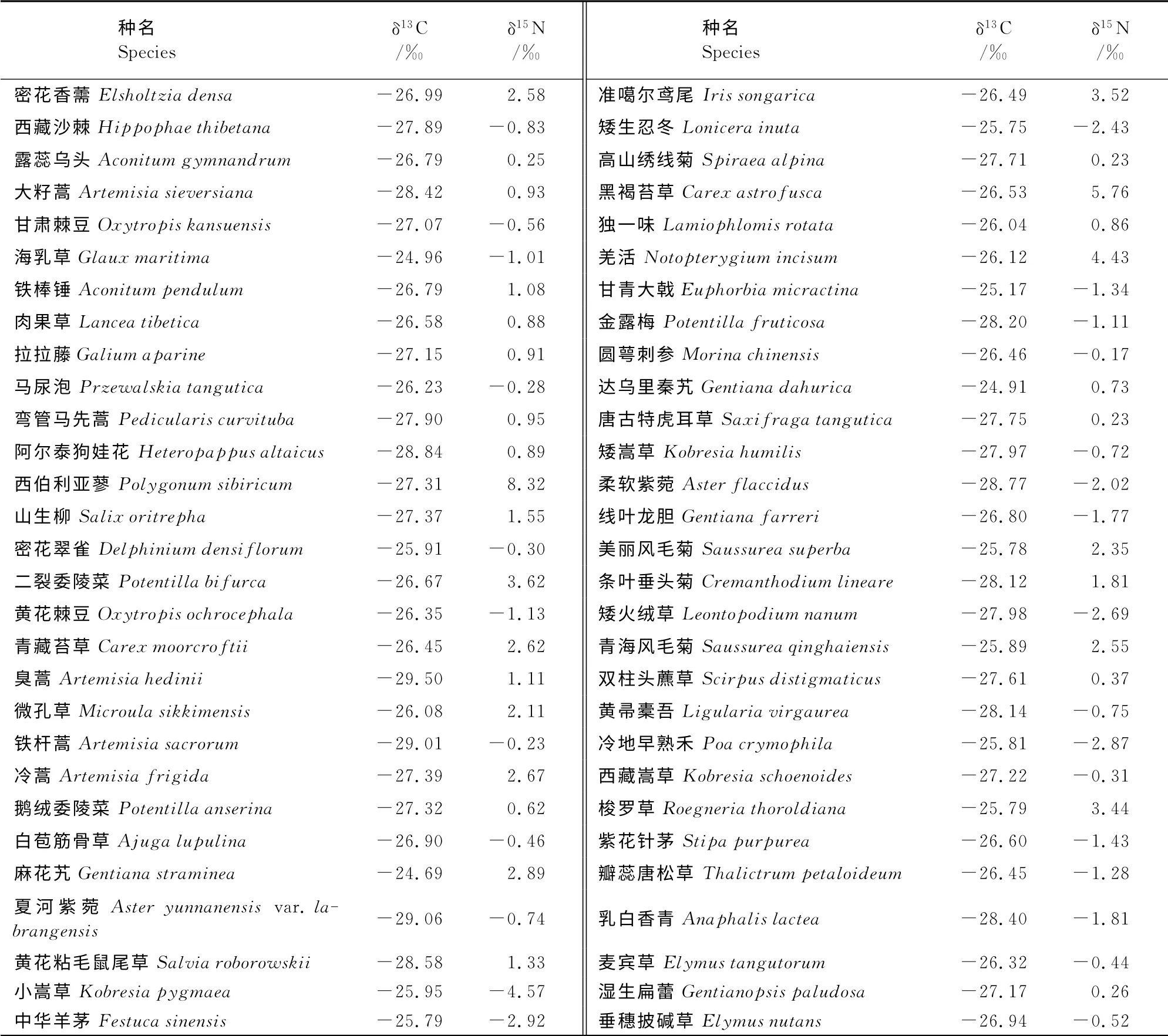
表2 植物稳定碳氮同位素组成Table2 Stable carbon and nitrogen isotope composition of plants
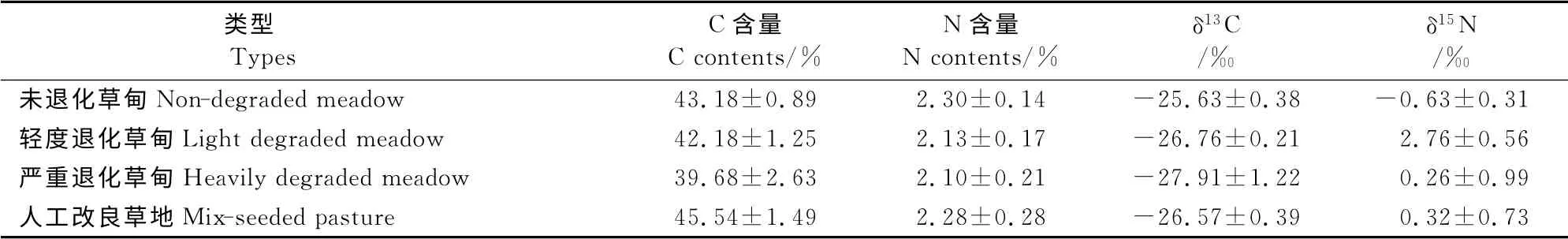
表3 不同退化高寒草甸及人工草地植物碳氮组成及稳定碳氮同位素Table3 Carbon,nitrogen,stable carbon and nitrogen isotope composition in different meadows and seeded pasture

图1 不同退化程度草甸δ13 C和δ15 N 关系Fig.1 Relationship betweenδ13 C andδ15 N in different meadows and seeded pasture
4 讨 论
不同植物随着海拔和个体的差异而碳氮含量不同[14-15]。李英年等[16]测定了祁连山冷龙岭南坡11种移地植物的碳、氮含量,其中麻花艽、美丽风毛菊、垂穗披碱草、鹅绒萎陵菜、肉果草、金露梅和矮嵩草为本次涉及植物。相较而言本区内垂穗披碱草碳含量和肉果草氮含量较之偏高,金露梅碳含量相对接近,除此之外碳氮含量均低于祁连山冷龙岭植物。一般认为,随着海拔的提升,植物叶片氮含量增加,碳含量降低。而本研究测定了整个地上植物的碳氮含量,这可能是造成碳氮含量存在差异的主要原因。
Wang等[17]比较了混播、单播、自然恢复、轻度及重度退化草甸植物的碳氮浓度,认为不同管理措施下禾草类、杂类草和莎草类植物的碳浓度没有显著差异,数据的总趋势是未退化草甸(或轻度退化草甸)植物碳浓度高于其他处理,而氮浓度有显著不同,退化草甸植物的氮浓度最高。王启基等[18]对高山嵩草草甸轻度退化草甸和重度退化草甸植物的碳氮浓度表明,重度退化草甸的植物碳浓度低于轻度退化草甸的,而两者的全氮浓度没有显著差别。本结果显示,人工草地植物碳、氮浓度均较高。未退化草甸或轻度退化草甸植物碳浓度有高于重度退化草甸的趋势,即草地退化引起养分供应或植物种类的变化,进而引起植物碳浓度的降低,本研究结果与前人研究结果类似。
根据Schnyder等[19]和Zhu等[20]统计显示全球C3植物的δ13C值分布在-34.00‰~-22.00‰,平均值-27.00‰;C4植物为-15.00‰~-7.00‰,平均值为-12.00‰。而本区内植物介于-29.50‰~-24.69‰,可以判断植物均属于C3植物,而未发现任何C4植物。Tieszen等[21]认为海拔高于3 000 m 就没有C4植物分布。王国安[22]对甘肃省肃南县(海拔>2 700 m)的考察发现该地年均温度低于3℃,在草地中根本没有C4植物。但旺罗等[23]在青藏高原低纬度干旱地区发现了藜科2种和禾本科6种C4植物,认为C4植物可出现在海拔4 000m 以上,甚至可达4 520m。李明财等[24]发布的青海高原高寒地区C4植物名录指出青海高原高寒地区有9科32属,共52种C4植物。由此可见青藏高原是存在C4植物的,但本研究中未涉及上述52种植物,也未能发现新C4植物,可以确定本研究区不适宜C4植物。与本研究相比,旺罗等研究区纬度较低,李明财等研究区海拔偏低。因此认为温度可能是制约本区C4植物分布的关键因素。
李嘉竹等[25]对贡嘎山的研究结果表明,δ13C 值随海拔高度的增加而变重,每千米变化幅度为1.3‰。本研究取样地海拔3 736~4 104m,海拔差约400m,但采样地区均属于典型的高寒草甸分布区,不同样地的土壤类型、降雨量,甚至地形地貌比较相似,所以差异不大,对结果的影响较小。未退化草甸(-25.63‰)同Körner等[26]在全球一些高海拔地区(2 500~5 600m)调查的草本植物叶片δ13C值(平均值为-25.65‰)结果相似,而Körner的调查区植被属于原始未退化草甸,因此,该δ13C 值可作为未退化草甸的参考。 轻度退化草甸(-26.76‰)和陈拓等[27]在青藏高原北部调查植物叶片δ13C 值结果(平均值-26.89‰)较为接近,而陈拓等正是对处于退化时期的草地进行调查分析。δ13C值的递减变化同草地退化程度是一致的,草地退化植物数量减少,光合量降低,δ13C值偏低。
大气中的N2其δ15N 值接近0,而土壤中δ15N值在-6‰~16‰[28]。因此,主要从土壤中吸收氮素的植物其δ15N 丰富度应高于通过固氮作用从大气中获得氮素的植物。未退化草甸植物种类多,植物吸收的氮素主要来源于生物固氮,因此δ15N 值较低。轻度退化草甸δ15N 值偏高,可能是植物吸取了深层土壤中的氮素[29]。人工草地是对严重退化草甸的改良,因而严重退化草甸和人工草地δ15N 平均值相对接近,这与土壤类型以及土壤中的微生物活动是密不可分的。
Martinelli等[30]对来自欧美一些地区的植物平均δ15N 组成与温度关系的调查和刘晓宏等[31]对东非裂谷带植物δ15N 组成与环境的关系研究都报道了植物δ15N 值随温度增加而变大的现象。本研究的3个采样地点间温度没有明显的差异,温度不可能是造成本研究所涉及植物的δ15N 差异的主要因素,不同植物种之间δ15N 的差异所反映出来的是种的特性,表明不同植物对含δ15N 氮素的固定和吸收能力是有差异的。
[1] DING X G(丁喜桂),YE S Y(叶思源),WANG J S(王吉松).Stable carbon and nitrogen isotopes in the Yellow River delta wetland[J].MarineGeologyFrontiers(海洋地质前沿),2011,27(2):66-71(in Chinese).
[2] WANG J L(王建林),ZHONG ZH M(钟志明),WANG ZH H(王忠红),etal.Distribution characteristics and its influence factors of vegetation C/N value of alpine steppe ecosystem in Tibetan Plateau[J].JournalofPlantResourcesandEnvironment(植物资源与环境学报),2013,22(4):20-28(in Chinese).
[3] ROBERTSON I,LOARDER N J,MCCARROLL D,etal.δ13C of tree-ring lignin as an indirect measure of climate change[J].WaterAir&SoilPollutionFocus,2004,4(14):531-544.
[4] SHEN Y T(沈亚婷),LU G H(路国慧),HU J D(胡俊栋),etal.13C Distribution characteristics in soil profiles with the impacts of shortterm trees,shrubs and grass replacement[J].ProgressinGeography(地理科学进展),2012,31(11):1 460-1 466(in Chinese).
[5] CERNUSAK L A,UBIERNA N,WINTER K,etal.Environmental and physiological determinants of carbon isotope discrimination in terrestrial plants[J].NewPhytologist,2013,200(4):950-965.
[6] DI MATTEO G,PERINI L,ATZORI P,etal.Changes in foliar carbon isotope composition and seasonal stomatal conductance reveal adaptive traits in Mediterranean coppices affected by drought[J].JournalofForestryResearch,2014,25(4):839-845.
[7] HERRERO A,CASTRO J,ZAMORA R,etal.Growth and stable isotope signals associated with drought-related mortality in saplings of two coexisting pine species[J].Oecologia,2013,173(4):1 613-1 624.
[8] LUO G Q(罗光强),GENG Y B(耿元波),YUAN G F(袁国富).Application and prospect of carbon isotope in the study of carbon cycle in grassland ecosystem[J].ProgressinGeography(地理科学进展),2009,28(3):441-448(in Chinese).
[9] MA Y(马 晔),LIU J CH(刘锦春).Applications ofδ13C in plant ecological research[J].ActaBotanicaBoreali-OccidentaliaSinica(西北植物学报),2013,33(7):1 492-1 500(in Chinese).
[10] DING Y J(丁永建),YANG J P(杨建平),LIU SH Y(刘时银),etal.Exploration of eco-environment range in the Source Regions of the Yangtze and Yellow Rivers[J].ActaGeographicaSinica(地理学报),2003,58(4):519-526(in Chinese).
[11] PAN J H(潘竟虎),WANG J(王 建),WANG J H(王建华).Dynamic change of frigid wetlands in the Source Region of the Yangtze and Yellow Rivers[J].WetlandScience(湿地科学),2007,5(4):298-304(in Chinese).
[12] MA Y SH(马玉寿),LANG B N(郎百宁),LI Q Y(李青云),etal.Study on rehabilitating and rebuilding technologies for degenerated alpine meadow in the Changjiang and Yellow River source region[J].PrataculturalScience(草业科学),2002,19(9):1-5(in Chinese).
[13] 张金屯.应用生态学[M].北京:科学出版社,2003:173-174.
[14] ZHOU X(周 欣),ZUO X A(左小安),ZHAO X Y(赵学勇),etal.Effect of change in semiarid sand dune habitat on aboveground plant biomass,carbon and nitrogen[J].ActaPratacultureSinica(草业学报),2014,23(6):36-44(in Chinese).
[15] YIN X R(银晓瑞),LIANG C ZH(梁存柱),WANG L X(王立新),etal.Ecological stoichiometry of plant nutrients at different restoration succession stages in typical steppe of Inner Mongolia,China[J].ChineseJournalofPlantEcology(植物生态学报),2010,34(1):39-47(in Chinese).
[16] LI Y N(李英年),ZHAO X Q(赵新全),ZHANG F W(张法伟),etal.Response of carbon and nitrogen of alpine plant leaves to transloca-tion experiment along the elevation gradient in the Southern Slope of Lenglongling,Qilian Mountains[J].ActaBotanicaBoreali-OccidentaliaSinica(西北植物学报),2011,31(4):788-794(in Chinese).
[17] WANG W Y,WANG Q J,WANG CH Y,etal.The effect of land management on carbon and nitrogen status in plants and soils of alpine meadows on the Tibetan Plateau[J].LandDegradation&Development,2005,16:405-415.
[18] WANG Q J(王启基),LI SH X(李世雄),WANG W Y(王文颖),etal.The despondences of carbon and nitrogrn reserves in plants and soils to vegetations cover change onKobresiapygmaemeadow of Yellow and Yangtze River source region[J].ActaEcologicaSinica(生态学报),2008,28(3):885-894(in Chinese).
[19] SCHNYDER H,LATTANZI F A.Partitioning respiration of C3-C4mixed communities using the natural abundance13C approach-testing assumptions in a controlled environment[J].PlantBiology,2005,7(6):592-600.
[20] BIAO ZHU,WEIXIN CHENG.13C isotope fractionation during rhizosphere respiration of C3and C4plants[J].PlantandSoil,2011,342(1-2):277-287.
[21] TIESZEN L L,SENYIMBA M M,IMBAMBA S K,etal.The distribution of C3and C4grasses and carbon isotope discrimination along an altitudinal and moisture gradient in Kenya[J].Oecologia,1979,37(3):337-350.
[22] WANG G A(王国安).Application of stable carbon isotope for paleoenvironmental research[J].QuaternarySciences(第四纪 研究),2003,23(5):471-484(in Chinese).
[23] WANG L(旺 罗),LV H Y(吕厚远),WU N Q(吴乃琴),etal.Discovery of C4species in Qinghai-Tibet Plateau at high altitudes[J].ChineseScienceBulletin(科学通报),2004,49(13):1 290-1 293(in Chinese).
[24] LI M C(李明财),YI X F(易现峰),ZHANG X A(张晓爱),etal.The list of C4plants in alpine locality of Qinghai Plateau[J].ActaBotanicaBoreali-OccidentaliaSinica(西北植物学报),2005,25(5):1 046-1 050(in Chinese).
[25] LI J ZH(李嘉竹),WANG G A(王国安),LIU X ZH(刘贤赵),etal.Variations in carbon isotope ratios of C3plants and distribution of C4plants along an altitudinal transect on the easternslope of Mount Gongga[J].ScienceinChinaSeriesD(中国科学),2009,(10):1 387-1 396(in Chinese).
[27] CHEN T(陈 拓),YANG M X(杨梅学),FENG H Y(冯虎元),etal.Spatial distribution of stable carbon isotope compositions of plant leaves in the North of the Tibetan Plateau[J].JournalofGlaciologyandGeocryology(冰川冻土),2003,25(1):83-87(in Chinese).
[28] SHEARER G,KOHL D H.N2-fixation in field settings:estimations based on naturalδ15N abundance[J].FunctionalPlantBiology,1986,13(6):699-756.
[29] ZHAO H L(赵哈林),LI Y Q(李玉强),ZHOU R L(周瑞莲).Effects of desertification on C and N storages in grassland ecosystem on Horqin sandy land[J].ChineseJournalofAppliedEcology(应用生态学报),2007,18(11):2 412-2 417(in Chinese).
[30] MARTINELLI L A,PICCOLO M C,TOWNSEND A R,etal.Nitrogen stable isotopic composition of leaves and soil:tropical versus temperate forests[M]//New Perspectives on Nitrogen Cycling in the Temperate and Tropical Americas.Springer Netherlands,1999:45-65.
[31] LIU X H(刘晓宏),ZHAO L J(赵良菊),GASAW M,etal.Foliarδ13C andδ15N values of C3plants in the Ethiopia Rift Valley and their environmental controls[J].ChineseScienceBulletin(科学通报),2007,52(2):199-206(in Chinese).
