扁茎大豆与栽培大豆营养器官解剖结构比较
2015-06-28张伟龙吴东梅李春姣睿王桂文陆静梅
张伟龙,李 岩,吴东梅,李春姣,张 睿王桂文,高 伟,陆静梅
(1.吉林省农业科学院大豆研究所,吉林长春130033;2.东北师范大学生命科学学院,吉林长春130024)
扁茎大豆与栽培大豆营养器官解剖结构比较
张伟龙1,2,李 岩2,吴东梅2,李春姣2,张 睿2王桂文2,高 伟2,陆静梅2
(1.吉林省农业科学院大豆研究所,吉林长春130033;2.东北师范大学生命科学学院,吉林长春130024)
利用光学显微摄影技术,比较了两个大豆属植物根、茎、叶和叶柄的解剖结构特征.结果表明:扁茎大豆根内皮层维管形成层带明显,次生木质部导管发达、口径大,且皮层内有菌窝出现;茎的横切面表面积较大,茎中维管束的管孔链数和维管束数目较多,中间髓细胞呈狭长柱形,四周髓细胞发达,体积大、排列紧密.导致扁茎大豆不抗倒伏、光合能力下降以及结实率低的因素可能是:茎的表皮细胞呈方形紧密排列,凯氏带中没有淀粉鞘;叶片只有两层栅栏组织和叶片较薄、海绵组织稀疏;叶柄横切面面积较小.中国普通大豆吉育67叶片有三层栅栏组织,并且海绵组织、栅栏组织和叶片的厚度均明显大于扁茎大豆,有利于植物光合作用;吉育67根的栓质化外皮层发达,对植物体起到了有效的保护作用.
扁茎大豆;营养器官;解剖结构
扁茎大豆来源于美国,是大豆中花序形态特殊的突变体,与普通大豆相比,在形态上有着较大的差异,主要表现为茎秆扁化、顶端花序轴扁平、花荚簇集,是大豆株型改良中非常有利用价值的遗传资源.很多学者曾对扁茎大豆的生物学特性、遗传规律、生理特征等进行了较为深入的研究[1-6].张桂茹等研究指出,中国扁茎大豆R4期光合速率高于高产品种黑农37,低于高光效品种黑农40,但在R5—R6期其光合速率高于黑农40,这种光合特性有利于高光效育种的应用[7-9].但由于扁茎性状由隐性基因(ff)所控制[10],该基因与很多优异农艺性状连锁遗传,即使经过多代连续定向选择也很难稳定,而且扁茎大豆还具有秆软、粒小等弱点,能否稳定地遗传扁茎性状并对其进行改良利用,直接关系到扁茎大豆的利用范围和利用价值.目前的报道通常都是采用扁茎大豆与普通大豆杂交的方法来改良品系,重点选择近似常规亲本类型或介于双亲之间的个体,增加扁茎大豆的多节、多花、多荚的特性,以期在籽粒产量和抗倒伏方面有明显的改进[11-13].
虽然有关普通大豆根、茎、叶和荚果的形成和解剖结构已有大量报道[14-21],但关于扁茎大豆的结构解剖学研究,目前只有零星的报道,尤其是利用结构解剖学方法来分析扁茎大豆的研究尚未见报道.因此本实验采用石蜡切片法和光学显微摄影技术,以美国扁茎大豆和中国普通大豆(吉育67)为研究对象,对其营养器官的解剖结构进行了比较,以为筛选和培育大豆优良品种提供形态解剖学方面的理论依据.
1 材料与方法
1.1 实验材料
以美国扁茎、吉育67为实验材料.材料均采自东北师范大学净月校区的大豆试验基地,材料性状见表1.

表1 两个大豆属植物的外部形态
1.2 实验方法
选取大豆主茎的功能叶片(上数3—4节位),将叶片切成4mm×6mm小块,以及盛花期的花和老嫩适中的大豆根、茎及叶柄(从中部截取5mm长的小段),采用石蜡制片法制作永久封片,然后在光学显微镜下观察并摄影.
2 结果与分析
2.1 根的解剖结构
美国扁茎大豆根的初生木质部为外始式四原型,中央无髓,没有通气组织,内皮层凯氏带加厚明显,薄壁型的傍管薄壁细胞稀疏地排列在导管周围.周皮木栓层薄,厚度约为29.12μm,后生木质部导管数目多达11个,次生木质部导管直径约为37.96μm,外皮层栓质化不明显,皮层中有菌窝形成(见图1).
吉育67根的直径较粗,木质部为外始式四原型,不具髓,内皮层凯氏带加厚明显,栓质化和木质化后形成带状的壁结构,该结构对水分和溶质有着障碍和限制作用.傍管薄壁细胞稀疏排列在导管四周,周皮木栓层较厚,约41.60μm,后生木质部有一个导管,占据髓的位置.次生木质部导管口径较大,直径约51.48μm,次生木质部导管数目为7个.内皮层由一层细胞组成,排列整齐紧密,无胞间隙.皮层中没有菌窝形成(见图1).

图1 两个大豆属植物根的解剖结构
2.2 茎的形态解剖结构
美国扁茎大豆茎的外形呈扁平柱形,大直径约为14.5mm,小直径约为2.6mm;叶序为对生、互生和轮生.表皮细胞呈方形且排列紧密,单细胞表皮毛较多,多细胞腺毛较少.维管束数目较多,排列不均匀,茎中维管束总管孔链数约为146个,大维管束管孔链数较多,多达11个.维管束数目越多,输导能力越强.维管束类型为外韧维管束.韧皮纤维呈团状分布在韧皮部外侧.凯氏带加厚明显,无淀粉鞘.刚分裂未分化的形成层带较活跃,位于初生木质部与初生韧皮部之间.后生木质部较原生木质部大.中部髓细胞形状不规则,呈狭长柱形,四周髓细胞发达,体积大、排列紧密,有利于营养物质的贮藏(见图2).
吉育67茎的外形呈圆柱形,直径较大,约为6.1mm,叶序为互生.单细胞表皮毛较多,表皮毛基呈突起形,多细胞腺毛少.茎的突起处维管束列数较多,达9列,大维管束数目为4列,茎中维管束总管孔链数多,约为54个,维管柱无显著的内皮层和中柱鞘,凯氏带加厚不明显.髓和髓射线发达,均由基本分生组织产生的薄壁组织构成.髓射线位于皮层和髓之间,在横切面上呈放射形,与髓和皮层相通,有横向运输的作用;同时髓和髓射线的功能类似皮层薄壁组织,是茎内贮藏营养物质的组织(见图2).

图2 两个大豆属植物茎的解剖结构
2.3 叶片的解剖结构
美国扁茎的叶片较薄,约143.78μm,单细胞表皮毛和多细胞腺毛较多,下表皮气孔明显,气孔由两个保卫细胞和两个副卫细胞构成,保卫细胞下陷.叶片中毛细脉较多,能增强输导能力.上下表皮由形态相似的椭圆形细胞构成,排列紧密.叶片栅栏组织两层,厚度约67.86μm,栅栏组织延伸至机械组织的现象比较明显.平脉叶肉细胞明显,且形状粗、长,海绵组织稀疏排列,无规则,厚度约41.08μm.主脉有两枚维管束,维管束的管孔链数约为16列(见图3).
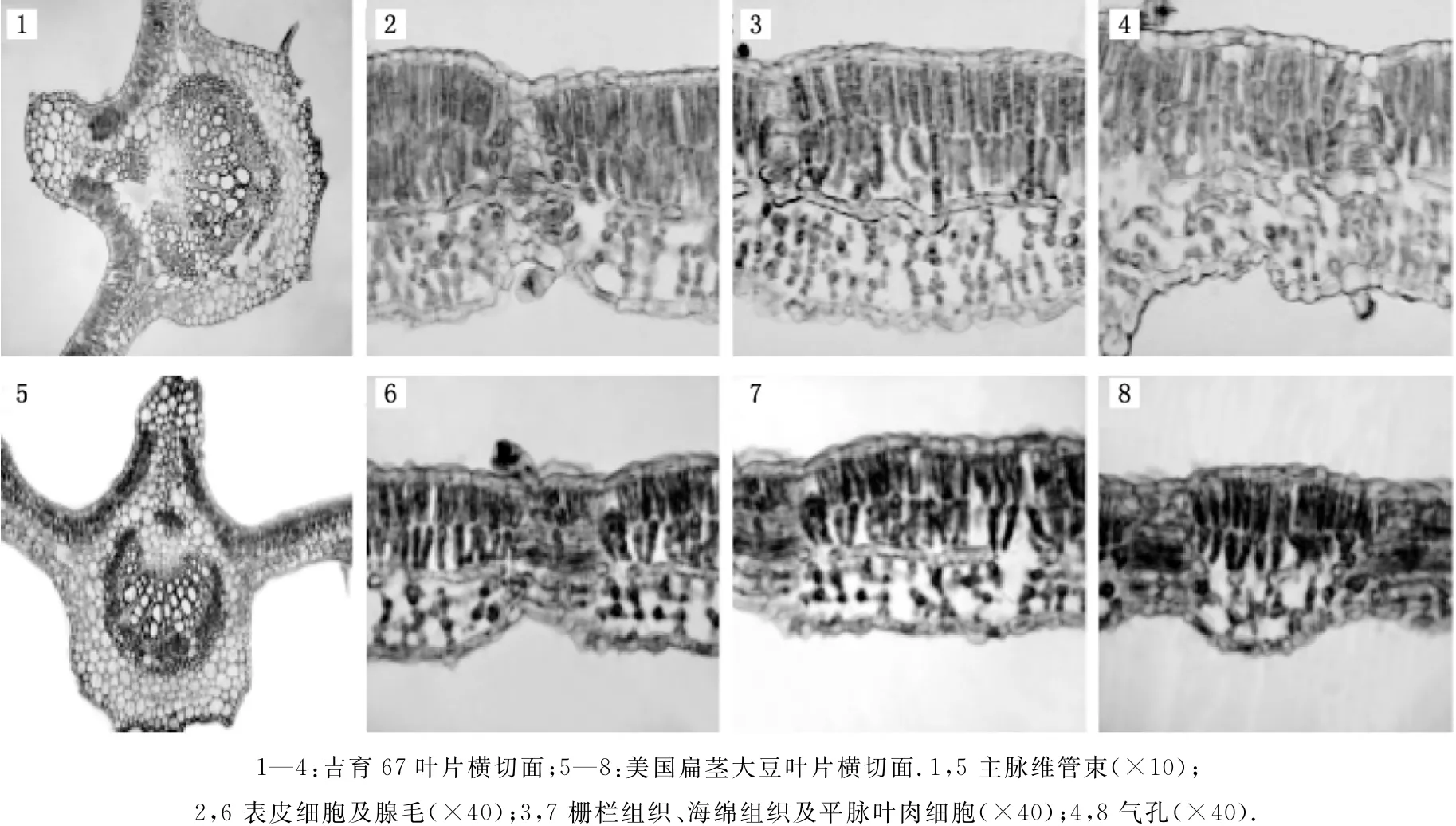
图3 两个大豆属植物叶片的解剖结构
吉育67的叶片较厚,约192.14μm,有单细胞表皮毛和多细胞腺毛.叶片主脉处上下表皮细胞小且排列紧密,下表皮细胞较上表皮细胞长.栅栏组织为3层,较厚,约95.68μm,且延伸到机械组织当中;平脉叶肉细胞明显,形状粗、长,有利于光合作用,相当于裸子植物的传输组织,可传输营养物质.海绵组织稍密集地不规则排列,气孔的保卫细胞不下陷,即保卫细胞、副卫细胞与表皮细胞处于同一平面,属典型的中生植物的结构特点.主脉维管束管孔链数约为16列(见图3).
2.4 叶柄的解剖结构
美国扁茎叶柄的外形像“凹”字,叶柄“凹”字形突起处具有发达的维管束,除去突起部分,近似于圆形,大直径约3.1mm,小直径约2.9mm,表皮细胞小,呈方形或不规则形,且排列紧密.有淀粉鞘,壁厚、腔小的韧皮纤维呈团状分布在韧皮部外面,薄壁细胞中有单晶体,并且薄壁细胞的形状大小不等.叶柄表面棱角较多,维管束列数多,并且规则排列,即发达维管束与小维管束相间而生.主脉大维管束数目为5个,大维管束管孔链数较多,为5~8列,主脉维管束管孔链数为55个,前端突起管孔链数为6个.基本组织细胞较大且形状不规则.维管束类型为外韧维管束(见图4).
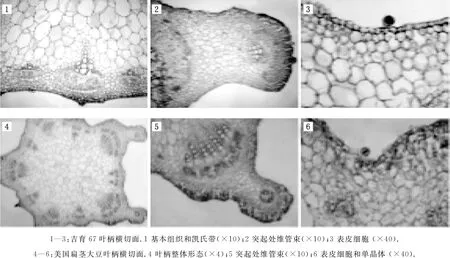
图4 两个大豆属植物叶柄的解剖结构
吉育67叶柄的大直径约3.4mm,小直径约3.2mm,有表皮毛,表皮毛基突起,表皮细胞小且不规则.叶柄“凹”字形突起处各有一枚维管束,维管束管孔链数为7个,内始式外向成熟,主脉大维管束数目为5个,大维管束管孔链数多,为5~6列,主脉维管束管孔链数为40个,除去突起处维管束,其余部分的维管束与茎的维管束相似,基本组织发达,薄壁细胞的形状为椭圆形.有淀粉鞘,韧皮纤维形状为宽带状,维管束排列无规则(见图4).
3 讨论
美国扁茎大豆内皮层细胞显著增大,维管形成层带明显,处于刚刚分裂且未分化的时期,由此可见其活动结果加速了次生组织的形成,为豆科植物的高产提供了可能.次生木质部导管发达、口径大,这一特征为植物体内物质及水分的运输提供了有利条件;皮层内的菌窝是一种高级演化的结构.吉育67根的栓质化(纤维素细胞壁框架添加了栓质物质,即脂类物质)外皮层发达,细胞破碎后形成褐色条状结构,使木栓具有高度不透水性,并有抗压、抗有机溶剂和多种化学药品的特性,对植物体起到了有效的保护作用.
美国扁茎大豆茎的横切面表面积较大,茎中维管束的管孔链数和维管束数目较多,中间髓细胞呈狭长柱形,四周髓细胞发达,体积大、排列紧密,相比栽培大豆,更有利于营养物质的贮藏和输导能力的增强,是优良性状的种质资源.实验还观察到美国扁茎大豆茎的表皮细胞呈方形紧密排列,凯氏带中没有淀粉鞘,与栽培大豆不同,这可能是影响美国扁茎大豆不抗倒伏的因素之一.
高产大豆吉育67有3层栅栏组织,此外,海绵组织、栅栏组织和叶片的厚度均明显大于扁茎大豆,这些都是有利于光合作用的结构特征,无疑能增强单位叶面积光合作用的能力,并能为选育高光效、高产品种和保存优良种质资源提供形态解剖学的依据.同时也能说明,美国扁茎大豆具有的两层栅栏组织、叶片较薄、海绵组织稀疏等特征是导致其光合能力下降、花荚脱落率高的因素之一.吉育67叶片的气孔也明显不同于扁茎大豆,两个保卫细胞和副卫细胞与表皮细胞处于同一平面,属于典型的中生植物的结构特点.
叶柄是连接茎和叶片的器官,在决定叶片角度、植株冠层结构以及光合产物的运输和贮藏碳水化合物等方面具有重要作用.本实验的结果表明:美国扁茎大豆叶柄横切面的表面积明显小于栽培大豆,这一结构特征与美国扁茎大豆的结实率低呈正相关.
[1] 田佩占,袁全,孙永纯,等.改变普通大豆生物学特性提高大豆产量的研究Ⅰ.扁茎大豆的生物学特性[J].大豆科学,1996,16(2):113-117.
[2] 田佩占,袁全,孙永纯,等.改变普通大豆生物学特性提高大豆产量的研究Ⅱ.中国扁茎大豆的生物学特性[J].大豆科学,1998,17(2):95-100.
[3] 田佩占,袁全,王素云,等.改变普通大豆生物学特性提高大豆产量的研究Ⅲ.大豆扁茎性状的遗传方式探讨[J].大豆科学,1999,18(2):95-100.
[4] TANG Y,SKORUPSKA H T.Expression of fasciation mutation in apical meristems of soybean,Glycine max(Leguminosae)[J].American Journal of Botany,1997,84(3):328-335.
[5] KARAKAYA H C,TANG Y,CREGAN P B,et al.Molecular mapping of the fasciation mutation in soybean,Glycine max(Leguminosae)[J].American Journal of Botany,2002,89:558-565.
[6] 石连旋,苗以农,朱长甫.大豆光合生理生态的研究.第18报.不同株型大豆某些生理特性的研究[J].大豆科学,2003,22(2):97-101.
[7] 张桂茹,杜维广,陈怡,等.扁茎大豆光合生理特性及种质改良研究Ⅰ.中国扁茎大豆群体条件下结荚鼓粒期光合特性[J].大豆科学,1999,18(2):134-138.
[8] 张桂茹,杜维广,满为群,等.扁茎大豆光合生理特性及种质改良研究Ⅱ.中国扁茎大豆不同叶位叶片光合特性[J].大豆科学,2000,19(2):115-118.
[9] 栾晓燕,杜维广,张桂茹,等.扁茎大豆光合生理特性及种质改良研究Ⅲ.中国扁茎大豆解剖学观察初报[J].大豆科学,2003,22(2):88-92.
[10] LEFFEL R C,BERNARD R L,YOCUM J O.Agronomic performance of fascinated soybean genotypes and their isogenic lines[J].Crop Science,1993,33:427-432.
[11] 郭泰,王志新,吴秀红,等.扁茎大豆材料的利用与新品种创新[J].中国农学通报,2009,25(23):189-194.
[12] 吴存祥,刘金,李兴宗,等.扁茎大豆的花序形态受光周期调控[J].中国油料作物学报,2004,26(1):36-41.
[13] 姜成喜,吕德昌,陈维元,等.扁茎大豆在育种上的应用研究[J].黑龙江农业科学,2000(1):18-20.
[14] 张达,王豫颖,苍晶,等.矮化大豆突变体叶片解剖结构及过氧化物酶活性研究[J].东北农业大学学报,2008,39(6):67-72.
[15] 陆静梅,刘友良,历锡亮,等.不同进化型大豆花的结构研究[J].应用生态学报,1997,8(4):377-380.
[16] 李晓梅,吴存祥,马启彬,等.大豆品种自贡冬豆花芽分化及开花逆转过程的形态解剖学研究[J].作物学报,2005,31(11):1437-1442.
[17] 李晓梅.大豆茎顶端分生组织石蜡切片的制备[J].大豆科学,2008,27(4):708-710.
[18] 严敏,赵延明,王晓峰.大豆叶柄外植体脱落过程中离区的显微观察[J].中国农学通报,2008,24(8):149-151.
[19] 张涛,张常钟,朱俊义,等.大气变化对大豆属植物叶结构的影响[J].东北师大学报:自然科学版,1999,31(4):62-64.
[20] 庄炳昌,王玉民,徐豹,等.大豆属Soja亚属植物花粉形态的比较观察[J].作物学报,1997,23(1):111-113.
[21] 庄炳昌,王玉民,徐豹,等.大豆属Glycine亚属植物花粉形态研究[J].作物学报,1996,22(3):279-282.
Anatomical structure comparison of nutritive organ between fasciated stem soybean and cultivated soybean
ZHANG Wei-long1,2,LI Yan2,WU Dong-mei2,LI Chun-jiao2,ZHANG Rui2,WANG Gui-wen2,GAO Wei2,LU Jing-mei2
(1.Soybean Research Center,Jinlin Academy of Agricultural Sciences,Changchun 130033,China;2.School of Life Sciences,Northeast Normal University,Changchun 130024,China)
The anatomical structures of roots,stems,leaves and petioles of various plant type soybeans were compared by paraffin slice and light microscopy.The results showed that:compared with other types,fasciated stem soybean has some merits contributed to storage and transportation of nutriments and water:including obvious vascular cambium in root endodermis,developed vascular in secondary xylem,big diameter and bacteria nest in cortex;larger stem transection,more siphonopore chain number of vascular bundle and vascular bundle number,long and narrow column type of intermediate myelocyte cells,developed ambient myelocyte cells,big volume,closely spaced.But the factors that make fasciated soybean prone to lodging,low photosynthetic capacity and low seed setting rate factor may be:stem square epidermal cells closely arranged,casparian strip without starch sheath;only two layer palisade tissue in leaf,thinner leaf,sparse sponge tissue;small petiole transection.In addition,new line 94-2-3-1,namely offspring of fasciated stem soybean,has parental genetic characteristics,big and developed vascular diameter of secondary xylem,and free cortical cell,with cell gap,be helpful for gas storage and enhancing drought resistance,satisfying oxygen supply.Jiyu 67had three layers palisade tissue.The spongy tissue,palisade tissue and the leaf were thicker than the fasciated stem soybean.All these structure features were beneficial to photosynthesis.The suberinized exodermis of Jiyu 67was well developed,and protected the plant effectively.
fascinated stem soybean;nutritive organ;anatomical structure
Q 944.53 [学科代码] 180·5130
A
(责任编辑:方 林)
1000-1832(2015)03-0116-06
10.16163/j.cnki.22-1123/n.2015.03.024
2014-05-12
国家自然科学基金资助项目(41271231);吉林省科技厅专利推进项目(20150312003ZG);现代农作物种业发展专项资金资助项目(2013-2015).
张伟龙(1978—)男,硕士,副研究员,主要从事大豆杂种优势利用研究;通讯作者:陆静梅(1952—),女,博士,教授,博士研究生导师,主要从事结构植物学研究.
