线叶嵩草草地群落构成及种间关联分析
2015-07-04康海军孔杨云德科加徐成体
王 伟,郭 倩,康海军,王 莹,孔杨云,张 明,德科加,徐成体*
(1 青海省畜牧兽医科学院,西宁810003;2 兰州大学 草地农业科技学院,兰州730020;3 青海省草原总站,西宁810008)
植物种间关联通常是由于群落生境差异影响物种分布而引起的,是群落基本特征之一,反映植物物种空间上相互关系、相互影响[1]。根据群落内种对间对生境的选择性及种对间的相互吸引和排斥,种间关系一般可分为正相关、负相关和不相关[2]。种对间表现为正相关是由于物种具有相近的生物学特性和对环境具有相似的生态适应性以及一个种群的存在可以为另一个种群的生存提供条件;而种对间表现为负相关是由于物种的生物学特性不同和对环境喜好的不同以及相互竞争[3]。植物种间关联既是一种通过物种存在与否的二元数据进行的定性分析,同时也是通过物种的数量数据进行的定量分析[4]。研究不同群落的植物种间关系,能够详细反映物种在空间上的分布以及群落物种间相互依存、相互制约的复杂联系,对揭示群落组成结构及功能、动态和演替等机制具有重要意义[5-7]。
高寒嵩草草甸约占青藏高原总面积的35% ,主要作为冬春放牧草场利用,在高原草地畜牧业中占有十分重要的地位[8]。嵩草属植物多为高寒草甸的建群种或优势种[9],线叶嵩草一般分布在土壤水分适中的滩地、湖盆、山地阳坡和阴坡,形成典型草甸[10]。国内在不同植被类型的种间关联方面的研究 很 多[3,11-13],高寒草甸的研究工作主要集中于高寒草甸的退化[8]、群落生产力、生物多样性和群落结构以及围封与放牧条件下群落的动态变化[14-15]等方面。一些学者对高寒草甸植物种间联结做了研究,柳剑丽等[16]发现锡林郭勒典型草原羊草(Leymus chinensis)单优群落植物种间总体关联性近似无关联,群落处于相对稳定阶段,而处于演替过渡状态的克氏针茅(Stipakrylovii))+羊草+冷蒿(Artemisiafrigida)群落、冷蒿+黄蒿(Artemisiascoparia)+羊草群落、羊草+糙隐子草(Cleistogenes quarrosa)+星毛委陵菜(Potentillaacaulis)群落总体为负关联,群落处于相对不稳定阶段;王卫等[17]发现那拉提草原禾草+杂类草群落种间关系表现为正联结,群落处于稳定期;房飞等[13]报道,巴音布鲁克高寒草原羊茅(Festucaovina)+紫花针茅(Stipa purpurea)群落种间关系为正相关,群落处于相对稳定阶段。但有关高寒草甸嵩草群落植物种间关联的研究相对缺乏。
植被稳定性是植物群落结构与功能的一个综合特征。稳定性的维持受群落内的生物多样性、种间关系等一系列生物学、生态学过程的控制[18],分析高寒草甸植物种间联结是研究草地恢复程度及稳定性的重要方法之一。本研究采用方差比例法、χ2检验、Jaccard指数以及Spearman秩相关,对青藏高原2个线叶嵩草群落植物种构成及主要物种种间关系进行分析,对研究草地群落演替、草地经营管理以及植被恢复和重建具有重要意义。
1 材料和方法
1.1 研究区概况
研究区位于青海省河南县境内,地理坐标N 34°04′52″~34°55′36″,E 100°53′25″~102°15′27″。河南县气候为高原大陆性气候,属高原亚寒带湿润气候区。由于海拔较高,地势复杂和受季风影响,高原大陆性气候特点比较明显。每年5~10月份温暖、多雨,11月至次年4月份寒冷、干燥、多大风天气。春秋时日短,四季不分明,无绝对无霜期。年均气温在9.2 ℃~14.6 ℃,年降水量597.1~615.5 mm。草甸类型以线叶嵩草草甸为主,主要植物种有线叶嵩草(Kobresiacapillifolia)、垂穗披碱草(Elymusnutans)、艹洽草(Koeleriacristata)、早熟禾(Poaannua)、委陵菜(Potentillasp.)、蒲公英(Taraxacummongolicum)、毛茛(Ranunculusjaponicus)、黄帚橐吾(Ligulariavirgaurea)等。
1.2 样地设置
在实地调查基础上,依据草地群落类型不同,于2009年7~8月,在青海省河南县的宁木特乡和优干宁镇的草地利用率约70%~85%的冬春线叶嵩草草地上,设置线叶嵩草群落样地6个,退化的线叶嵩草群落样地5个。各样地大小为0.5~1.0hm2,共设置11个样地。
1.3 测定指标和方法
于2010~2013年8月中、下旬,在各样地内,分别随机设置0.5m×0.5m 的样方5个,调查各样方内植物种的高度、盖度和频度。
1.3.1 重要值 植物种重要值(Important value,IV)的计算公式为:IV=(RC+RF+RH)/3
式中,RC、RF和RH分别表示植物种的相对盖度、相对频度和相对高度。取每个群落中重要值前15的植物种进行研究。
1.3.2 总体关联性 采用Schluter提出的方差比率(VR,Variance ratio)法[19],其公式为:

式中,S为总物种数,N为总样方数,Tj为样方j内出现的所研究物种的总数,ni为物种i出现的样方数,t为全部样方中种的平均数,ST2为所有样方物种数的方差,δT2为所有物种出现频度的方差。VR值为群落内植物种间的总体联结指数。VR=1,接受零假设即所有种间无关联;VR>1或VR<1否定零假设,接受备择假设即种间表现出正关联或负关联。为了检验VR偏离1是否显著,引入新的统计量W=VR×N。W服从χ2分布(df=N-1)。若W<(N)或W>(N),则物种间总体关联显著(P<0.05);反之若χ20.95(N)<W<(N),则物种间总体关联不显著。
1.3.3 种对间联结性检验 一般采用χ2检验对植物种对进行定性研究,χ2检验以2×2列联表为基础计算[20]。在种间联结性的研究中,由于数据的不连续性,需采用Yates校正公式[4]。其公式为:

式中,N为总样方数,a为所有样方中种A和种B共同出现的样方数,b、c分别为所有样方中种A或种B单独出现的样方数,d为种A和种B都不出现的样方数。当时,说明种对间联结性极显著;当时,说 明种对间联结性显著;当时,则 可认为2个种独立分布,即无联结。当ad-bc>0时为正联结,当ad-bc<0时为负联结。
1.3.4 植物种间关联程度测定 Jaccard指数(JI)反映的是物种共同出现的百分率。一般来说,χ2检验只有和关联度指数结合分析才能较准确反映种间关系[21]。采用JI指数测定物种间关联程度,它们仍以2×2列关联表为基础计算[4,22]。其公式为:
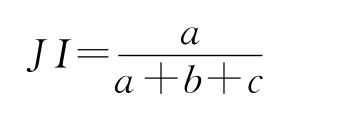
JI指数满足当a=0时,取值为0,表示种A和种B不同时出现在同一样方内,即种A和种B完全独立或相异;当a=S(样方数)时,b=c=0,取值为1,表示物种A、B同时出现在所有样方中,即种A与种B绝对关联;当c=0时,表示完全关联,即种B依赖于种A,种B随种A出现而出现,而种A可以单独出现。0.8<JI≤1 为强联结,0.6<JI≤0.8为次强联结,0.4<JI≤0.6为次弱联结,0.2<JI≤0.4为弱联结,0≤JI≤0.2为无联结[23]。
1.3.5 Spearman秩相关分析 Spearman秩相关系数属于非参数性检验,由植物种重要值计算。其公式为:
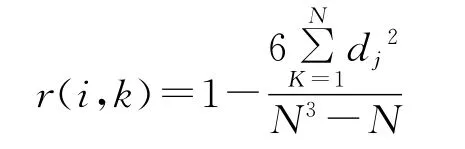
式中,r(i,k)为物种i与物种k的相关系数,N为样方数,dj=(xij-xkj),xij、xkj分别为物种i和物种k在样方j中的秩[4]。
1.4 数据分析
用Excel进行数据处理,用Matlab 程序进行2×2列联表及秩相关系数计算。
2 结果与分析
2.1 群落主要植物种重要值
草地群落主要植物种重要值显示,线叶嵩草群落以线叶嵩草为优势种,垂穗披碱草为亚优势种,洽草、早熟禾、矮嵩草、蒲公英、秦艽、毛茛等为主要伴生种;而退化的线叶嵩草群落以垂穗披碱草为优势种,洽草、珠芽蓼、委陵菜、线叶嵩草、蒲公英、早熟禾等为主要伴生种(表1)。比较2个线叶嵩草群落的物种构成,15种植物种有12种相同,仅3 种不同,可得出2个群落物种组成相似,差异较小。
2.2 2个线叶嵩草群落总体关联性
草地群落15个优势种间总体关联性分析可知,2个群落的VR值均小于1且W不在<W<范围内(表2)。说明,2个线叶嵩草群落中15个优势种间的总体关联性表现为显著负相关,群落整体关系比较紧密。
2.3 2个线叶嵩草群落物种间关联性
χ2检验统计结果中,线叶嵩草群落正联结的种对数为51对,负联结的种对数为54,分别占总种对数的48.6%和51.4%;退化的线叶嵩草群落正联结的种对数为50对,负联结的种对数为55,分别占总种对数的47.6%和52.4%。2个线叶嵩草群落中正联结与负联结的种对数相近,正联结种对数略少于负联结种对数。线叶嵩草群落中极显著相关的种对数分别为29对,其中正相关15对,负相关14对,分别占总种对数的14.3%和13.3%;显著相关的种对数为12对,9对呈正相关,3对呈负相关,分别占总种对数的8.6%和2.9%。退化的线叶嵩草群落中极显著相关的种对数分别为25对,其中正相关10对,负相关15对,分别占总种对数的9.5%和14.3%;显著相关的种对数为5对,3对呈正相关,2对呈负相关,分别占总种对数的2.9%和1.9%。线叶嵩草群落和退化的线叶嵩草群落中无关联的种对数分别为64对和75对,分别占总种对数的61.0%和71.4%。χ2检验的结果表明2个线叶嵩草群落物种间联结性弱,独立性强,且群落中正负联结种对数相近。
两种群落的主要种对的JI指数显示,线叶嵩草群落中强联结的种对数为1对,次强联结的种对数为9对,次弱联结的种对数为31对,弱联结的种对数为57对,无联结的种对数为7对,分别占总种对数的1.0%、8.6%、29.5%、54.2%和6.7%。退化的线叶嵩草群落中强联结的种对数为5对,次强联结的种对数为14对,次弱联结的种对数为45对,弱联结的种对数为25对,无联结的种对数为16对,分别占总种对数的15.2%、13.3%、42.9%、23.8%和15.2%(表3)。在正联结中,关联度指数越大,说明种对间关联程度越紧密,反之则说明种对间关联程度越弱;在负联结中,关联度指数越小,说明种对间关联程度越强,反之则说明种对间关联程度越弱。线叶嵩草群落中χ2检验与JI指数在大多种对间表现出一致性,如种对珠芽蓼-秦艽、蒲公英-异针茅、秦艽-美丽风毛菊等χ2检验表现出显著正相关,JI指数均大于0.6;种对洽草-秦艽、蒲公英-黄帚槖吾、矮嵩草-委陵菜、委陵菜-三脉梅花草等χ2检验表现出显著负相关,JI指数均小于0.4。结果表明,线叶嵩草群落和退化的线叶嵩草群落中弱联结及无联结的种对数分别占总种对数的90.4%和81.9%,说明2个线叶嵩草群落中大多植物种种间关联程度较弱。
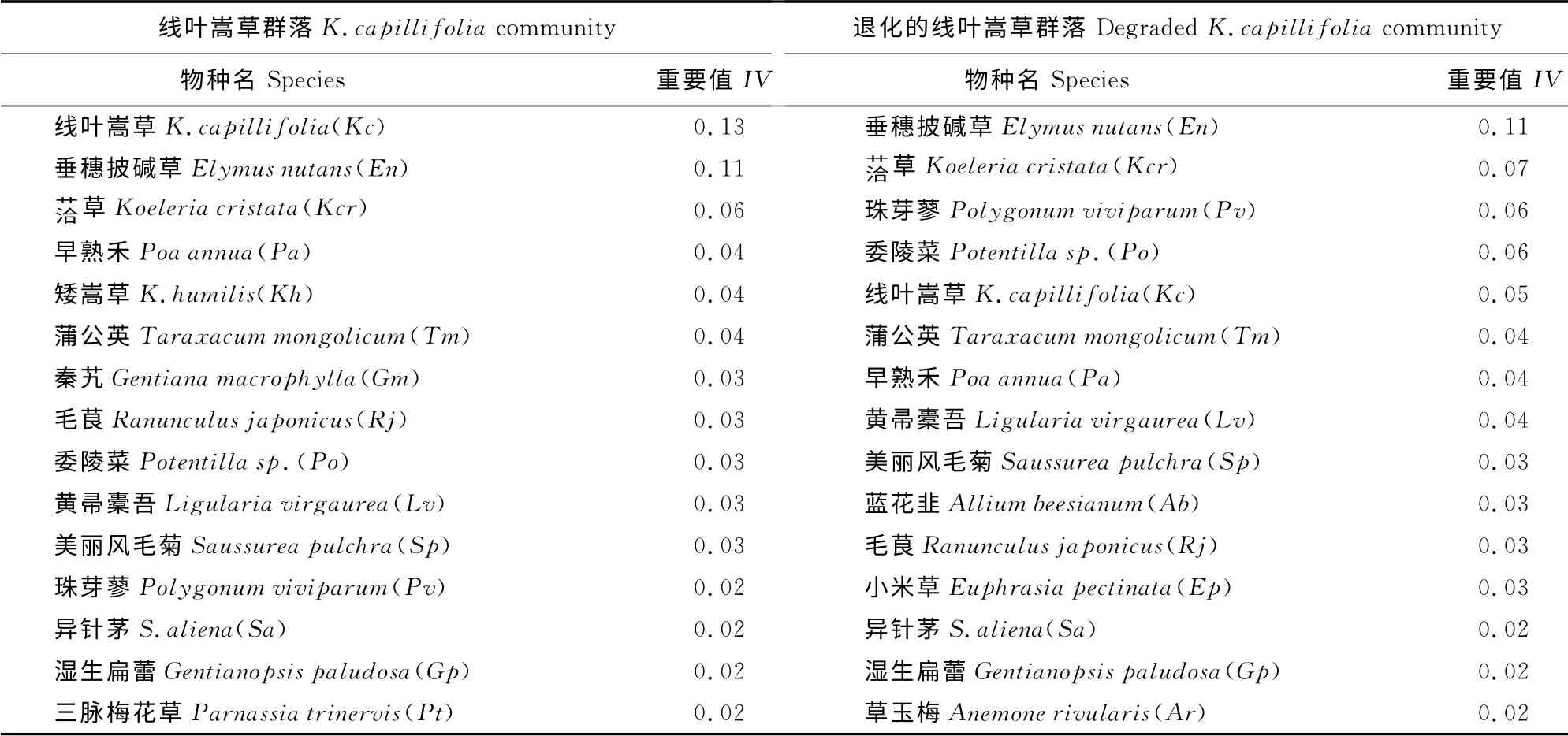
表1 2个线叶嵩草群落中主要植物种重要值的4年均值Table1 The mean of important values in four year about the main plant species in the two K.capillifoliacommunities
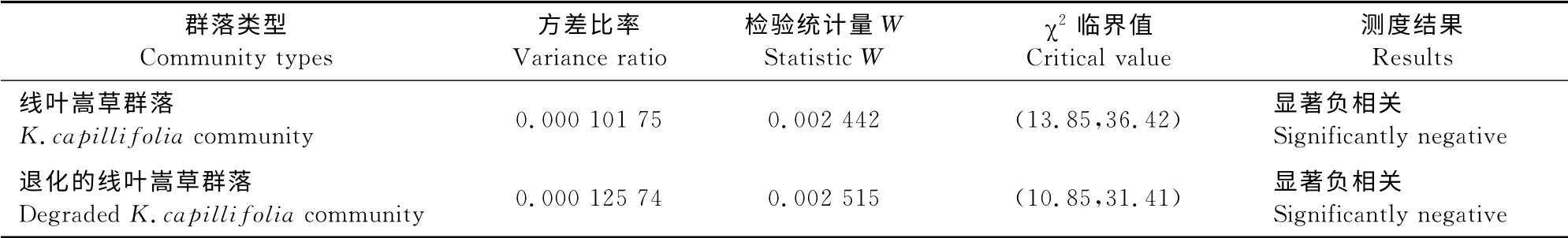
表2 2个线叶嵩草群落主要植物种间的总体关联性Table2 The overall association among main plants distributed in the two K.capillifoliacommunities
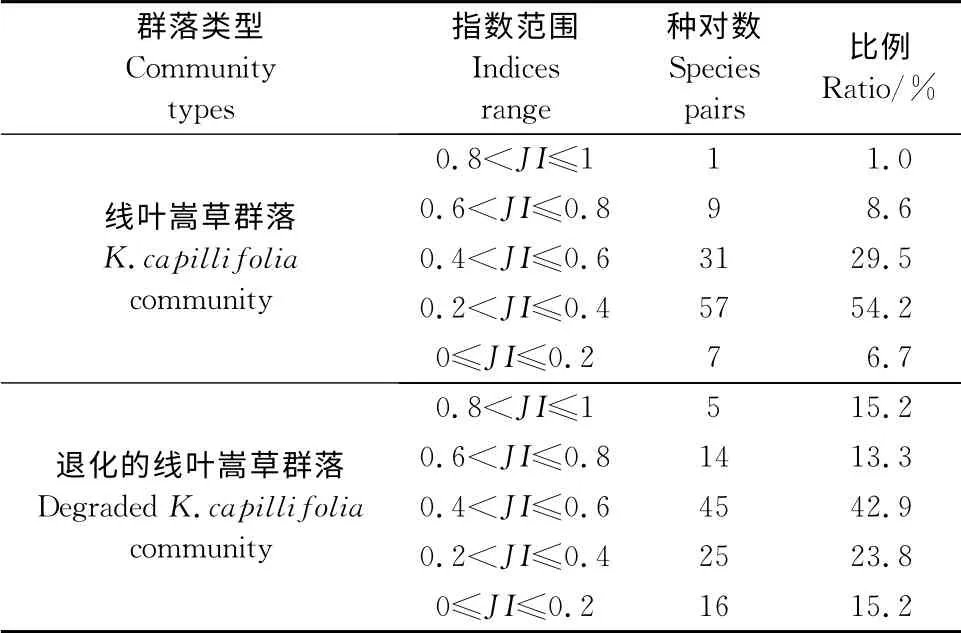
表3 2个线叶嵩草群落的Jaccard指数Table3 The Jaccard indices in the two K.capillifoliacommunities
2.4 2个线叶嵩草群落主要植物种种间Spearman秩相关分析

图1 2个群落主要植物种Spearman秩相关分析半矩阵图Fig.1 Semi-matrix of spearman rank correlation analysis of main species in the two communities
Spearman秩相关分析显示,线叶嵩草群落中,正相关的种对为56对,其中极显著相关的3对,显著相关的9对,不显著相关的44对;负相关的种对为4 9对,显著相关的7对,不显著的4 2对(图1)。退化的线叶嵩草群落中,正相关的种对为60对,极显著的1对,显著的9对,不显著的50对;负相关的种对为45对,极显著的1对,显著的3对,不显著的41对。线叶嵩草群落和退化的线叶嵩草群落中正相关的种对数分别占总种对数的57.1%和53.3%,说明这2个群落中正相关种对数与负相关种对数接近。线叶嵩草群落和退化的线叶嵩草群落中显著关联的种对数分别为19对和14对,分别占植物总种对数的18.1%和13.3%;其中显著(极显著)正相关的种对分别为12对和10对,占总种对数的11.4%和9.5%;显著(极显著)负相关的种对分别为7对和4对,占总种对数的6.7%和3.8%,说明2个群落中大多数植物种对间联结性较弱,种间独立性强,群落处于不稳定的阶段。在线叶嵩草群落中,线叶嵩草与异针茅、垂穗披碱草、矮嵩草、蒲公英等为正联结关系,其中与异针茅的联结最为显著;线叶嵩草与洽草、早熟禾、秦艽、美丽风毛菊等为负联结关系。在退化的线叶嵩草群落中,线叶嵩草与洽草、异针茅、湿生扁蕾、草玉梅、蓝花韭等为正联结关系且与湿生扁蕾的联结性最强;线叶嵩草与蒲公英、毛茛、小米草、垂穗披碱草、珠芽蓼等为负联结关系。由此可以看出,群落处于不同的演替阶段,群落中同一种对的关系存在差异。
3 讨 论
研究区的2个线叶嵩草群落总体上群落植物种间关系均为显著负相关,整体上比较紧密。但χ2检验和Jaccard指数及Spearman秩相关分析结果一致表明,群落内大多物种间联结性较弱,独立性较强;且呈正相关和负相关的种对数在所有种对数中几乎各占一半,可能由于群落内部在长期演替过程中,相互依存与相互排斥作用共存引起。有一部分种群占据了适宜的生态位,彼此适应,和谐共处;而另外一部分种群独立性较强或存在一定竞争[24]。通常,植物的分布范围与气候、土壤、地形、历史及人类活动和植物适应环境属性等有关[25],草本植物对温度、光照、水分、土壤等诸多环境因子的反应差异可能不同程度地影响了物种间的关系,特别是光照强、水分蒸发量大的环境中,草本植物对生境的要求不同及水分等资源利用上的差异,必然会导致物种间对资源的利用性竞争,物种对间往往会表现负相关。负联结种对一般通过生态位的分化,来实现种间的共存,对水分条件有不同需求的种群之间呈负相关;对水分因子适应性相似的种群之间一般呈现正相关[26]。
种间联结的测定方法很多,如χ2检验、联结系数、Jaccard指数、点相关系数等基于2×2列联表的定性分析[27],此方法只与物种在样方中的存在与否有关,因此其结果与样方大小和数目多少紧密相关。Spearman秩相关是一种非参数性检验,不要求数据服从何种分布,具有更高的准确性和灵敏性,应用范围更广,可定量化分析群落植物种间关联程度。本研究中,χ2检验和Spearman秩相关检验结果有一定的相似性,但仍存在着一定的差异。两种检验方法的结果看出2个线叶嵩草群落正负联结数相近,但χ2检验结果中显著关联的种对数较多。引起这种差异结果的原因可能有两种,一方面2×2列联表要求每一格的理论期望值≥5 并要进行连续性矫正,否则χ2检验的结果将会偏大[28]。对于植被数据而言,往往难以满足这一要求,因此某些关联不显著的种对,经检验可能是显著(或极显著)的关联,在一定程度上不能真实反映植物种间联结,这与王凤兰等[29]的研究结果一致;另一方面,2×2列联表是基于二元数据的定性分析,无法了解随着某个种的数量指标(多度、重要值、盖度等)的变化以及另一个种的数量指标的变化情况,因此其局限性强,精确度不高,存在较大误差。所以,以Spearman秩相关检验为主,两种检验方法相互比较,得出的结果才能较好的反映出植物间的关系。
Spearman秩相关分析结果与χ2检验结果在群落某些种对间存在一致性。如线叶嵩草群落中洽草-蒲公英、洽草-毛茛、洽草-珠芽蓼、矮嵩草-委陵菜、蒲公英-黄帚橐吾、委陵菜-珠芽蓼、委陵菜-三脉梅花草等种对间均表现出显著性,退化的线叶嵩草群落中珠芽蓼-异针茅、线叶嵩草-草玉梅、蒲公英-蓝花韭、蒲公英-湿生扁蕾、早熟禾-异针茅、异针茅-湿生扁蕾等均为显著相关。本研究还发现同一种对在不同群落中有不同的关系,如种对线叶嵩草-披碱草在线叶嵩草群落中表现为不显著的正相关,而在退化的线叶嵩草群落中表现为不显著的负相关,这可能由于植物种群在不同群落中具有不同的资源位和生态位所导致[30]。
2个线叶嵩草群落总体上群落植物种间的关系均表现为显著负相关,群落种间关系整体上比较紧密,但群落内大多物种间的联结性较弱,独立性较强,且呈正相关和负相关的种对数在所有种对数中几乎各占一半。本研究2个线叶嵩草群落均处于不稳定阶段。
[1] 王伯荪.植物种群学[M].广州:中山大学出版社,1989:115-120.
[2] HUBALEK.Z.Coefficient of association and similarity based on binary data:an evaluation[J].BiologicalReviews,1982,57(30):669-680.
[3] CHEN Y K(陈玉凯),YANG X B(杨小波),LI D H(李东海),etal.Interspecific associations among dominant plant populations inKeteleeriahainanensiscommunities in Bawangling,Hainan Island[J].JournalofWuhanBotanicalResearch(植物科学学报),2011,29(3):278-287(in Chinese).
[4] 张金屯.数量生态学[M].北京:科学出版社,2011:101-107.
[5] HUANG SH N(黄世能),WANG B S(王伯荪).Dynamics of associations between tree species in a secondary tropical mountain rain forest at Jianfengling on Hainan Island[J].ActaPhytoecologicaSinica(植物生态学报),2000,24(5):569-574(in Chinese).
[6] WANG L(王 琳),ZHANG J T(张金屯).Interspecific association and correlation of dominant species of Lishan Mountain meadow in Shanxi Province[J].ActaBotanicaBoreali-OccidentaliaSinica(西北植物学报),2004,24(8):1 435-1 440(in Chinese).
[7] LOU Y J(娄彦景),ZHAO K Y(赵魁义).Analysis of interspecific association ofCarexlasiocarpacommunity in recent 30-year succession in Sanjiang Plain[J].ChineseJournalofEcology(生态学杂志),2008,27(4):509-513(in Chinese).
[8] CAO G M(曹广民),DU Y G(杜岩功),LIANG D Y(梁东营),etal.Character of passive-active degradation process and its mechanism in alpineKobresiameadow[J].JournalofMountainResearch(山地学报),2007,25(6):641-648(in Chinese).
[9] 周兴民.中国嵩草草甸.北京:科学出版社,2001:51-62,217-234.
[10] LI Q X(李巧峡),ZHAO Q F(赵庆芳),MA SH R(马世荣),etal.Research progress onKobresiaspecies[J].JournalofNorthwest NormalUniversityNaturalScience,Bimonthly(西北师范大学学报·自然科学版),2006,42(6),78-82(in Chinese).
[11] WANG X M(王秀梅),LIU ZH M(刘志民),LIU B(刘 博),etal.Interspecific associations of plants in interdune lowlands of mobile and stabilized dunes in eastern Inner Mongolia of China[J].ChineseJournalofEcology(生态学杂志),2010,29(1):16-21(in Chinese).
[12] WANG H M(王慧敏),PANG CH H(庞春花),ZHANG F(张 峰),etal.Interspecific association of dominant species of wetland vegetation in the middle and lower reaches of Fenhe River,Shanxi Province of China[J].ChineseJournalofEcology(生态学杂志),2012,31(10):2 507-2 512(in Chinese).
[13] FANG F(房 飞),HU Y K(胡玉昆),ZHANG W(张 伟),etal.Numerical analysis of interspecific relationships in alpine steppe community in Bayanbulak[J].ActaEcologicaSinica(生态学报),2012,32(6):1898-1907(in Chinese).
[14] SHI H X(石红霄),FAN Y J(范月君),HOU X Y(侯向阳),etal.Analysis of plant community characteristics ofKobresiapygmaeameadow in the three headwaters under fencing and grazing[J].ChineseJournalofGrassland(中国草地学报),2014,36(3):67-72(in Chinese).
[15] SU SH L(苏淑兰),LI Y(李 洋),WANG L Y(王立亚),etal.Effect of Fencing on plant biomass and functional group structure of different types of degraded grassland in Qinghai-Tibet Plateau[J].ActaBotanicaBoreali-OccidentaliaSinica(西 北 植 物 学 报),2014,34(8):1 652-1 657(in Chinese).
[16] LIU J L(柳剑丽),WANG Z L(王宗礼),LI P(李 平),etal.Quantitative analysis of relationships among species of different communities in typical steppe in Xilingol[J].ChineseJournalofGrassland(中国草地学报),2013,35(5):74-79(in Chinese).
[17] WANG W(王 卫),ZHANG X H(张鲜花),AN SH ZH(安沙舟).Grassland community structure and interspecies relationships in Nalati Pasture[J].ActaAgrestiaSinica(草地学报),2011,19(4):553-559(in Chinese).
[18] ZHANG J Y(张继义),ZHAO H L(赵哈林).Review on the study of vegetation stability[J].ChineseJournalofEcology(生态学杂志),2003,22(4):42-48(in Chinese).
[19] SCHLUTER D.A variance test for detecting species associations with some examples applications[J].Ecology,1984,65(2):998-1 005.
[20] DICE L R.Measures of the amount of ecologic association between species[J].Ecology,1945,26:297-302.
[21] NIU L Q(牛莉芹),SHANGGUAN T L(上官铁梁),CHENG ZH H(程占红).Interspecific relations of the dominant plant populations in the middle part of Zhongtiaoshan Mountain[J].ActaBotanicaBoreali-OccidentaliaSinica(西北植物学报),2005,25(12):1 645-2 471(in Chinese).
[22] LEGENDRE P,LEGENDRE L.Numerical Ecology[M].Amserdam:Elsevier Science,1998:79-86.
[23] YANG Q(杨 琦),ZHOU J(周 婧),TAO CH(陶 楚),etal.Comparative study on interspecific association in tropical evergreen monsoon elfm forest at two succession stages in Tongguling of Hainan Island[J].ChineseAgriculturalScienceBulletin(中 国农学 通报),2014,30(22):8-15(in Chinese).
[24] DU D L(杜道林),LIU Y CH(刘玉成).Studies on the interspecific association of dominant species in a subtropicalAstanopsisfargesiiforest of Jinyun Mountain,China.Acta Phytoecologica Sinica[J].ActaPhytoecologicaSinica(植物生态学报),1995,19(2):149-157(in Chinese).
[25] GASTON K J.The Structure and Dynamics of Geographic Ranges[M].Oxford,Oxford University Press,2003.
[26] QI K(奇 凯),ZHANG CH Y(张春雨),HOU J H(侯继华),etal.Dynamics of species diversity and interspecific associations of herbaceous plants in a Pinus tabulaeformis forest on a sandy site in Chifeng,China[J].ActaEcologicaSinica(生态学报),2010,30(18):5 106-5 112(in Chinese).
[27] KERSHAW K A,LOONEY J H.Quantitative and Dynamic Plant Ecology[M].London:Edward Arnold Publication,1985.
[28] ZHANG F(张 峰),SHANGGUAN T L(上官铁梁).Numerical analysis of interspecific relationship in anElaeagnusmolliscommunity in Shanxi[J].ActaPhytoecologicaSinica(植物生态学报),2000,24(3):351-355(in Chinese).
[29] WANG F L(王凤兰),NIU J M(牛建明),ZHANG Q(张 庆).Study on interspecific relationships among main species ofSfipabrevifloracommunities[J].ActaAgriculturaeBoreali-Sinica(华北农学报).2009,24(1):159-164(in Chinese).
[30] ZHANG J T(张金屯),JIAO R(焦 蓉).Interspecific association between woody plants in Shenweigou of Guandi Mountain,Shanxi Province.[J].BulletinofBotanicalResearch(植物研究),2003,23(4):458-463(in Chinese).
