中华大蟾蜍视网膜Müller细胞形态观察
2015-06-27杨鹏飞赵留涛赵新芳闫峰宾杜宝宝刘忠虎河南农业大学牧医工程学院河南郑州450002
杨鹏飞, 赵留涛, 赵新芳 ,闫峰宾 ,杜宝宝, 刘忠虎(河南农业大学牧医工程学院, 河南 郑州 450002)
中华大蟾蜍视网膜Müller细胞形态观察
杨鹏飞, 赵留涛, 赵新芳 ,闫峰宾 ,杜宝宝, 刘忠虎
(河南农业大学牧医工程学院, 河南 郑州 450002)
为了解两栖动物中华大蟾蜍(Bufo gargarizans)视网膜中神经胶质细胞形态结构及分布,利用生物显微技术、投射电镜技术和免疫组化技术显示了中华大蟾蜍视网膜胶质细胞形态结构。观察结果显示,中华大蟾蜍视网膜中仅分布有Müller细胞。 光镜下Müller细胞在视网膜中成单层分布,细胞长轴沿着眼球半径成放射状排列,胞体位于内核层,胞体发出至少2个主干突起。电镜下视网膜Müller细胞的胞质及胞核着色较周围细胞深,细胞内有明显的纤维成分,与其他神经成分易于区别。Müller细胞近巩膜侧的主干突起延伸到视杆与视锥细胞的内节与胞核交界处,并彼此相互连接形成外界膜,近玻璃体侧的突起末端形成圆锥形的终足,锥形的底部紧邻玻璃体,且在此处彼此相互连接形成内界膜。
中华大蟾蜍;视网膜;Müller 细胞
脊椎动物视网膜含有多种胶质细胞,如星形胶质细胞(Astrocytes)、小胶质细胞(Microglial cell)、少突胶质细胞(Oligdendrocytes)和Müller细胞等。但不同种的脊椎动物视网膜胶质细胞分布类型存在差异。FISCHER等[1]认为,小胶质细胞和Müller细胞是视网膜中稳定存在细胞,星形胶质细胞和少突胶质细胞在不同物种视网膜中存在情况不同。如在鸡、豚鼠和兔等有血管视网膜(Vascular retina)中分布有少突胶质细胞,它们形成神经纤维层(Nerve fiber layer ,NFL) 节细胞轴突的髓鞘[2-4],同时有血管视网膜中也分布有大量星形胶质细胞,这些细胞紧密贴附在视网膜内血管的壁上[5-7], 而内部无血管分布的视网膜几乎没有星形胶质细胞[8]。研究认为星形胶质细胞并非由视网膜神经上皮细胞分化形成,而是在视网膜发育过程中沿着视神经由脑部迁移而来的[9,10]。少突胶质细胞和星形胶质细胞是有共同祖细胞分化形成的[11]。小胶质细胞是在视网膜发育早期阶段由视神经处的中胚层细胞分化形成,且形成后潜伏入视网膜层中,或由伴随血管延伸入视网膜的细胞而来[12]。Müller细胞是视网膜中主要胶质细胞,也是仅存在于视网膜中的胶质细胞,且该类细胞与视网膜神经元是由同一祖细胞分化形成的[13]。 Müller细胞对视网膜中的神经元不仅起着支持、营养、保护、绝缘等功能,而且具有其他多种重要功能[14-15]。如视网膜内Müller细胞表达大量的电压门控通道(Voltage-gated channels)和神经递质受体(Neurotransmitter receptors),可以识别多种神经信号、触发细胞去极化和引起细胞内Ca2+流变化。此外,Müller细胞可以通过调节细胞外神经活性物质的浓度影响视网膜中神经元的活动,如Müller细胞可以通过其高亲和性载体的吸收作用调节谷氨酸和GABA浓度,通过Na+-HCO3-共转移作用和碳酸酐酶(Carbonic anhydrase)的酶促作用来调节H+浓度等。Müller细胞和神经元之间的双向信息交流说明了其在视网膜中起着非常重要的作用。另外,一些研究报道Müller细胞与视网膜损伤后神经元再生有关,是神经祖代细胞(Nerve progenitor)来源之一[16~18]。关于两栖动物视网膜胶质细胞国外已有部分资料报道[14,19-21],KALININA[21]对湖蛙(RanaridibundaPall)研究认为该蛙视网膜中存在4种胶质细胞: Müller 细胞、少突胶质细胞、小胶质细胞和spider 细胞。SKATCHKOV等[22]认为,在两栖类视网膜中Müller细胞是唯一的胶质细胞。国内关于Müller细胞仅有少量对人视网膜Müller细胞报道[23-24],对两栖动物视网膜Müller细胞形态和分布还未见报道,两栖动物视网膜Müller细胞数量较多,易于分离[14],是用于研究Müller细胞的形态结构和生理功能理想的模型系统。本研究利用光镜、电镜技术和免疫组化染色法显示胶质细胞中特有的胶质纤维酸性蛋白(Glial fibrillary acidic protein, GFAP),系统观察了中华大蟾蜍Müller细胞在视网膜中的形态和分布特征,为研究Müller细胞在两栖类视网膜中的生理功能和视神经损伤再生过程中的作用提供形态学基础。
1 材料与方法
1.1 试验动物
健康3年龄成体中华大蟾蜍(Bufogargarizans)10只(雄性6只,雌性4只),平均体长(9±0.3)cm。由河南农业大学动物房提供。
1.2 光镜观察
中华大蟾蜍用2% MS-222麻醉后迅速摘除眼球,清除球外眼组织角膜、晶状体及玻璃体后,将组织立即投入预冷Bouin,s液固定2 d,常规石蜡包埋,切片厚度6 μm,选取过眼球最大直径的切面, H.E染色后显微镜下(BA310 DIGITAL C,MOTIC厦门)观察并采集数码照片(Motic Advanced3.2)和进行显微测量统计。
1.3 免疫组织化学
眼睛常规石蜡包埋和切片(同1.2),经二甲苯脱蜡和乙醇复水,PBS(0.01 mol·L-1,pH7.4)冲洗3次,每次5 min,3%H2O2室温15 min灭活内源性过氧化物酶活性,PBS冲洗, 正常羊血清封闭室温1 h,鼠抗GFAP抗体,4℃过夜。PBS冲洗,羊抗兔IgG室温1 h,辣根过氧化物酶标记链霉卵白素室温1 h,DAB显色5 min,苏木精复染,中性树胶封片。阴性对照切片以正常羊血清替代一抗。显微镜下观察并采集数码照片和进行显微测量统计。
1.4 透射电镜
中华大蟾蜍用2% MS-222麻醉后迅速摘除眼球,清除球外眼组织角膜、晶状体及玻璃体后,经2.5%戊二醛-1.5%多聚甲醛固定液中固定2 d,在视网膜中央区和边缘区分别剪取大小2 mm×3 mm视网膜组织块,经1%锇酸后固定1 h,乙醇-丙酮脱水,Epon812树脂包埋,半薄切片定位,选取过眼球半径的切面和过眼球切向的切面进行超薄切片,醋酸铀-柠檬酸铅染色。电镜(JEM-100SX型)观察和采集数码图像。
2 结果与分析
2.1 光镜下中华大蟾蜍视网膜组织形态
中华大蟾蜍视网膜常规组织学观察可见明显的内界膜(Inner limiting membrane, ILM)与外界膜(Outer limiting membrane, OLM)(图1),Müller细胞即分布于视网膜外界膜与内界膜之间的区域。

中华大蟾蜍视网膜组织形态,H.E染色示视网膜各层结构。RPE,视网膜色素上皮;OLM,外界膜;ONL,外核层;OPL,外网层;INL,内核层;IPL,内网层;GCL,节细胞层;NFL,神经纤维层;ILM,内界膜。
The histological section of retina showing the structure of retina ofBufogargarizansstaining with H.E. RPE,retina pigmented epthelium; OLM,outer limiting menbrane; ONL,outer nuclear layer; OPL,outer plexiform layer; INL,inner nuclear layer; IPL,inner plexiform layer; GCL,ganglion cell layer; NFL,nerve fiber layer; ILM,inner limiting membrane.
图1 中华大蟾蜍视网膜组织形态
Fig.1 Structure ofBufogargarizansCantor’s retina
视网膜切片免疫组化染色发现,在沿视网膜辐向切面和切向切面上均可见到免疫组化染色呈棕色的阳性结构,在辐向切面上可以观察到Müller细胞成单层排列,细胞长轴沿着眼球半径方向成放射状排列(图2-A)。在视网膜辐向切面上Müller细胞长度为(62.1±0.54)μm;每100 μm 长度视网膜切面中Müller细胞的密度为(6.2±0.1)个。
Müller细胞胞体位于内核层(Inner nuclear layer,INL)中部(图2-B),自胞体分别发出伸向视网膜近玻璃体侧和近巩膜侧的至少2个主干突起(Stem processe,sp)。伸向视网膜近玻璃体侧的主干突起(Vitread stem processe,vsp)终止于节细胞层(Ganglion cell layer, GCL)或神经纤维层(Nerve fiber layer, NFL),且其主干终末膨大为圆锥型终足(endfood,ef)(图2-A)。伸向视网膜近巩膜侧的主干突起(Sclera stem process,ssp)终止于外核层(Uuter nuclear layer,ONL)外侧,突起之间相互联接包绕在每个光感受器细胞的周围形成外界膜(图2-A, 图2-B)。对内网层中的vsp和内核层ssp的横切面的面积和个数进行了统计。其中内网层每100 μm 长度视网膜切面中含vsp个数为(43.4±0.42)个,每个面积为(3.61±0.32)μm2;内核层100 μm 长度视网膜切面中含ssp个数为(27.8±0.72)个,每个面积为(9.56±0.65)μm2(图2-C)。

Müller细胞在视网膜中的分布及密度。ef,终足;GCL,节细胞层;INL,内核层;IPL内网层;M,Müller细胞胞体;OLM,外界膜;ssp,近巩膜侧主干突;vsp,近玻璃体侧主干突起。A、B为沿眼球半径方向切面,示Müller细胞纵切面突起结构。C为沿眼球切向切面,示Müller细胞突起结构横切面。
The distribution and density of Müller cell in the retina ofBufogargarizans. ef,endfood;GCL,ganglion cell layer;INL,inner nuclear layer;IPL,inner plexiform layer; M, Müller cell body; OLM, out limiting membrane; ssp, the scleral stem processes; vsp, the vitread stem processes. A,B: Sections cross the radius of eyeball,showing the lognitudinal processes of Müller cell. C:Sections cross the tangent of eyeball,showing tranverse section of Müller cell processes.
图2 中华大蟾蜍视网膜GFAP免疫组化染色
Fig.2 The retinal section staining by immunohistology with antibody to GFAP
由Müller细胞发出的2个方向上的主干突起在几乎横跨整个视网膜同时,又发出许多细小的侧向分支突起,尤其是在内网层(Inner plexiform layer,IPL)与外网层(Outer plexiform layer, OPL)这些侧支充满了神经元树突和轴突之间(图2-C)。
2.2 电镜下观察中华大蟾蜍视网膜Müller细胞
在透射电镜下,视网膜内、外核及节细胞层对其进行了细致的观察。外核层及视锥、视杆细胞层中(图3-A、3-B),Müller细胞胞突向外伸至视锥、视杆细胞外侧与内节之间,形成Müller细胞的近巩膜侧突起ssp,突起内有细丝,应为细胞内的GFAP纤维(图3-B)。Müller细胞外侧突起顶端之间、Müller细胞外侧突起与视细胞内节之间及视细胞内节与内节之间有连接复合体,共同构成外界膜如图中箭头所示(图3-A)。外界膜并不是一层膜,而是由细胞与细胞之间的连接结构粘连小带所构成。在内核层中可见Müller细胞(图3-B中M示Müller细胞)较其他内核层中细胞深染,突起内也可见明显的纤维成分。
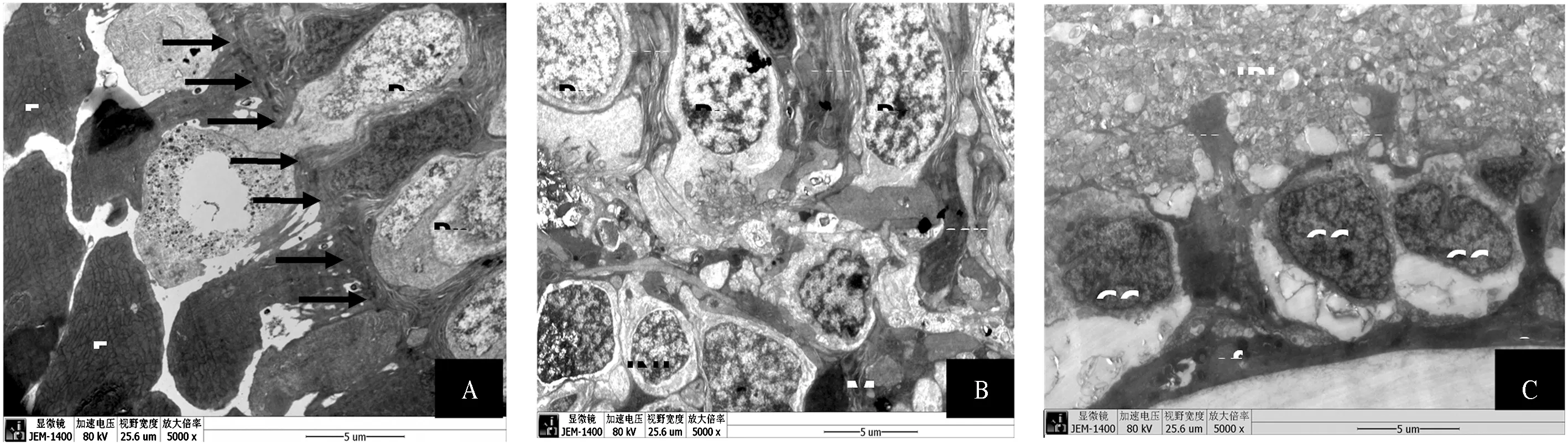
A. 箭头所示为外界膜,E为光感受器椭球体,Pn为光感受器细胞核。B. M为内核层中的Müller细胞,INL为内核层,Pn为光感受器细胞核,ssp为包绕在光感受器细胞外的Müller细胞近巩膜侧突起。C. ef为终足,GC为节细胞核,IPL为内网层,vsp为内核层中的Müller细胞近玻璃体侧突起。
A.Arrows showing the outer limiting membrane;E,ellipsoid body of photoreceptor cell;Pn,the nuclear of photoreceptor cell ;B. M,Müller cells ;INL,inner nuclear layer;ssp,the scleral stem processes. C. ef,endfood;GC,ganglion cell nuclear;IPL,inner plexiform layer; vsp, the vitread stem processes.
图3 中华大蟾蜍视网膜电镜观察
Fig.3 The structures of Müller cells ofBufogargarizanson electron microscopy
内网层中Müller细胞的近玻璃体侧突起vsp较该层中神经细胞突起粗且染色深(图3-C),vsp及其分支在节细胞层中伸于节细胞核之间,形成神经元周围的微环境。vsp末端膨成锥形,形成终足。终足的底部彼此相互连接共同组成视网膜内界膜。内界膜直接与玻璃体相邻,将视网膜细胞与玻璃体隔开。
3 结论与讨论
对Müller细胞形态和分布的研究过去多采用硝酸银浸染法,但这种方法过程复杂,且结果不稳定。以后又出现了特殊的染色法,以及活性染料视网膜整染法[25]。在对人视网膜研究中曾有利用烯醇化酶NNE(Non-neuronal enolase)抗体来显示胶质细胞的[23],但人视网膜中还有其他类型的胶质细胞,且烯醇化酶(Enolase)是糖酵解过程中重要的酶,与细胞的代谢过程密切相关,胶质细胞染色结果受细胞生理状态影响。
本研究利用免疫组化法显示视网膜中的GFAP阳性细胞即Müller细胞,在脊椎动物视网膜中存在的胶质细胞类型不完全与脑组织中的相对应,且因动物进化地位不同而不同,少突胶质细胞和星型胶质细胞是在脑组织中存在的2种大胶质细胞(macroglia),少突胶质细胞在大多数动物视网膜中完全没有分布,星型胶质细胞仅存在于哺乳动物视网膜中,而且星型胶质细胞仅分布于哺乳动物视网膜的NFL;对两栖动物豹蛙(Rana pipiens)研究发现Müller细胞是视网膜中唯一观察到的胶质细胞[22]。另外,本研究观察到的GFAP细胞个体大,其突起横跨整个视网膜层,胞体位于内核层,完全符合 Müller细胞的特征[14-25]。因此,利用抗GFAP抗体显示两栖类的Müller细胞是一种较好的方法。
本研究观察到伸向视网膜近玻璃体侧的主干突起终末膨大为圆锥型终足,终足圆锥型结构的底部紧贴与玻璃体,此结构特点被认为与两栖类动物稳定细胞外K+浓度有关,光激活视网膜神经元,在2个突触层(IPL和OPL) 诱发 K+浓度增高。这种增高的K+必须被迅速清除,否则神经元的兴奋性程度会发生激烈的变动[26]。在两栖类中Müller 细胞首先将神经元释放的多余的K+摄入细胞内,然后通过终足区将大部分K+转移至玻璃体,这种空间缓冲机制清除多余K+的效率比单纯经由胞外空间扩散要高1倍以上[26]。
在内核层的内中间区,从Müller细胞的胞体发出放射状突起,这些坚韧的突起纵贯视网膜全层,几乎占据了神经细胞所没有占据的空间。从形态学上支持Müller细胞是整个视网膜的支架。在神经纤维层,放射状突起的终末端呈圆锥形膨大,参与内界膜的结构。在外核层、内核层及神经节细胞层,从Müller细胞放射状突起的侧壁发出带状分支,这些分支突起形成网状,包绕着神经细胞的胞体,有利于视网膜微环境的稳定。在外网状层、内网状层及神经纤维层,从Müller细胞放射状突起的侧壁向水平方向发出细微的分支,这些水平分支包绕着神经细胞的树突、轴突及其突触。跟中枢神经系统的神经胶质细胞类似,Müller细胞对视网膜神经细胞神经冲动的传递可能会起到绝缘作用。
[1] FISCHER A J, ZELINKA C, SCOTT M A. Heterogeneity of glia in the retina and optic nerve of birds and mammals[J]. PLoS ONE,2010,5(6):e10774.
[2] NARANG H K, Right-left asymmetry of myelin development in epiretinal portion of rabbit optic nerve[J]. Nature,1977,266: 855-856.
[3] KOHSAKA S, TAKAMATSU K, NISHIMURA Y, et al. Neurochemical characteristics of myelin-like structure in the chick retina[J]. J Neurochem,1980,34: 662-668.
[4] WYSE J P, SPIRA A W. Ultrastructural evidence of a peripheral nervous system pattern of myelination in the avascular retina of the Guinea pig[J]. Acta Neuropathol,1981,54: 203-210.
[5] WEST H, RICHARDSON W D, FRUTTIGER M .Stabilization ofthe retinal vascular network by reciprocal feedback between blood vessels and astrocytes[J]. Development ,2005,132: 1855-1862.
[6] FRIEDLANDER M, DORRELL M I, RITTER M R, et al. Progenitor cells and retinal angiogenesis[J]. Angiogenesis ,2007,10: 89-101.
[7] DORRELL M I, FRIEDLANDER M. Mechanisms of endothelial cell guidance and vascular patterning in the developing mouse retina[J]. Prog Retin Eye Res,2006,25: 277-295.
[8] WON M H, KANG T C, CHO S S .Glial cells in the bird retina: immunochemical detection[J]. Microsc Res Tech,2000, 50: 151-160.
[9] STONE J, DREHER Z. Relationship between astrocytes, ganglion cells and vasculature of the retina[J].J Comp Neurol,1987;255:35-49
[10] CHAN L T. Glial neuronal and vascular interactions in the mammalian retina[J]. Prog Ret Eye Res,1994,13:357-389.
[11] ROMPANI S B, CEPKO C L. A common progenitor for retinal astrocytes and oligodendrocytes[J]. J Neurosci, 2010,30(14):4970-4980.
[12] Gallego A. Comparative studies on horizontal cells and a note on microglial cells[J].Prog Ret Eye Res,1986,5:165-206.
[13] TURNER D L, CEPKO C L. A common progenitor for neurons and glia persists in rat retina late in development[J]. Nature,1987;128:131-136.
[14] NEWMAN E, REICHENBACH A. The Müller cell: a functional element of the retina [J]. Trends Neurosci, 1996, 19:307-312.
[15] 杨雄里. Müller 细胞与视网膜功能[J]. 生理科学进展,1998 , 29 (1): 7-10.
[16] FISCHER A J. Neural regeneration in the chick retina [J]. Prog Retin Eye Res, 2005, 24(2):161-182.
[17] FISCHER A J, REH T A. Potential of Müller glia to become neurogenic retinal progenitor cells [J]. Glia, 2003, 43(1):70-76.
[18] HITCHCOCK P, OCHOCINSKA M, SIEH A, et al. Persistent and injury-induced neurogenesis in the vertebrate retina [J]. Prog Retin Eye Re., 2004 , 23(2):183-194.
[19] UGA S S. Comparative study of the fine structure of retinal Muller cells in various vertebrates [J]. Invest Ophthalmol, 1973 , 12(6):434-448.
[20] ZHU B S,GIBBINS I. Müller cells in the retina of the cane toad,Bufomarinus, express neuropeptide Y-like immunoreactivity[J]. Visual Neuroscience,1996, 13: 501-508.
[21] KALININA A V. Glial cells of the retina in Rana ridibunda pall[J],Arkh Anat Gistol Embriol.,1983, 84(4):33-38.
[22] SKATCHKOV S N, KRUSEK J, REICHENBACH A, et al. Potassium buffering by Müller cells isolated from the center and periphery of the frog retina[J]. Glia,1999,27(2):171-180.
[23] 周国民,肖虹蕾, 佘振钰,等.视网膜中 Müller细胞分布的组织化学研究[J]. 上海医科大学学报, 1994,21(5):382-384 .
[24] 李爱冬, 羊惠君. 胎儿视网膜Müller细胞免疫组化研究[J]. 四川解剖学杂志,1995 , 3(2):108-116.
[25] UCKERMANN O, IANDIEV I, FRANCEK M, et al. Selective staining by vital dyes of müller Glial cells in retinal wholemounts[J]. Glia, 2004, 45:59-66.
[26] REICHENBACH A , HENKE A , EBERHARDT W , et al. K+regulation in retina[J]. Can J Physiol Pharmacol, 1992, 70( Suppl):239-247.
(责任编辑:蒋国良)
Research on morphology of retinal Müller cells inBufogargarizans
YANG Pengfei, ZHAO Liutao, ZHAO Xinfang, YAN Fengbin, DU Baobao, LIU Zhonghu
(Engineering College of Animal Husbandry and Veterinary Science, Henan Agricultural University,Zhengzhou 450002,China)
The morphology and distribution of neuroglial cells in retina of Amphibian,Bufogargarizanswas shown by light microscopy,transmission electron microscope and immunohistochemical staining method in the retinas of the toads. The results show that there is only one kind of the neuroglial cell the Müller cells within retina.The Müller cells arrayed in regular monolayer, the long axes of cells stretched radially along the radiuses of eyeball. The somata of the Müller cells lay in the inner nuclear layer and gave rise to at least two opposite stem processes,the scleral stem processes (ssp) and the vitread stem processes (vsp).At the ultrastructural level, retinal Müller cells were deeper in color than the surrounding cytoplasm and nuclei of cells and other neural components.There were lots of visible fibers in the Müller cells processes, so the Müller cells were remarkably different from the other neural cells component of retina. The ssp of Müller cells radially oriented to the base of the inner segments of the photoreceptor cells,here ssp enclosed the photoreceptor cells and formed the outer limiting membrane,the vsp of Müller cells lay the inner plexiform layer and ganglion cell layer,the terminal of vsp were adjacent to the surface of the vitreous,and expanded as conical endfeet,the endfeet junctioned each other and form the inner limiting membrane.
Bufogargarizans;retina;Müller cell
2015-05-21
河南省教育厅自然科学基础与前沿项目(13A240482)
杨鹏飞(1990-),男,河南汝州人,硕士研究生,从事动物细胞生物学方面的研究。
刘忠虎(1964-),男,河南博爱人,副教授,博士。
1000-2340(2015)06-0822-05
Q954.6
A
