中国特有金钱槭属植物物种分化研究
2015-06-28柏国清李思锋李为民李忠虎赵桂仿
柏国清,李思锋,李为民,李忠虎,杨 娟,赵桂仿*
(1陕西省西安植物园/陕西省植物研究所,西安710061;2西北大学生命科学学院西部资源生物与现代生物技术教育部重点实验室,西安710069;3西安交通大学环境与疾病相关基因教育部重点实验室生物医学研究实验中心/医学院遗传学与分子生物学系,西安710061)
中国特有金钱槭属植物物种分化研究
柏国清1,2,李思锋1,李为民1,李忠虎2,杨 娟3,赵桂仿2*
(1陕西省西安植物园/陕西省植物研究所,西安710061;2西北大学生命科学学院西部资源生物与现代生物技术教育部重点实验室,西安710069;3西安交通大学环境与疾病相关基因教育部重点实验室生物医学研究实验中心/医学院遗传学与分子生物学系,西安710061)
金钱槭属(Dipteronia Oliv.)隶属于槭树科(Aceraceae),是北温带植物区系中古老、残遗的木本属之一,为中国特有属,属下仅包括金钱槭(D.sinensis Oliv.)和云南金钱槭(D.dyerana Henry)2个物种。本实验从形态学、分子遗传学及基因序列分析等方面对金钱槭(D.sinensis Oliv.)和云南金钱槭(D.dyerana Henry)进行对比研究,探讨其物种间分化程度。结果表明:(1)形态学研究显示两物种形态性状存在显著差异,表明其在形态上发生了显著分化;(2)基因组DNA的AFLP标记分析表明,两物种间存在极显著遗传分化(ΦST=0.390 4),在物种水平及群体水平,两物种遗传多样性及遗传变异水平皆有明显差异(金钱槭>云南金钱槭);(3)从GenBank数据库中选出2物种已知的叶绿体基因(psbM-trnD、trnD-trnT、trnL、trnL-trnF、rpl16)和核基因ITS(ITS1、5.8SRNA、ITS2)进行序列进化比对分析,t-test检验表明,2物种叶绿体基因序列间有极显著差异。研究结果表明:金钱槭和云南金钱槭两物种间产生了极显著的分化,推测可能由于长期的地理隔离所致。
金钱槭;云南金钱槭;形态学;分子遗传学;种间分化
金钱槭属(Dipteronia Oliv.)隶属于槭树科(Aceraceae),是北温带植物区系中古老、残遗的木本属之一,为中国特有属[1]。该属植物为落叶乔木,花期4~6月,果期8~10月,染色体2n=18,属于少种属。属下仅包括云南金钱槭(D.dyerana Henry)和金钱槭(D.sinensis Oliv.)2个物种,分别被列为国家二级、三级珍稀濒危保护植物。两物种形态特征存在显著差异,二者不同之处在于金钱槭为圆锥状花序无毛,而云南金钱槭圆锥状花序有密的黄绿色短绒毛;金钱槭果实小于云南金钱槭。两物种在自然地理分布区上也存在着明显不同,无重叠分布区。金钱槭分布于中国陕西、甘肃、河南、湖北、湖南、贵州、四川及重庆等地。而云南金钱槭仅零星分布于中国云南省文山县、蒙自县、屏边县等地区[2]。长期以来,关于金钱槭属植物的研究多集中在该属的形态发育和系统分类方面[3-6]。鉴于金钱槭属内两物种生物学性状存在明显差异,本研究拟对金钱槭和云南金钱槭进行形态学比较研究,检测两物种的形态分化程度。
AFLP(amplified fragment length polymorphism)标记是一种用于群体分子生态、群体遗传研究的有效方法[7]。近年来在种群生态、品系鉴定、基因定位、遗传多样性研究等方面表现出广泛的应用价值。AFLP是显性标记,该技术不但具备其他DNA分子标记如:RAPD、RFLP、SSR等所具有的优点,而且还具有用量少、灵敏度高、扩增条带丰富、可重复性高、快速高效等特点,一次分析可获得基因组大量的遗传信息,并可检测出大量的多态性条带,特别是对具亲缘关系及遗传上区别不大的种类来说是较合适的分子标记方法[8-9]。本研究将结合形态学、基于基因组DNA的AFLP标记对金钱槭属内两物种进行比较研究,分析两物种的形态分化程度,检测两物种的遗传多样性水平,揭示其种间分化程度,探讨种间亲缘关系。另外结合GenBank中两物种已知的几个叶绿体基因及核基因的序列比对结果,从分子水平进一步揭示两物种间的分化水平,为金钱槭属及槭树科植物的进化生物学研究奠定理论基础。
1 材料和方法
1.1 研究材料
本研究所用的植物材料分别采自金钱槭自然地理分布区陕西、河南、甘肃、四川、重庆、湖北、湖南、贵州等8个省区的13个居群,云南金钱槭采自云南境内4个居群(表1)。为保证取样的可靠性,对各个居群中的每个被调查个体,从东、南、西、北4个方向选最基部分枝和近基部第3个分枝,从分支中部选取2对羽状复叶,将复叶上近基部的6片小叶摘下浸于FAA固定液中;并从东、西、南、北4个方向,分上下2层在果序上选取成熟翅果,并压制成标本;同时采集金钱槭、云南金钱槭各个居群新鲜幼嫩的植物叶片,迅速装入密封袋用硅胶迅速干燥,带回实验室置于超低温冰箱(-80℃)保存,用于分子遗传学实验。
1.2 研究方法
1.2.1 形态学研究 参照李珊[10]的方法,形态学性状研究共测量17项宏观、微观形态性状。每个居群取3个个体的平均值作为该居群的各项指标数值。用SPSS 11.0软件包计算同种内不同居群各项形态指标的平均值±标准误差,用OriginPro 7.5软件对2种植物各项形态学指标数值分别进行单因素方差分析,在0.01及0.05水平下对2物种总样本的各项数值进行显著性t检验。
1.2.2 群体遗传学研究 采用植物基因组DNA的AFLP分子标记对金钱槭和云南金钱槭进行遗传多样性、遗传结构和种间分化研究。采用改良的CTAB法提取金钱槭属植物基因组DNA[11-12]。
AFLP反应按照酶切连接一步法、预扩增-选择性扩增两步法进行[12]。选择性扩增从32对PstⅠ+3/MseⅠ+3引物组合中筛选出多态位点检出率高、扩增结果稳定的8对引物组合(P-GAA/MCAA、P-GAA/M-CAC、P-GAA/M-CAG、P-GAG/MCAA、P-GAG/M-CAA、P-GAG/M-CAC、P-GAG/MCTG、P-GAG/M-CTT),对金钱槭和云南金钱槭进行比较研究。
扩增结果按照条带的有无(显性、隐性)记为0、1数据,形成AFLP原始数据矩阵。应用POPGENE(ver.1.31)[13]计算种水平、群体水平的多态位点百分率(PPB)、Shannon多样性指数(I)、Nei基因多样性指数(H)以及种内居群间遗传分化系数(Gst)。检测种内群体间基因多样度(Ht)以及群体内基因多样度(Hs)。AMOVA(Analysis of Molecular Variation)用来分析种间遗传分化系数(ΦST)以及种内居群间、居群内遗传变异成分(Vc)。
1.2.3 序列比对分析 从GenBank搜索金钱槭和云南金钱槭叶绿体基因(psbM-trnD、trnD-trnT、trnL、rpl16、trnL-trnF)以及核基因ITS(ITS1、5.8SRNA、ITS2),采用BioEdit v5.0.6[14]软件中ClustalW进行序列比对分析,计算插入缺失、成对序列的相似性等。采用MEGA v5.0[15]分析成对序列的碱基组成、相同位点、转换、颠换、转换与颠换比值、同一基因及间隔区序列间的遗传距离。采用SPSS 11.0软件对两物种已知叶绿体基因序列进行显著性检验。

表1 2个物种的居群信息Table 1 Population resource of two species
2 结果与分析
2.1 形态学分析
对金钱槭和云南金钱槭植物的17项形态指标(表2)分析比较发现,云南金钱槭小叶长、小叶宽、小叶脉数及复叶叶柄长高于金钱槭,且有显著差异。云南金钱槭果实长、果实宽、果实厚及果实重量高于金钱槭,且有显著差异。云南金钱槭种子长、种子宽、种子厚及种子重显著高于金钱槭。通过结果分析发现,两物种形态水平存在显著差异,较明显的差异是果实的大小、种子的重量及小叶形态。
2.2 遗传多样性
通过筛选出的8对引物组合对金钱槭和云南金钱槭进行选择性扩增,共得到324个扩增片段,由表2可以看出,金钱槭种内多态性片段(PPB)为86.73%,云南金钱槭为78.09%;Nei’s基因多样性指数(H)在金钱槭和云南金钱槭物种水平分别为0.332 1和0.304 7;Shannon多样性指数(I)在2物种水平分别为0.487 4和0.445 0。在群体水平,3个指数(PPB,H,I;表3)在金钱槭种群内平均值为57.29%、0.234 3、0.339 5,在云南金钱槭种群内为52.16%、0.204 4、0.299 1。AMOVA分析表明在金钱槭种内有21.53%变异发生在群体间,78.47%变异存在于群体内;云南金钱槭种内有26.49%变异发生在群体间,73.51%变异存在于群体内。

表2 2个物种的形态比较Table 2 Morphological comparation of two species

表3 2个物种间遗传参数的比较Table 3 A comparison of genetic parameters between two species
2.3 遗传分化
本研究对金钱槭、云南金钱槭两物种的基因多样性(Nei’s)进行分析,结果(表3)表明:金钱槭的基因多样度为0.333 1(Ht)和0.234 2(Hs),云南金钱槭为0.303 5(Ht)和0.204 4(Hs);两物种居群间遗传分化指数(Gst)分别为0.297 0和0.326 6;基因流(Nm)分别为1.183 8和1.031 0,AMOVA分析表明金钱槭与云南金钱槭两种间存在极显著的遗传分化(ΦST=0.390 4,P<0.001)。
金钱槭和云南金钱槭叶绿体基因(psbM-trnD、trnD-trnT、trnL、rpl16、trnL-trnF)和核基因ITS(ITS1,5.8SRNA,ITS2)序列比对结果(表4)表明,2物种叶绿体基因和核基因序列间发生了较大的碱基替换。其中,转换与颠换比值在叶绿体基因序列间为0.8~1.4,在核基因ITS序列为3.1。叶绿体基因序列的显著性检验结果表明,2物种已知叶绿体基因序列间存在极显著差异(P=0.006 8<0.01)。
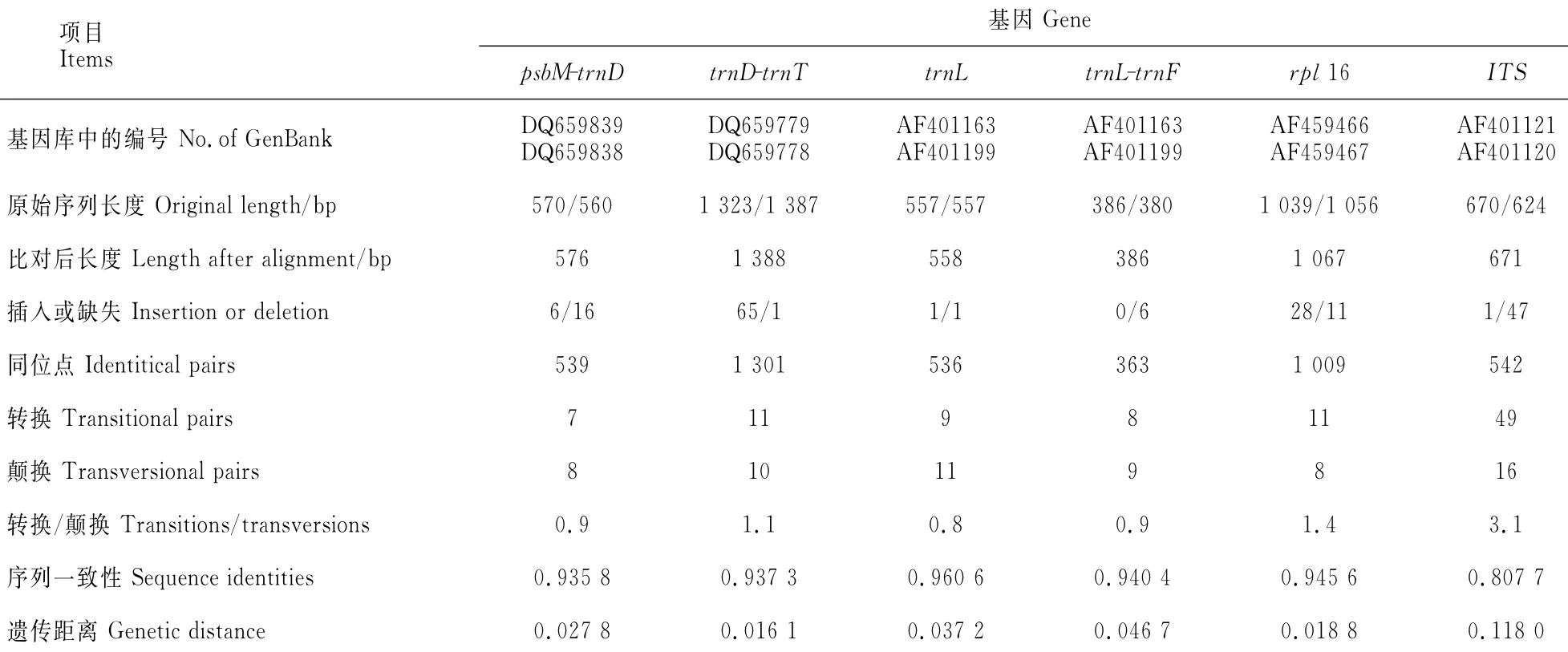
表4 基因库中金钱槭和云南金钱槭的序列分析Table 4 The alanalysis of sequence pairs of species D.sinensis and D.dyeranafrom GenBank
3 讨 论
关于物种不同的研究方向其定义不同。在分类学上,物种是依据表型特征识别和区分生物的基本单位。在现代遗传学上,物种是一个具有共同基因库,与其他类群有生殖隔离的群体。在生态学上,物种是生态系统中的功能单位,不同物种占有不同的生态位,如果2个物种以相似的方式利用同一有限的资源和能源,他们必定会发生竞争和相互排斥;必定有一个获得相对的胜利,如果一个物种的种内发生了变异,占据了多个生态位,从生态学角度看,就意味着新种的生成。对于金钱槭属,经典分类学根据其属内植物的表型特点,将其分为金钱槭和云南金钱槭,本研究采用了17个常用形态指标对两物种进行对比研究发现,金钱槭和云南金钱槭在形态上已产生显著的分化。最显著的形态差异是果实及种子的大小和重量,云南金钱槭远大于金钱槭平均水平;其次是小叶形态,云南金钱槭也明显较金钱槭大。在生态学上,金钱槭和云南金钱槭作为2个独立的物种,具有各自的生态位,无重叠的分布区。分子遗传学研究表明:金钱槭属内两种间存在极显著的分化(ΦST=0.390 4;P<0.001),且两物种的遗传多样性和居群遗传结构存在明显的差异。金钱槭种内、群体平均水平遗传多样性及遗传变异皆高于云南金钱槭,这与广布种的遗传多样性高于狭域种相符。物种的遗传多样性大小是长期进化的产物,是其生存(适应)和发展(进化)的前题[16]。物种的遗传多样性越高、遗传变异越丰富,对环境变化的适应能力越强,越容易扩散其分布范围和开拓新的环境[17]。理论推导和大量实验证据表明,生物居群中遗传变异的大小与其进化速率成正比[18]。因此,对遗传多样性的研究可以揭示物种的进化历史。
而本研究中金钱槭种内、居群内遗传多样性均高于云南金钱槭,推测可能的原因有以下几方面:首先,田欣等[4]通过广义形态学研究和系统分子学对槭树科42种进行研究表明:金钱槭是槭树科中最原始的类群,位于槭树科系统树的最基部。而云南金钱槭位于系统树的中部,是较进化的类群。因此本研究认为:金钱槭较云南金钱槭古老、原始,其进化历史较云南金钱槭长,在较长的进化过程中积累了更多的遗传变异。其次,李珊等[6]采用叶绿体DNA对金钱槭属植物进行的亲缘地理学研究发现,金钱槭具有的叶绿体DNA单倍基因型种类多于云南金钱槭,单倍型分布模式较为复杂,推测冰期时金钱槭存在于多个“避难所”,现今存活的金钱槭来源于多个不同的“避难所”,初始种群间的差异较大,现存个体的遗传物质多样化水平高于云南金钱槭。
本研究揭示金钱槭和云南金钱槭两种间产生了极其显著的分化,这与前人的研究较一致。广义形态学研究和系统分子学研究[19-20]都显示,云南金钱槭与槭属内部分种聚在一起,而不是直接与其同属的位于槭树科系统树基部的金钱槭聚在一起。可见,金钱槭与云南金钱槭确实存在显著的分化。其产生分化的原因可能是金钱槭和云南金钱槭本身就来源于不同的起源模式。到目前为止,还未发现云南金钱槭与金钱槭有重叠分布区,长期的地理隔离也是造成其种间分化加剧的因素之一。
[1] 应俊生,张玉龙.中国种子植物特有属[M].北京:科学出版社,1994:536-539.
[2] 方文培.中国植物志[M].北京:科学出版社,1981.
[3] ZHAO X G(赵先贵),XIAO L(肖 玲),MAO F CH(毛富春).Pollen morphology of Aceraceace[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),1998,18(2):252-255(in Chinese).
[4] TIAN X(田 欣),JIN Q J(金巧军),LI D ZH(李德铢).Pollen morphology of Aceraceae and its systematic implication[J].Acta Botanica Yunnanica(云南植物研究),2001,23(4):457-465(in Chinese).
[5] QI Y ZH(祁云枝),DU Y J(杜勇军),LI L M(李莲梅).The rare and endangered plants of Shaanxi Province for exsitu conservation in Xi’an Botanical Garden[J].Journal of Northwest Forestry University(西北林学院学报),2001,16(1):33-36(in Chinese).
[6] LI SH(李 珊),CAI Y L(蔡宇良),XU L(徐 丽),et al.Morphological differentiation of samaras and seeds of Dipteronia dyeriana,Aceraceae[J].Acta Botanica Yunnanica(云南植物研究),2003,25(5):589-595(in Chinese).
[7] ZABEAU M,VOS P.Selective restriction fragment amplification:ageneral method for DNA fingerprinting[J].European Patent Application,1993,31(3):1-47.
[8] POWELL W,MACHRAY G,PROVAN J.Polymorphism revealed by simple sequence repeats[J].Trends Plant Sci.,1996,1(7):215-222.
[9] HILL M.,WITSENBOER H,ZABEAU M,et al.PCR-based fingerprinting using AFLPs as a tool for studying genetic relationships in Lactucaspp.[J].Theoretical and Applied Genetics,1996,93(8):1 202-1 210.
[10] 李 珊.金钱槭属植物保护遗传学与分子亲缘地理学研究[D],西安:西北大学,2004.
[11] 王关林,方宏筠.植物基因工程原理与技术[M].北京:科学出版社,1998:370-375.
[12] YANG J(杨 娟),LI SH(李 珊),CAO D W(曹东伟),et al.Construction and analysis of the AFLP reaction system of Dipteronia Oliv.[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2005,25(11):2 173-2 177(in Chinese).
[13] YEH F C,YANG R C.Popgene Computer Program(ver 1.31)[M].University of Albert and Center for International Research,Canada,1999.
[14] HALL T A.Bioedit:A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT[J].Nucleic Acids Symposium Series,1999,41:95-98.
[15] TAMURA K,PETERSON D,PETERSON N,et al.MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Mol.Biol.Evol.,2011,28(10):2 731-2 739.
[16] SOLTIS P S,SOLTIS D E.Genetic varitation in endemic and widespread plant species examples from Saxifragaceae and Polystichum[J].Aliso,1991,13(1):215-223.
[17] HUENNEKE L F.Ecological Implications of Genetic Variation in Plant Populations[M].New York:Oxford University Press,1991:31-44.
[18] AYALA F J,VALENTINE J W.Genetic variability in the pelagic environment:aparadox[J].Ecology,1979,60(1):24-29.
[19] TIAN X(田 欣),GUO ZH H(郭振华),LI D ZH(李德铢).Phylogeny of aceraceae based on ITS and trnL-F data sets[J].Acta Botanica Sinica(植物学报),2002,44(6):714-724(in Chinese).
[20] XU T ZH(徐廷志).The systematic evolution and distribution of the genus acer[J].Acta Botanica Yunnanica(云南植物研究),1998,20(4):383-393(in Chinese).
(编辑:宋亚珍)
Interspecific Differentiation of the Endemic Plant Genus Dipteronia Oliv.
BAI Guoqing1,2,LI Sifeng1,LI Weimin1,LI Zhonghu2,YANG Juan3,ZHAO Guifang2*
(1Xi’an Botanical Garden of Shaanxi Province/Institute of Botany of Shaanxi Province,Xi’an 710061,China;2Key Laboratory of Resource Biology and Biotechnology in Western China,Ministry of Education,College of Life Science,Northwest University,Xi’an 710069,China;3Department of Genetics and Molecular Biology,Medical School of Xi’an Jiaotong University/Key Laboratory of Environment and Disease-related Gene,Ministry of Education,Xi’an 710061,China)
The genus Dipteronia Oliv.(Aceraceae)is endemic in China which consists of two species:D.sinensis Oliv.and D.dyerana Henry.Dipteroniais one of the woody relic genera in the floristic regions of the northern temperate zone.In this study,we carried out a comparative study of D.sinensis Oliv.and D.dyerana Henry.to explore their degree of differentiation between two species from the aspects of morphology,molecular genetics and gene sequence analysis.(1)Morphological studies indicated that the presence of two species in morphological characters have significant differences.(2)Molecular genetic studies based on the AFLP of genomic DNA markers were also used to analyze the two species.And whether they are in species level or group level,the results showed that there was extremely significant genetic differentiation between the two species(ΦST=0.390 4).The level of genetic variation and genetic diversity of the two spe-cies have distinct differences(D.sinensis Oliv.>D.dyerana Henry).(3)We also downloaded several chloroplast genes(psbM-trnD,trnD-trnT,trnL,trnL-trnF,rpl16)and nuclear gene sequences of ITS(ITS1,5.8SRNA,ITS2)of two known species from GenBank to analyze and compare their differentiation.t-test showed a significant difference between the two species occurred in chloroplast gene sequences.Hence,the results of the present study showed that the D.sinensis Oliv.and D.dyerana Henry have a extremely significant differentiation.And we infer the main reasons of the significant differentiation between the two species attributed to the long-term geographic isolation between them.
Dipteronia sinensis Oliv.;D.dyerana Henry;morphology;molecular genetics;interspecific differentiation
Q789;Q941+.3
A
10.7606/j.issn.1000-4025.2015.06.1123
1000-4025(2015)06-1123-06
2014-06-14;修改稿收到日期:2015-05-20
国家自然科学基金(31470311);教育部博士点基金优先发展领域(20136101130001);陕西省财政厅项目(2010JM5005);陕西省科学院科技计划(2011-K03)
柏国清(1983-),男,在读博士研究生,助理研究员,主要从事系统与进化植物学研究。E-mail:bgq@ms.xab.ac.cn
*通信作者:赵桂仿,教授,主要从事植物系统进化与分子生态研究。E-mail:gfzhao@nwu.edu.cn
