模拟降雨量变化与CO2浓度升高对羊草光合特性和生物量的影响
2015-06-26刘玉英李卓琳韩佳育穆春生
刘玉英,李卓琳,韩佳育,穆春生*
(1.吉林省气候中心,吉林长春130062;2.东北师范大学草地科学研究所,植被生态科学教育部重点实验室,吉林长春130024)
模拟降雨量变化与CO2浓度升高对羊草光合特性和生物量的影响
刘玉英1,李卓琳2,韩佳育2,穆春生2*
(1.吉林省气候中心,吉林长春130062;2.东北师范大学草地科学研究所,植被生态科学教育部重点实验室,吉林长春130024)
羊草作为欧亚大陆草原东缘干旱半干旱区域的优势种,现生产力已逐年降低。为明确未来降雨量变化和CO2浓度升高对羊草草地变化的影响,我们模拟3个降雨量梯度(正常降雨量40%)和两个CO2浓度(380±20,760±20μmol/mol)进行了光合特性与生物量的研究。结果表明,降雨量变化和CO2浓度的交互作用显著影响羊草的净光合速率等光合特性,地下生物量和根冠比,而不影响地上生物量。在目前CO2浓度时随着降雨量的增加,以及在CO2浓度升高时的降雨量由低降雨量增至正常降雨量,羊草的净光合速率、气孔导度、蒸腾速率和地上、地下及总生物量均显著增加,但在CO2浓度升高,降雨量由正常降雨量增至高降雨量时上述指标无显著变化。根冠比在目前CO2浓度时差异不显著,在CO2浓度升高下低降雨量时显著增加。在降雨量相同条件下,CO2浓度的升高使胞间CO2浓度、水分利用效率显著增加,气孔导度和蒸腾速率显著降低。低降雨量时CO2浓度升高对羊草净光合速率和生物量的促进作用显著高于正常降雨量和高降雨量,而高降雨量时CO2浓度升高对羊草生物量没有促进作用。以上结果暗示着在目前CO2浓度下,随着降雨量的增加,羊草生物量随之增加,在未来CO2浓度升高的背景下,高降雨量年份对生物量的积累并无显著的促进作用,CO2浓度升高可以补偿低水分条件对多年生根茎型禾草——羊草生长发育所造成的不利影响。
CO2浓度;降雨量;羊草;生物量;光合特性
近几十年来,人类活动及现代工业的迅速发展导致大气CO2浓度逐步上升,已由工业革命前期的270 μmol/mol增加到现在的380μmol/mol,并预计到本世纪末将倍增[1]。CO2浓度的升高会影响植物的生长发育和生理特性等[2-4]。一般CO2浓度升高会加强植物的光合作用[5-6],增加C吸收,进而促进生物量累积[7-8]。在草原生态系统,已有报道显示CO2浓度的升高均促进瑞士草原,新西兰草原,堪萨斯高草草原以及科罗拉多矮草草原的生产力[5]。但是不同光合途径的植物对CO2的响应不同,例如C3植物对CO2浓度升高的刺激作用更为敏感,而C4植物由于其独有的光合结构,现在的大气CO2水平可以满足其光合作用的需要,所以CO2浓度的增加对C4植物的影响较小[9-10]。还有部分研究表明CO2对植物光合作用的促进作用一直持续且并没有发生光合下调现象,但也有相反的研究结果[11-12]。CO2的间接作用主要是通过增加植物的水分利用效率以及加强土壤的持水能力进而影响植物的生长发育[13-14]。CO2浓度变化对生物量分配影响的研究结果则大不相同[15-17]。
温室效应引起的气候变化可能会加重中国半干旱区域的干旱化[18-19]。在干旱与半干旱地区,因其较低的年降水量,该地区的生态系统主要受水分的限制和驱动,因此水分是制约植被生产力和植被恢复的关键因子[20]。降雨量的增加会促进植被生产力,而降雨量的减少则会产生相反的作用[21-22]。在轻度乃至中度干旱的条件下,植物会通过降低气孔导度来减少蒸腾,但同时也降低了光合速率[7],而严重的干旱会损坏植株的光合系统,降低光合能力。一般在降雨量低的地区,植物会产生较小的叶面积和植株个体,通过发达的根系来吸收养分和水分维持植物的生长,保持植物个体适合度最高的生长格局[15-17]。
在干旱与半干旱地区的草地生态系统,CO2对植被的影响一般也会受到主要限制因子水分的调控,所以降雨量变化和CO2浓度升高的交互作用对植物生长发育的影响更为重要。现有新的证据表明,CO2浓度升高在干旱条件下对植物生长的刺激作用要大于在水分充足的条件[5,23-24],然而也有学者认为干旱胁迫削弱CO2对植物生长的刺激作用[25-27]。这种结果的差异可能是由于不同的物种,发育阶段以及环境因子所致。羊草 (Leymus chinensis)作为欧亚草原东缘的优势种,是一种典型的以根茎繁殖为主的C3多年生禾本科牧草[28],不但具有较强的抗旱和耐盐碱能力,而且还具有较好的营养价值及适口性[29]。由于多年来草地管理不当以及气候变化等原因,羊草草地生态系统已逐步退化,以其为优势种的草地生产力也随之降低。目前尽管已有文献报道了CO2浓度升高或干旱对一些C3植物光合特性和生物量累积的影响,但对羊草的研究仍然较少。因此,本实验以羊草为研究对象,探讨模拟降雨量变化(正常降雨量及±40%)和CO2浓度升高(380±20,760±20μmol/mol)对羊草光合特性和生物量累积的影响,旨在探讨在未来降雨量变化及在CO2浓度升高背景下,羊草的适应对策和变化趋势。
1 材料与方法
1.1 研究区域概况
研究区位于中国东北部松嫩草原西南部的吉林省(44°44′N,123°44′E,海拔167 m)。该地区属温带半干旱大陆性季风气候,无霜期140 d左右。年平均气温为6.4℃,年均降水量为361.6 mm(2000-2011年),大部分集中在6-8月份,该阶段的降雨量约占全年降水量的70%以上。该地主要土壤类型为盐碱化草甸土。
1.2 实验材料
供试羊草种子和土壤于2011年7月下旬采自松嫩草地西南部的吉林省长岭县,东北师范大学草业科学定位研究站的人工羊草草地。将所用土壤(草甸盐碱土)过筛去除杂物,充分混合后,装入24个内径为19 cm、高14 cm的花盆中(每盆3.3 kg)。土壤总氮和有机质含量分别为6.8%和0.3%,p H为8.63。为了获得足够且长势均一的幼苗,2012年6月1日将20粒羊草种子均匀地播种在每个花盆中(共24个花盆,480粒种子)。
播种完后将所有花盆放入东北师范大学的大型人工气候室(LT/ACR-2002 Phytotron System,E-Sheng Tech.,Beijing,China),气候室内的光合有效辐射强度为350μmol/(m2·s),光源采用飞利浦高压生物钠灯,光照时间14 h,黑暗时间10 h。气候室内的温度设置为:5:30-8:30,22℃;8:30-11:30,25℃;11:30-14:30,28℃;14:30-17:30,25℃;17:30-19:30,22℃;19:30-5:30,18℃。期间每3 d补水一次(8 mm降雨量,每月共80 mm;相当于维持土壤田间持水量的50%~60%)。其中,白天/夜间时长的选择主要是依据松嫩草地6-8月份的昼夜时长变化而定。温度和降雨量的变化是根据2000-2011年松嫩草地夏季6-8月份的气象数据模拟所得(图1,图2)。待播种15 d后进行间定苗,每盆留苗10株。
1.3 模拟降雨量变化和CO2浓度升高设置
在羊草种子播种一个月后开始进行模拟降雨量变化和CO2浓度升高,处理时间为3个月。在处理初期,将24个花盆随机均分到两个气候室,一个气候室作为对照,室内的CO2浓度24 h维持在(380±20)μmol/mol。另一个气候室作为CO2倍增处理,CO2气体是由液体CO2钢瓶提供,并通过直径为0.64 cm的塑料管向室内输送,使室内的CO2浓度维持在(760±20)μmol/mol。气候室内的CO2浓度是由电脑每1 s监测一次,若低于目标值,每5 s向室内输送一次CO2气体。
在每一个CO2浓度水平,模拟3个降雨量梯度:分别是正常降雨量(precipitation,P)、降雨量增加40%(高降雨量,high precipitation,HP)和降雨量减少40%(低降雨量,low precipitation,LP)。每月降雨量平均分为10次补给,即每3 d补水一次,每次浇水量折合为8 mm降雨量。这是依据松嫩草地(吉林省长岭县)2000-2011年夏季(6-8月份)的平均降雨量、最高和最低降雨量,以及降雨频次(图2)而定。通过对松嫩草地过去12年降水量的分析表明,降水最丰年的生长季内降水量比多年平均水平高出约40%,而降水最欠年基本比多年平均水平低约40%,因而模拟实验期间以总降雨量增加或减少40%为处理。
实验处理过程中,为了消除两个气候室内光照等因素的干扰,每3 d随机置换一次花盆的位置。此外,我们每半个月置换两个气候室内的花盆,同时更改CO2处理的设置以尽量减少气候室差异造成的系统误差。
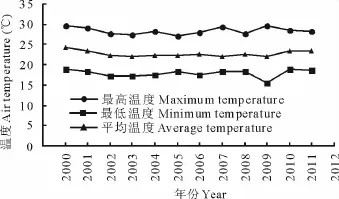
图1 2000-2011年松嫩草地夏季6-8月份的最高、最低及平均气温变化Fig.1 The average maximum,minimum and mean air temperatures on the semi-arid Songnen Grassland during summers(Jun-Aug)from 2000 to 2011
1.4 气体交换参数测定
在处理3个月时,测定生物量之前,我们从不同处理的不同花盆中随机选取最上完全展开的叶片12片(每盆3片叶子),利用LI6400XT(Li-Cor,Inc.,Lincoln,NE,USA)便携式光合测定仪连接LED红蓝光源测定羊草叶片的气体交换参数,LED光源设置成与室内自然光照相同的强度350μmol/(m2·s),二氧化碳浓度分别设置为380和760μmol/mol,测定羊草的净光合速率(photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间二氧化碳浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,E),最后通过计算净光合速率和蒸腾速率的比值得出水分利用效率(water use efficiency,WUE)。
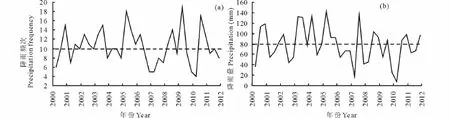
图2 2000-2011年松嫩草地夏季6-8月份的降雨量及降雨频次变化Fig.2 The change of precipitation frequency and precipitation in summer(Jun-Aug)from 2000 to 2011
2 结果与分析
2.1 降雨量变化与CO2浓度升高对土壤含水量的影响
降雨量变化和CO2浓度升高对土壤含水量并没有显著的交互作用。土壤含水量随着降雨量的增加而显著增加,CO2浓度升高轻微提高土壤含水量,但总体并未达到显著水平(P=0.074)(表1,图3)。
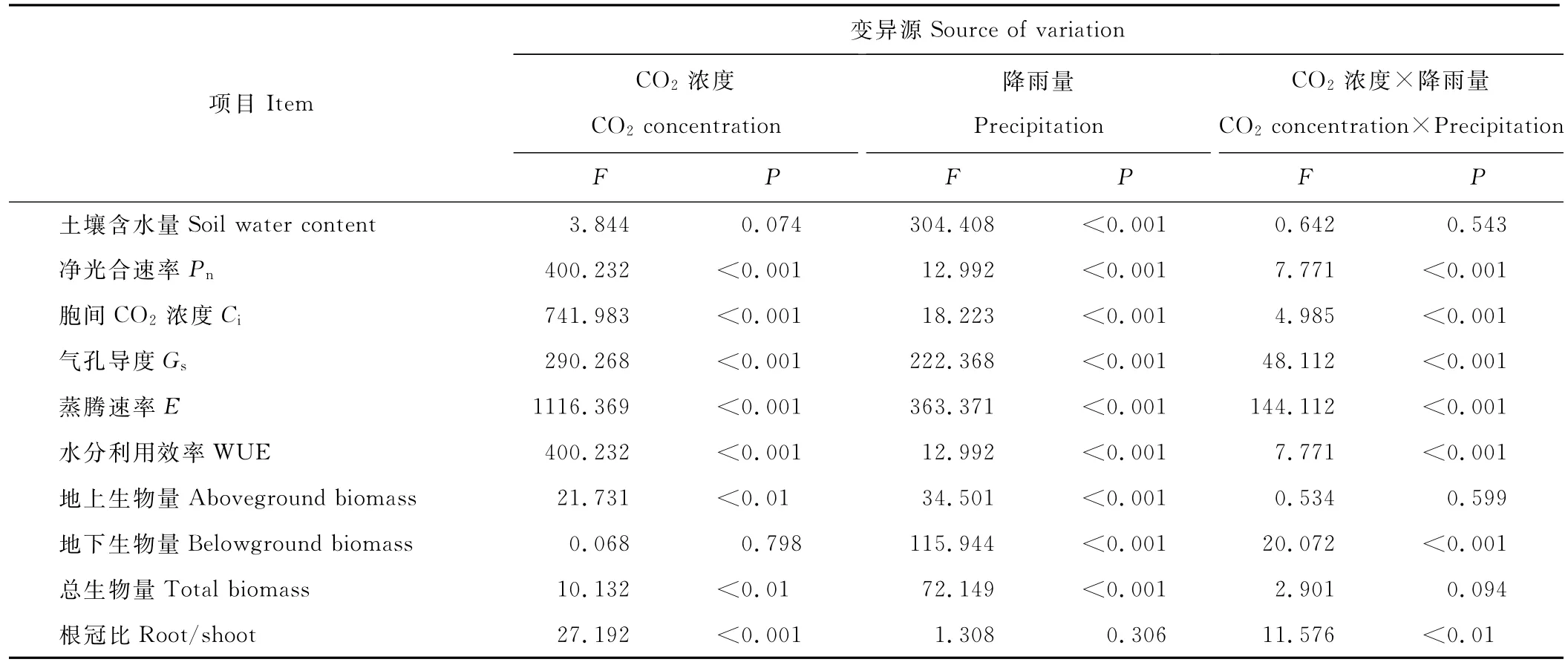
表1 降雨量变化和CO2浓度升高对土壤含水量、羊草光合特性、生物量及分配的影响Table 1 Effect of elevated CO2,precipitation on soil water content,photosynthetic characteristic and biomass of L.chinensis
2.2 降雨量变化与CO2浓度升高对羊草光合特性的影响
降雨量变化和CO2浓度升高均显著影响羊草的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(E)和水分利用效率(WUE)(表1)。由表2可知,在目前CO2水平下,随着降雨量的增加,羊草的净光合速率、气孔导度和蒸腾速率均显著增加,且蒸腾速率增长的幅度大于净光合速率,导致水分利用效率在高降雨量时比低降雨量和正常降雨量条件下的显著减少(P<0.001)。同时,在CO2浓度为760μmol/mol时,羊草的光合特性基本和CO2浓度为380μmol/mol时相似,但高降雨量条件下的净光合速率虽然比正常降雨量条件下高,但并未达到显著水平,水分利用效率也随着降雨量的增加显著降低。
与目前CO2水平相比,CO2浓度升高促使羊草的净光合速率在3个降雨量梯度(从低到高)分别增加了82.4%,36.7%和14.0%,在低降雨量条件下的促进作用最强,在高降雨量条件下最弱且未达到显著水平(P>0.05)。胞间CO2浓度和净光合速率的趋势基本一致;而气孔导度在3个降雨量梯度(从低到高)分别比目前CO2水平极显著减少了40.0%,50.0%和58.8%(P<0.001),蒸腾速率在3个降雨量梯度(从低到高)极显著减少了60.5%,65.6%和71.9%,导致在高CO2水平时,水分利用效率在3个降雨量梯度(从低到高)分别显著增加了398.3%,307.3%和308.0%(P<0.001)。在低降雨量时水分利用效率最高,这表明CO2浓度升高尤其是在干旱条件下对羊草净光合速率和水分利用效率的促进作用最大,补偿干旱对羊草造成的负面影响。
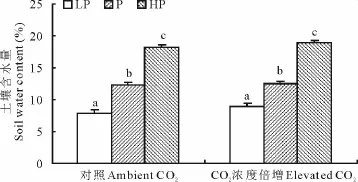
图3 不同CO2浓度条件下降雨量变化对土壤含水量的影响Fig.3 Effect of precipitation on soil water content under two CO2concentration

注:不同小写字母代表不同降雨量梯度间差异显著(P<0.05),不同大写字母代表两个CO2浓度间差异显著(P<0.05)。下同。Note:Different lower-case letters indicate significant differences(P<0.05)among precipitation levels and different capital letters indicate significant differences between two CO2levels(P<0.05).The same below.
2.3 降雨量变化和CO2浓度升高对羊草生物量积累的影响
降雨量变化显著影响羊草的地上、地下和总生物量,CO2浓度升高显著影响羊草的地上生物量、总生物量和根冠比,两者的交互作用只显著影响羊草的地下生物量和根冠比(表1)。由表3可知,在目前CO2水平,羊草单株的地上生物量、地下生物量及总生物量均随着降雨量的增加而显著增加(P<0.05)。而在CO2浓度升高的背景下,当降雨量由低水平到正常水平时,地上生物量、地下生物量及总生物量显著增加,根冠比显著减少。而当降雨量由正常水平增加到高水平时,地上生物量、地下生物量及总生物量并没有增加,根冠比也差异不显著,表明CO2浓度升高和降雨量增加对地下生物量和根冠比产生拮抗效应。
与CO2为380μmol/mol相比,CO2浓度升高在3个降雨量梯度(从低到高)促使羊草的地上生物量分别增加了29.6%,32.6%和22.9%,其中只有在正常降雨量时达到显著水平(P<0.05)。CO2浓度升高显著增加低降雨量时的羊草地下生物量,而显著降低高降雨量时的羊草地下生物量(P<0.05)。因此,CO2浓度升高在低降雨量时显著增加羊草的总生物量,而在高降雨量时对羊草的总生物量没有影响,并且只有在正常降雨量时对地上生物量的促进作用是显著的。此外,在正常降雨量和高降雨量时,CO2浓度升高也显著减少了羊草的根冠比(P<0.05),这表明CO2浓度升高在水分适宜的条件下会促进羊草地上植被的生长,而在干旱时会增加根系的分配而吸收水分。
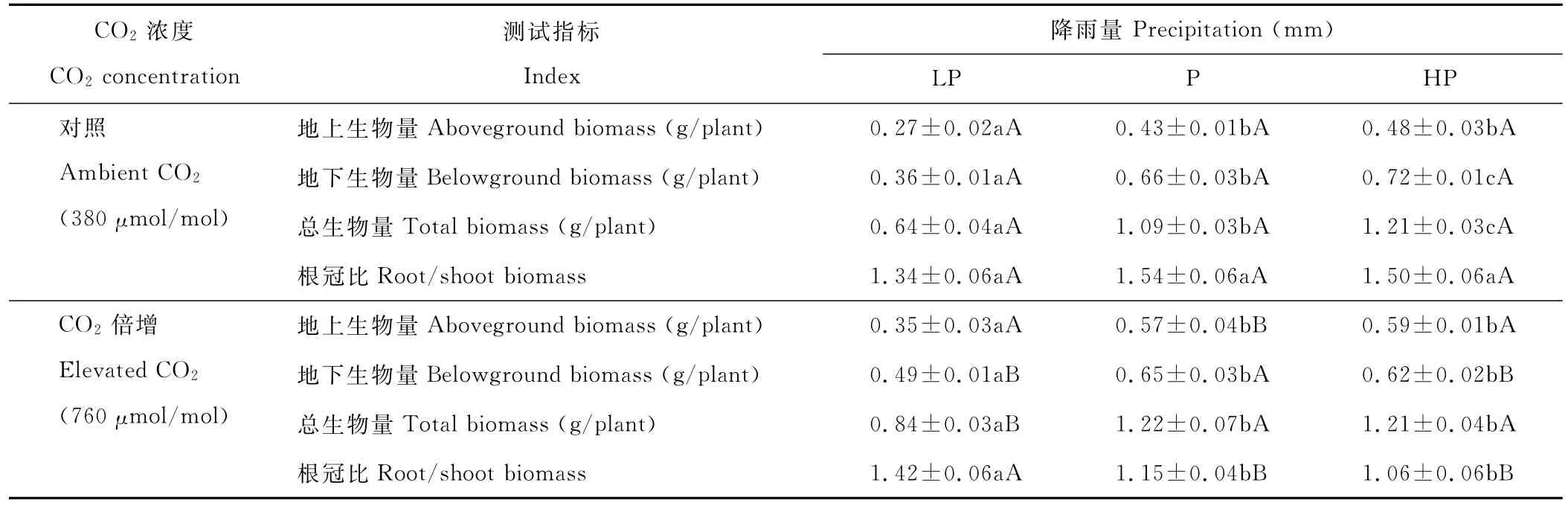
表3 不同CO2浓度条件下降雨量变化对羊草生物量积累的影响Table 3 Effect of precipitation on biomass of L.chinensis under two CO2concentration
3 讨论
在干旱和半干旱地区,水分是调控植物生长的主要驱动因子,它可以影响植物的物质代谢和信号传递,进而影响植株的生长发育。CO2浓度升高作为全球气候变化的基础因子,与单独研究降水相比,研究CO2浓度升高和降水量变化的交互作用更有意义。目前大部分研究显示,CO2浓度升高增加羊草的光合速率进而促进其生物量累积,而降雨量的减少则呈现相反的趋势[7,21,23-24,30-32],而对其交互作用的研究结果各不相同[3,33-34]。
3.1 CO2浓度升高条件下降雨量的过多增加对羊草生物量的累积和净光合速率并无显著的促进作用
羊草具有较强的耐旱性,在松嫩草地夏季(6-8月)羊草生长地的土壤田间持水量一般为50%~60%,所以较高的降雨量对羊草生长的促进作用未必显著。在本研究中的高降雨量处理组,较高的土壤含水量使土壤一直处于湿润状态,计算其田间持水量达到或超过80.0%,在CO2浓度为380μmol/mol时,高降雨量条件下的羊草总生物量仅比正常降雨量条件下的增加了11.0%,而在CO2浓度为760μmol/mol时,并没有显著差异(P>0.05)(表3),同时CO2浓度升高和降雨量增加也没有表现出协同作用(表1),这表明CO2浓度升高在高降雨量时并未对羊草的总生物量起到促进作用。Xu和Zhou[35]研究水分胁迫和夜间高温对羊草影响的结果显示对照组(田间持水量为75%~80%),轻度胁迫(田间持水量为60%~65%)和中度胁迫(田间持水量为50%~55%)对羊草的生物量影响不显著,而且轻度胁迫反而会刺激羊草的生物量累积,这与我们的研究结果基本一致。新疆大叶苜蓿(Medicago sativa)作为一种耐旱植物,有学者研究其应对较高的土壤含水量时也得到类似的结果[36]。净光合速率的变化与生物量的变化呈现相似的趋势,在CO2浓度为380μmol/mol时,随着降雨量的增加净光合速率显著增加,而在CO2浓度为760μmol/mol时,正常降雨量和高降雨量条件下的净光合速率差异不显著(表2)。这也表明CO2浓度升高在高降雨量时对羊草叶片光合作用的促进作用非常微弱,这可能是羊草长期生长在半干旱地区所产生的适应对策。以上结果暗示由于羊草对较干旱环境的长期适应,在高CO2浓度的背景下,降雨量过多的增加对羊草的生物量累积和净光合速率并无显著的促进作用。
3.2 CO2浓度升高会部分补偿干旱对羊草生物量和净光合速率造成的负面影响
在我们的研究中,与目前CO2水平相比,CO2浓度升高在低降雨量时对羊草的净光合速率影响最大(表2)。这与前人的研究结果相同[5,9,14,37]。在Poorter和Pérez[23]的研究中,13个草本植物在干旱条件下与水分适宜条件下的相比,CO2的刺激作用要更强烈,这也支持我们的研究结果。CO2浓度升高在低降雨量时气孔导度减少最小(表2),这可能是干旱已经降低植物的气孔导度以减少蒸腾作用,CO2浓度升高在此基础上减少的最小,所以增加底物浓度仍会促进植物的光合速率。而在高降雨量时即使底物浓度增加,但气孔导度迅速降低来阻碍CO2的进入,所以对植物的净光合速率影响不显著。一般来说,在干旱半干旱地区的草地生态系统,较低的年均降雨量导致植被的水分利用效率成为刺激植被生产力的重要因子之一[38]。在本研究中,CO2浓度升高极显著地提高了羊草的水分利用效率(表1,表2),并且在低降雨量的条件下轻微地增加了土壤含水量,虽然未有显著差异(表1,图3)。前人的研究也得出,CO2浓度升高条件下的土壤含水量要高于目前CO2水平,尤其是在降雨量少到限制植物正常生长的时期[39]。CO2升高1倍,水分利用效率提高3~4倍,也就是说固定单位CO2所需的水要少3~4倍。这在低降雨量条件下尤为重要,所以CO2对植物生长的促进作用即使在低降雨量的条件下受到一些限制,但仍可以部分补偿干旱所造成的负面影响。
净光合速率的增加暗示着羊草加强了对C的固定,CO2浓度升高分别促使羊草单株的总生物量在3个降雨量梯度(从低到高)增加了31.3%,11.9%和0.0%,这与羊草净光合速率变化趋势一致(表3)。此外在低降雨量时增加了羊草的根冠比,而在正常降雨量和高降雨量时减少羊草的根冠比,这一结果支持了前人的研究结论[9,26-27]。这可能是由于在低降雨量时,即使CO2的作用会增加羊草的水分利用效率,增加土壤的保水能力,但仍使羊草受到干旱胁迫,所以增加地下生物量吸收更多的养分和水分来供应植株地上部分的生长,根冠比的增加是植物应对干旱胁迫的生长对策[35]。虽然还有研究表明CO2浓度升高在水分良好的条件下对植株生长的刺激作用要高于在干旱条件[25-27],这种结果的差异可能是由于不同植物本身的抗旱能力以及干旱尺度等原因。因此在低降雨量且CO2浓度升高的背景下,羊草的生长和生物量的积累是CO2作为光合底物促进羊草的C吸收和促进植株水分利用效率的共同作用。这暗示CO2浓度升高可以部分补偿未来降雨量轻微减少对羊草生长的负面影响。
简而言之,本研究结果表明,在目前CO2浓度下,随着降雨量的增加,羊草净光合速率和生物量随之增加,但在CO2浓度升高条件下,当降雨量增加到一定程度后对羊草净光合速率和生物量的积累并无显著的促进作用,并减少根系的投入,CO2浓度升高可以补偿低水分条件对多年生根茎型禾草——羊草生长发育所造成的不利影响。
[1] Houghton J T,Ding Y,Griggs D J,et al.Climate change 2001:the scientific basis[R].Cambridge:Cambridge University Press,2001.
[2] Centritto M,Helen S,Lee J,et al.Interactive effects of elevated[CO2]and drought on cherry(Prunus avium)seedlings I.Growth,whole-plant water use efficiency and water loss.New Phytologist,1999,141:129-140.
[3] Bhattacharya N,Hileman D,Ghosh P,et al.Interaction of enriched CO2and water stress on the physiology of and biomass production in sweet potato grown in open-top chambers.Plant Cell Environment,2006,13:933-940.
[4] Ge Z M,Zhou X,Kellomäki S,et al.Responses of leaf photosynthesis,pigments and chlorophyll fluorescence within canopy position in a boreal grass(Phalarisarundinacea L.)to elevated temperature and CO2under varying water regimes.Photosynthetica,2011,49:172-184.
[5] Morgan J A,LeCain D R,Pendall E,et al.C4grasses prosper as carbon dioxide eliminates desiccation in warmed semi-arid grassland.Nature,2011,476:202-205.
[6] Kinmonth S H,Kim S H.Carbon gain,allocation and storage in rhizomes in response to elevated atmospheric carbon dioxideand nutrient supply in a perennial C3grass,Phalaris arundinacea.Functional Plant Biology,2011,38:797-807.
[7] Jackson R,Luo Y,Cardon Z,et al.Photosynthesis,growth and density for the dominant species in a CO2-enriched grassland.Journal of Biogeography,1995,22:1225-1229.
[8] Cheng W,Sakai H,Yagi K,et al.Interactions of elevated[CO2]and night temperature on rice growth and yield.Agricultural and Forest Meteorology,2009,149:51-58.
[9] Mishra R,Abdin M,Uprety D.Interactive effects of elevated CO2and moisture stress on the photosynthesis,water relation and growth of Brassica species.Journal of Agronomy and Crop Science,1999,182:223-230.
[10] Kakani V,Reddy K R.Temperature response of C4species big bluestem(Andropogon gerardii)is modified by growing carbon dioxide concentration.Environmental and Experimental Botany,2007,61:281-290.
[11] Ellsworth D S,Thomas R,Crous K Y,et al.Elevated CO2affects photosynthetic responses in canopy pine and subcanopy deciduous trees over 10 years:a synthesis from Duke FACE.Global Change Biology,2012,18:223-242.
[12] Komatsu M,Tobita H,Watanabe M,et al.Photosynthetic downregulation in leaves of the Japanese white birch grown under elevated CO2concentration does not change their temperature-dependent susceptibility to photoinhibition.Physiologia Plantarum,2012,147:159-168.
[13] Drake B G,Gonzàlez M A,Long S P.More efficient plants:a consequence of rising atmospheric CO2.Annual Review of Plant Biology,1997,48:609-639.
[14] Wang H,Zhou G S,Jiang Y L,et al.Interactive effects of changing precipitation and elevated CO2concentration on photosynthetic parameters of Stipa breviflora.Chinese Journal of Plant Ecology,2012,36(7):597-606.
[15] Wand S J,Midgley G,Jones M H,et al.Responses of wild C4and C3grass(Poaceae)species to elevated atmospheric CO2concentration:a meta-analytic test of current theories and perceptions.Global Change Biology,1999,5:723-741.
[16] Poorter H,Nagel O.The role of biomass allocation in the growth response of plants to different levels of light,CO2,nutrients and water:a quantitative review.Australian Journal of plant physiology,2000,27:595-607.
[17] Poorter H,Niklas K J,Reich P B,et al.Biomass allocation to leaves,stems and roots:meta-analyses of interspecific variation and environmental control.New Phytologist,2012,193:30-50.
[18] Li X Z,Liu X D,Ma Z G.Analysis on the drought characteristics in the main arid regions in the world since recent hundredodd years.Arid Zone Research,2004,21(2):97-103.
[19] Ainsworth E A,Rogers A.The response of photosynthesis and stomatal conductance to rising[CO2]:mechanisms and environmental interactions.Plant Cell Environment,2007,30:258-270.
[20] Rao L E,Allen E B.Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts.Oecologia,2010,162:1035-1046.
[21] Flexas J,Medrano H.Drought-inhibition of photosynthesis in C3plants:stomatal and non-stomatal limitations revisited.Annals of Botany,2002,89:183-189.
[22] Báez S,Collins S L,Pockman W T,et al.Effects of experimental rainfall manipulations on Chihuahuan Desert grassland and shrubland plant communities.Oecologia,2013,172:1117-1127.
[23] Poorter H,Perez M.The growth response of plants to elevated CO2under non-optimal environmental conditions.Oecologia,2001,129:1-20.
[24] Atwell B J,Henery M L,Rogers G S,et al.Canopy development and hydraulic function in Eucalyptus tereticornis grown in drought in CO2-enriched atmospheres.Functional Plant Biology,2007,34:1137-1149.
[25] Smith S D,Huxman T E,Zitzer S F,et al.Elevated CO2increases productivity and invasive species success in an arid ecosystem.Nature,2000,408:79-82.
[26] Erice G,Irigoyen J J,Sánchez M,et al.Effect of drought,elevated CO2and temperature on accumulation of N and vegetative storage proteins(VSP)in taproot of nodulated alfalfa before and after cutting.Plant Science,2007,172:903-912.
[27] Xu Z,Zhou G,Wang Y.Combined effects of elevated CO2and soil drought on carbon and nitrogen allocation of the desert shrub Caragana intermedia.Plant and Soil,2007,301:87-97.
[28] Wang Y,Zhou G,Wang Y.Modeling responses of the meadow steppe dominated by Leymus chinensis to climate change.Climatic Change,2007,82:437-452.
[29] Huang Z H,Zhu J M,Mu X J,et al.Advances on the mechanism of low sexual reproductivity of Leymus chinensis.Grassland of China,2002,24(6):55-60.
[30] Ainsworth E A,Long S P.What have we learned from 15 years of free-air CO2enrichment(FACE)?A meta-analytic review of the responses of photosynthesis,canopy properties and plant production to rising CO2.New Phytologist,2005,165:351-372.
[31] Ziska L,Sicher R,George K,et al.Rising atmospheric carbon dioxide and potential impacts on the growth and toxicity ofpoison ivy(Toxicodendron radicans).Weed Science,2007,55:288-292.
[32] Xu Z,Shimizu H,Yagasaki Y,et al.Interactive effects of elevated CO2,drought,and warming on plants.Journal of Plant Growth Regulation,2013,32:692-707.
[33] Tolley L C,Strain B.Effects of CO2enrichment and water stress on growth of Liquidambar styraciflua and Pinus taeda seedlings.Canadian Journal of Botany,1984,62:2135-2139.
[34] Ottman M,Kimball B,Pinter P,et al.Elevated CO2increases sorghum biomass under drought conditions.New Phytologist,2001,150:261-273.
[35] Xu Z Z,Zhou G S.Effects of water stress and high nocturnal temperature on photosynthesis and nitrogen level of a perennial grass Leymus chinensis.Plant and Soil,2005,269:131-139.
[36] Luo Y Z,Li G.The effect of water stress on growth and biomass of Medicago sativa cv.Xinjiangdaye.Acta Prataculturae Sinica,2014,4:213-219.
[37] Fay P A,Polley H W,Jin V L,et al.Productivity of well-watered Panicum virgatum does not increase with CO2enrichment.Journal of Plant Ecology,2012,5:366-375.
[38] Niu S,Xing X,Zhang Z,et al.Water-use efficiency in response to climate change:from leaf to ecosystem in a temperate steppe.Global Change Biology,2011,17:1073-1082.
[39] Morgan J,Newton P,Nösberger J,et al.The influence of rising atmospheric CO2on grassland ecosystems.Proceedings of the XIX International Grasslands Congress,2001,200(1):973-980.
参考文献:
[14] 王慧,周广胜,蒋延玲,等.降水与CO2浓度协同作用对短花针茅光合特性的影响.植物生态学报,2012,36(7):597-606.
[18] 李新周,刘晓东,马柱国,等.近百年来全球主要干旱区的干旱化特征分析.干旱区研究,2004,21(2):97-103.
[29] 黄泽豪,朱锦懋,母锡金,等.羊草有性繁殖力低的成因研究进展.中国草地,2002,24(6):55-60.
[36] 罗永忠,李广.土壤水分胁迫对新疆大叶苜蓿的生长及生物量的影响.草业学报,2014,23(4):213-219.
Influences of precipitation regimes and elevated CO2on photosynthesis and biomass accumulation in Leymus chinensis
LIU Yu-Ying1,LI Zhuo-Lin2,HAN Jia-Yu2,MU Chun-Sheng2*
1.Climate Center of Jilin Province,Changchun 130062,China;2.Institute of Grassland Science,Northeast Normal University,Key Laboratory of Vegetation Ecology,Ministry of Education,Changchun 130024,China
Leymus chinensis is a dominant,rhizomatous perennial C3species found in the grasslands of Songnen Plain,Northern China,where its productivity has decreased year by year due to environmental changes.As a dominant species growing in arid and semiarid regions,precipitation is a key factor limiting plant productivity.To determine how this species’productivity responds to different precipitation regimes and increased CO2levels,we conducted an experiment that measured photosynthetic parameters,along with the accumulation and partitioning of biomass.Plants were subjected to combinations of three precipitation gradients(normal precipi-tation,normal±40%)and two CO2levels(380±20 and 760±20μmol/mol)in environment-controlled chambers.The interaction between increased CO2and precipitation had significant effects on photosynthetic parameters,belowground biomass and root∶shoot ratio,but no effect on aboveground biomass.Net photosynthetic rate,stomatal conductance,transpiration rate,aboveground and total biomass rose with increases in precipitation and CO2concentration,but no significant change was observed when precipitation increased from normal to high under CO2enrichment.There was no significant difference in the ratio of root to shoot among precipitation regimes at the low CO2condition,but it changed significantly with CO2enrichment and low precipitation.Water use efficiency and intercellular CO2concentration increased significantly when precipitation was low,but decreased when precipitation was high under CO2enrichment.The effect of elevated CO2on photosynthesis and biomass accumulation was more obvious at low precipitation than at normal or high precipitation.The results suggest that at ambient CO2levels,net photosynthetic rate and biomass of L.chinensis increase with precipitation,but that these measures are not further affected by additional precipitation when CO2is elevated.Furthermore,CO2may partly compensate for the negative effect of low precipitation on the growth and development of L.chinensis.
elevated CO2;precipitation regime;Leymus chinensis;biomass accumulation;photosynthesis
10.11686/cyxb2014474 http://cyxb.lzu.edu.cn
刘玉英,李卓琳,韩佳育,穆春生.模拟降雨量变化与CO2浓度升高对羊草光合特性和生物量的影响.草业学报,2015,24(11):128-136.
LIU Yu-Ying,LI Zhuo-Lin,HAN Jia-Yu,MU Chun-Sheng.Influences of precipitation regimes and elevated CO2on photosynthesis and biomass accumulation in Leymus chinensis.Acta Prataculturae Sinica,2015,24(11):128-136.
2014-01-19;改回日期:2015-01-14
973课题(2015CB150801),国家自然科学基金(31172259,31370432,31372369)和高等学校博士学科点专项科研基金(201200 43110010)资助。
刘玉英(1963-),女,吉林长春人,高级工程师。E-mail:ccliuyuying@126.com
*通讯作者Corresponding author.E-mail:mucs821@nenu.edu.cn
