科尔沁草甸生态系统净碳交换特征及其驱动因子
2015-06-26刘廷玺雷慧闽张圣微段利民
王 婧,刘廷玺*,雷慧闽,张圣微,段利民
(1.内蒙古农业大学水利与土木建筑工程学院,内蒙古呼和浩特010018;2.清华大学水利水电工程系,北京100084)
科尔沁草甸生态系统净碳交换特征及其驱动因子
王 婧1,刘廷玺1*,雷慧闽2*,张圣微1,段利民1
(1.内蒙古农业大学水利与土木建筑工程学院,内蒙古呼和浩特010018;2.清华大学水利水电工程系,北京100084)
以2013年10月-2014年9月连续观测的CO2通量数据为基础,分析了科尔沁草甸生态系统净碳交换量(NEE)的时间变化特征及其驱动因素。结果表明,NEE日变化季节差异明显,生长季变化幅度大,净CO2日吸收速率7月>8月>9月>6月>5月;生长季内,NEE主要受控于叶面积指数和光合有效辐射。NEE与光合有效辐射(PAR)之间的关系可用直角双曲线方程来描述,拟合得到的表观初始光能利用率α为0.0015μmol CO2/μmol PAR,最大光合速率Pmax为0.65μmol CO2/(m2·s)。叶面积指数(LAI)对NEE的影响可由分段函数表示,当LAI>3.08时,表现为渐进饱和型,且LAI越大NEE对PAR的响应越明显;当高饱和水汽压差(VPD)在1.5~2.0 kPa时,光合作用开始降低,NEE明显受到VPD值的抑制;短暂强降雨(累计降雨量>40 mm/d)对昼间NEE有一定的抑制,而持续低强度降雨(降雨时长>15 h)对夜间NEE存在激发作用;夜间NEE随土壤温度呈指数增长,温度敏感系数(Q10)为2.63。
净生态系统交换量;驱动因子;涡度相关系统;科尔沁沙地
陆地生态系统的碳循环及其对环境因子的响应研究是理解生态系统碳平衡的关键。草地生态系统作为陆地生态系统的重要组成部分,几乎覆盖了陆地面积的20%,其净初级生产力约占全球陆地生物区净初级生产力的1/3。美欧科学家于1996年初步建立起全球CO2通量网,利用涡度技术在全球范围内开展了不同纬度、不同类型生态系统CO2通量的全球性研究,但其中70%是针对不同森林植被类型的,对草原生态系统的研究主要集中在低海拔的平原地区[1-2],且研究较少。Dugas等[2]对德高草原碳收支状况研究发现叶面积指数(LAI)、光合有效辐射(PAR)及土壤含水量影响碳通量大小,Shaver等[3]应用Pan-Arctic模型对生态系统净生态系统碳交换(以下简称“NEE”)研究表明,NEE的变异约75%可由LAI、气温(Ta)、PAR函数的单一回归模型解释。我国自20世纪90年代开始应用涡度相关法开展了陆面过程中碳循环通量的研究,尽管起步稍晚,但也取得了部分有价值的研究成果。伏玉玲等[4]研究了中国草地生态系统碳收支及其对环境的响应,郝彦宾等[5]研究了内蒙古羊草(Leymus chinensis)草原碳交换的季节变异规律,发现水分和PAR是影响碳交换的两个重要生态因子。Zhao等[6]研究了青藏高寒草甸净碳交换特征,表明叶面积指数(GLAI)、土壤含水量(θ)、Ta是3个对NEE季节变异影响最大的指标。
科尔沁沙地草甸区作为我国北方典型的半干旱草原区,具有极其脆弱的草原生态资源。与其他下垫面相比,对气候变化的响应更为敏感。近年来,有关该地区的碳通量研究主要局限于短期、不连续的观测数据[7]及碳排放的观测[8],针对NEE的研究比较缺乏。而研究半干旱区草甸生态系统NEE的日、季变化及其与气象因子的关系,对于了解碳的源汇过程、开展草原碳循环的模拟和气候变化情景下的预测研究具有十分重要的意义。本文依据涡度相关系统2013年10月-2014年9月的连续观测数据,对科尔沁草甸生态系统的碳收支及其过程机制进行定量评价,分析各气象要素对碳交换的影响,揭示草甸草原NEE的日、季变化规律,解析主要环境因子对碳源汇的影响及其差异,为评估该地区碳收支和明晰全球碳循环提供更为完善的试验数据和理论基础。
1 材料与方法
1.1 研究区概况
试验研究区隶属内蒙古通辽市科尔沁左翼后旗,面积约55 km2。该区多年平均气温约6.6℃,1月平均气温最低,为-13.3℃,7月平均气温最高,为23.8℃;多年平均降水量389 mm,降水主要集中在6-8月,占年总降水量的69.3%;多年平均蒸发量1412 mm,蒸发主要集中在4-9月。此间蒸发量占全年的17%。试验区为沙丘-草甸相间分布地区,草甸面积约占整个研究区面积的26.6%,本文选取草甸试验站(122°39′15″E,43°20′14″N)作为研究对象,该试验站地面相对高程约188.5 m,南临小型湖泊,地势平坦。天然植被主要有羊草、芦苇(Phragmites australis)等。6-8月份植被生长旺季,植被平均高度为145 cm,平均叶面积指数为2.47。
1.2 观测仪器与方法
草甸试验点主要安装有涡度相关系统传感器、微气象观测系统。分别在5,10,20,40,80,160 cm处埋有6层土壤温度、土壤含水量观测传感器。每30 d进行一次数据采集与处理,涡度原始数据与气象数据同步下载,本研究主要以2013年10月-2014年9月连续观测的CO2通量数据及微气象数据为基础。
1.2.1 涡度相关系统和微气象系统观测 涡度相关系统传感器安装在距地面2.6 m处,主要用于观测湍流通量。该系统主要包括三维超声风速仪(CSAT-3,Campbell Scientific,USA)及红外气体分析仪(LI-7500,Li-COR Inc,Nebraska,USA),分别用于测定三维风速与大气CO2和H2O密度,由数据采集器(CR3000,Campbell Scientific,USA)记录并存储系统同步计算的二氧化碳和水热通量,采样频率为10 Hz,数据记录时间间隔为30 min。
微气象观测系统主要包括安装在涡度观测塔上的四分量净辐射仪(NR-LITE,Campbell Scientific,USA)、光合有效辐射仪(LI190SB,Campbell Scientific,USA),以及3层(1.5,2.4,10.0 m)温湿度测量仪器(HMP45C,Campbell Scientific,US)、土壤热通量板(HFP01,Campbell Scientific,USA)、120 cm口径水面蒸发量(255,Nova Lynx,US)、降水量(TE525MM,Campbell Scientific,US)等。这些监测要素由数据采集器(CR1000,Campbell Scientific,USA)按10 min时间间隔计算其平均值并存储。
1.2.2 叶面积指数观测 对草甸群落叶面积指数LAI的动态测定也与碳通量观测同步进行,利用植物冠层分析仪(LAI-2200,Li-COR Nebraska,USA)通过测定冠层上方(简称“A”值)、冠层下方(简称“B”值)光照辐射的衰减来计算叶面积指数(叶面积指数=叶面积/地面积)。草甸植被生长初期的4-5月份,每隔15~20 d测量一次,6-9月份每20~30 d测量一次。选取3个1 m×1 m的样方,每个样方测定1个A值,4个B值,采用90度遮盖帽,取3组叶面积指数的平均值作为代表值。
1.3 数据处理
1.3.1 CO2通量的计算公式 CO2通量即净生态系统碳交换量,由CO2(μmol/m2·s)/H2O(mmol/m2·s)的浓度与垂直风速的原始数据经过协方差计算来确定。式中,w'为垂直湍流速度脉动,c'为二氧化碳浓度。NEE由涡度相关系统测定的CO2通量直接获得,NEE为正表示生态系统向大气释放CO2,反之则表示从大气吸收CO2。

1.3.2 通量数据的质量控制与插补 对涡动相关法观测到的10 Hz原始湍流数据运用Logger Net软件进行格式转换(TOB3-TOB1),然后采用美国LI-COR公司研发的Eddy Pro软件[9]进行系列校正计算,包括异常值及野值点剔除、坐标旋转修正、频率损失修正、超声虚温修正、空气密度效应修正(WPL修正)等基本处理,得到采样间隔为30 min的通量数据。
对30 min数据,若野点数大于100,则剔除该时次数据;或根据LI-7500红外气体分析仪诊断异常标志值(diag≠0)[10],以及相关监测数据超出CO2浓度阈值(100~1000 mg/m3),对夜间观测,用给定的一个临界阈值uc*加以检验。当u*<uc*时,则舍弃该通量数据不进行分析[11]。uc*值一般取0.1~0.2 m/s,本文取0.1 m/s。
对原始通量数据进行质量控制后,剔除的低质量数据会影响数据的连续性。因此,本研究对夜间和昼间缺失的NEE数据,采用分别的方法进行插补。针对夜间缺失的NEE数据,利用5 cm深度处的土壤温度(Ts)采用Vants Hoff拟合方程加以插补[12]。

式中,Reco为夜间生态系统呼吸速率(由于夜间没有光合作用,Reco即为NEE;Reco,ref为生态系统10℃时的参考呼吸速率(本文取0.05 mg/m2·s);Q10为呼吸随温度的变化速率(本文取2.63),即5至40℃之间,温度每增加10℃单位时间土壤呼吸增加的倍数,Raich和Schlesinger[13]通过收集和分析全球各种生态系统的土壤呼吸数据,发现Q10在1.3~3.3之间波动,Ts为5 cm深度处土壤实测温度。
针对昼间缺失的NEE数据,则利用Michaelis-Menten函数进行插补[13]。

式中,α为生态系统表观光量子效率;pmax为生态系统最大光合速率,即PAR→∞时NEE的渐进值;Reco,d为暗呼吸速率,即PAR→0时的NEE值。
2 结果与分析
2.1 净二氧化碳通量日内变化特征
草甸NEE的日变化基本呈现单峰型(图1),且呈现出上午碳吸收较强,下午碳吸收较弱的不对称现象,在日出(5:00-6:00时)、日落(18:00-19:00时)前后碳通量波动较大,此时的呼吸作用与光合作用达到一个平衡状态。夜间NEE为正,整个生态系统表现为碳排放,白天NEE均为负值,即随着光合作用的加强,整个生态系统表现为净碳吸收,5-9月日均碳吸收速率分别为0.02,0.12,0.33,0.29,0.24 mg/(m2·s),呈现为7月>8月>9月>6月>5月,且碳吸收出现在凌晨的时间依次滞后0.25,0.25,0.50,0.50 h,碳排放出现在傍晚的时间依次提前0.5,0.5,1.0,1.5 h。同时,日出、日落前后碳吸收量与排放量达到相对平衡的状态。

图1 生长季草甸生态系统碳通量月平均日变化Fig.1 Diurnal changes of CO2fluxes during growing season over the meadow grassland ecosystem
2.2 净二氧化碳通量及其影响因子的季节变化特征
为了解NEE与环境因子的季节变化特征,本文选取2013年10月-2014年9月生态系统NEE及其主要环境因子(饱和水汽压差VPD、气温Ta、土壤温度Ts、光合有效辐射PAR)的月变化加以分析(图2)。气温与5 cm深度处的土壤温度最大值均出现在7月中旬,分别为29.3和26.6℃,草甸生长季节的气温、昼夜温差、土壤温度的日平均值分别为19.3,10.0,16.0℃;VPD的日平均值为1.08 kPa,最高、最低值分别出现在7月9日和10月18日;PAR的月平均日变化呈现“单峰型”,最大值出现在7月5日,为1874μmol/(m2·s);8月降水量最高,为245.9 mm,7月次之,为104.6 mm。生长季平均叶面积指数LAI为2.57。4-6月LAI迅速增加,6-8月增速放缓,8月中下旬达到最大值3.34,9月微弱下降,但降幅较小。
NEE在全年表现出“两峰一谷”的特征(图2d),其中“峰值”出现在1,10月,均值分别为0.13和0.08 mg/(m2·s),“谷值”出现在7月,均值-0.33 mg/(m2·s),较徐丽君等[14]研究的贝加尔针茅草(Stipa baicalensis)原碳吸收谷值偏小。4月草甸NEE均值开始由正转负,生长季的5-9月,生态系统主要表现为碳汇,净CO2吸收总量1.02 mg/(m2·s),5,6,7,8,9月分别贡献了11.2%,27.0%,30.5%,22.5%,8.4%。但不同纬度的草甸生态系统NEE正负转换的时间有较大差异,如Shi等[15]研究嵩草草甸生态系统7-9月为碳吸收,其余为碳排放。而本研究区从10月出现碳排放,10月至翌年3月,净CO2日排放均值0.11 mg/(m2·s)。同时生态系统碳交换因受到环境因素影响而产生差异(表1),如生长季NEE与PAR、LAI相关性显著(P<0.01),非生长季NEE主要受控于温度因素(R2=0.568),生态系统主要表现为碳源。研究时段,草甸作为碳汇从大气中吸收218.52 g CO2/m2。
2.3 叶面积指数对NEE的影响
LAI是反映植物长势的重要特征参数,它对植物光能利用、干物质积累等都有明显影响。生长初期(LAI<0.97),由于植被稀疏LAI较小,对散射光的吸收较少,草甸CO2吸收量随LAI的增加而线性增长;生长旺期(0.97<LAI<3.08)植被生长稠密,LAI增加冠层有利于植物吸收来自各个方向的散射光,CO2吸收量随LAI的增加而缓慢增加,之后渐进饱和状态;生长末期(LAI>3.08)部分叶片开始枯黄、凋落,LAI有所下降,碳吸收减少且随LAI的增加而趋于平稳(图3),这与Suyker等[16]对玉米(Zea may)生殖生长研究得出的结论相似。同时由于生长末期群体底层存在大量枯叶,利用冠层分析仪进行测量时,仪器无法自动消除这部分枯叶的影响,导致LAI测定结果偏大[17]。
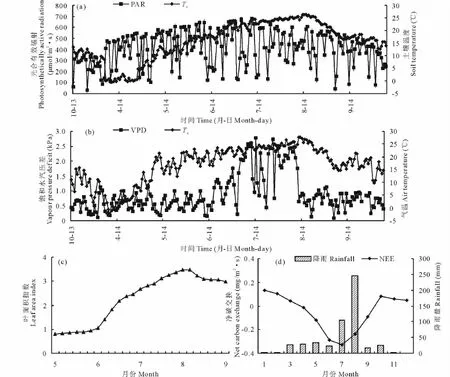
图2 净碳交换量NEE与环境因子的季节变化Fig.2 Seasonal variations of environmental factors with net carbon exchange(NEE)
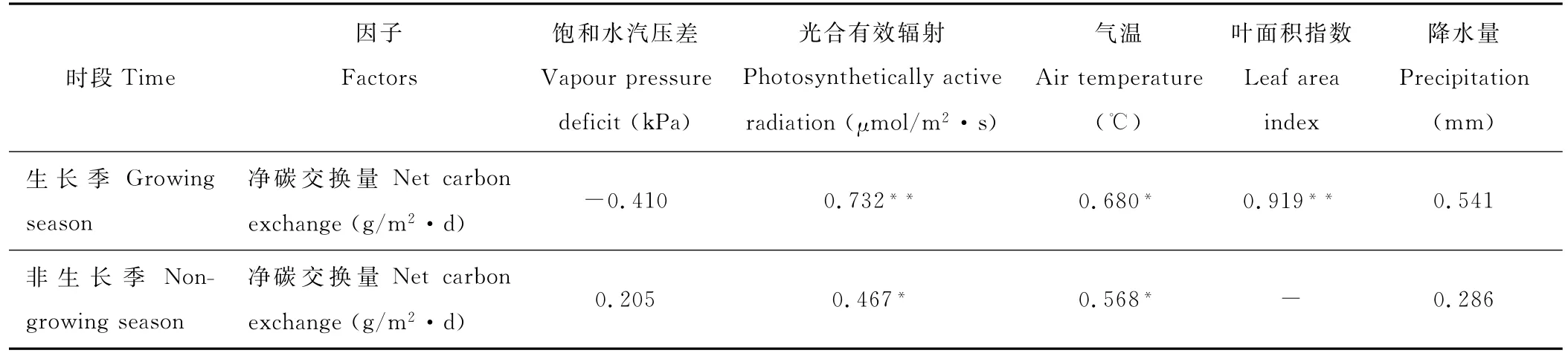
表1 净碳交换NEE与气象因子标准化回归分析Table 1 Normalized regression analysis between net ecosystem CO2exchange(NEE)and meteorology factors
生长季LAI与PAR的偏相关分析(表2)表明,当控制PAR时,LAI与NEE相关性不显著(R2=0.38,P>0.05),而考虑PAR影响时,LAI与NEE相关系数提高了40.6%,二者显著相关(P<0.01),LAI与PAR存在明显的交互作用,共同影响着生态系统净碳交换过程。
2.4 光合有效辐射对NEE的影响
生长季的NEE与PAR变化关系较为分散,通常用一个直角双曲线模型表示(公式3)。其中表观初始光能效率α,利用低光强下NEE-PAR的线性拟合关系公式反推得到[18](NEE=0.0015PAR+0.264,R2=0.924),即α值为0.0015μmol CO2/μmol PAR。由NEE-PAR的拟合曲线(图4a)可以估计光补偿点的范围变化在80~150μmol/(m2·s)之间。

表2 LAI与PAR偏相关分析Table 2 Partial correlation analysis between LAI and PAR
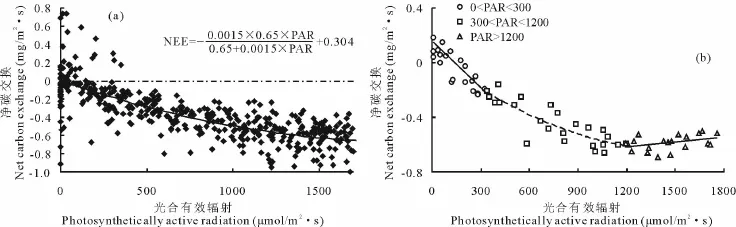
图4 净碳交换NEE与光合有效辐射PAR的关系Fig.4 Relationships between net ecosystem CO2exchange(NEE)and photosynthetically active radiation(PAR)in growing season
草甸生态系统NEE对光强PAR的响应分为3个不同阶段(图4b):PAR<300μmol/(m2·s)时,NEE吸收值随PAR的升高呈线性较快速增加,这时光是唯一的环境限制因素;300<PAR<1200μmol/(m2·s)时,NEE吸收值随PAR的升高呈曲线式较缓慢增长,此时温度、外界CO2浓度等环境因素成为共同的限制因素;PAR>1200μmol/(m2·s)时,NEE吸收量不再随着PAR的升高而增长,即达到光合作用的光饱和阶段。这与许大全等[19]的研究结果相一致。
2.5 水分因素对NEE的影响
水汽压差VPD间接地代表空气的水分状况,有研究指出高光强和高水汽饱和差是植物光合作用降低的两个条件[20]。将生长季VPD观测数据划分为3个区间:低区(0.5~1.0 k Pa)、中区(1.0~1.5 k Pa)、高区(1.5~2.0 kPa),VPD由低区到高区曲线斜率依次增大-减小-增大(图5)。VPD在低区时,随着VPD值的升高,碳吸收速率增加,即更有利于生态系统的碳蓄积;VPD在中区时,NEE吸收速率随VPD的增加继续升高,但增加幅度越来越小,二者变化趋于饱和;VPD处于高区时,NEE吸收速率随VPD增加开始降低,不利于生态系统的碳蓄积,原因可能是高水汽压导致气孔部分关闭,气孔阻力增大,光合作用降低。进而影响草甸生态系统的生产力[21]。
降水是影响生态系统碳交换的重要水分因子,Lawerence等[22]在对加拿大北部温带C3草地生态系统连续观测结果表明,降水充足的年份CO2日吸收峰值约为干旱条件下的3倍。同时降雨格局的差异也会影响生态系统的碳交换能力,以2014年8月16日(短时强降雨)、22日(短时弱降雨)、28日(持续降雨)为例,探讨光照强度、降雨对NEE的共同影响(图6)。
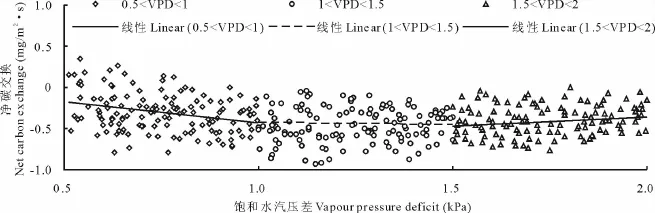
图5 净碳交换NEE与饱和水汽压差VPD的关系Fig.5 Relationships between net ecosystem CO2exchange(NEE)and vapor pressure deficit(VPD)
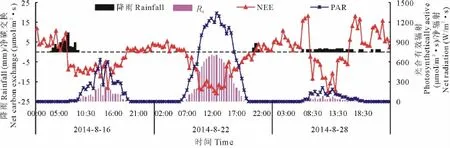
图6 降雨对NEE日变化的影响Fig.6 Diurnal variation of CO2exchange(NEE)under different rainfall conditions
8月16日,降雨开始前3 h内,草甸呼吸速率由0.35 mg/(m2·s)迅速降到0.02 mg/(m2·s),降幅达94%。受凌晨降雨影响,白天PAR仅在0~600μmol/(m2·s)范围波动,而表层土壤含水量出现15.8%的小幅递增,CO2日吸收峰值为0.79 mg/(m2·s),出现在雨后14:00,夜间NEE与雨前碳排放速率大体持平。8月22日,白天NEE与PAR呈明显“单峰”变化,PAR极值达1347μmol/(m2·s),正午12:30左右,到达碳吸收峰值0.93 mg/(m2·s),而夜间21:00-22:00发生了5.8 mm的微量降雨,但未对碳排放造成明显影响。8月28日,降雨历时15.5 h,净辐射(Rn)与PAR均受其影响,数值仅在0~50 W/m2和0~300μmol/(m2·s)范围内波动,表层土壤含水量在降雨开始3 h内缓慢增加,而后稳定。午间13:00左右随雨量增加,生态系统碳汇功能降低(-0.48 mg/m2·s)。夜间受降雨影响,生态系统呼吸速率明显上升。可以看出,降雨削弱了到达地面的辐射强度,却增加了表层土壤水含量,进而影响生态系统碳交换。短暂强降雨过程使白天生态系统碳汇功能降低,而连续低强度降雨虽然使生态系统处于低光强(0<PAR<300μmol/m2·s)的环境,但提高了草甸生态系统夜间的碳交换能力,增加了地温对夜间呼吸的控制[23]。
3 讨论
生长季NEE日变化呈明显“倒钟型”,全天出现一个吸收高峰。日出后的6:00-6:30,草甸植被便开始吸收CO2,8:00-10:30之间达到碳吸收高峰,午间的11:00-13:00由于光饱和作用,碳吸收量减小,出现群落“午休”现象,这与张法伟等[24]研究的青海湖高寒草甸光合午休时间较为一致。
12月至翌年3月草甸植被尚未复苏,4月中下旬植被才开始返青,草甸系统碳吸收微弱,土壤呼吸作用显著。曾有研究指出,非生长季土壤呼吸是生长季生态系统释放CO2的一个重要组成部分,显著影响着系统的碳收支[25],陈骥等[26]在对青海湖北岸高寒草甸碳收支的研究中发现,非生长季土壤呼吸速率是生长季的土壤呼吸速率的0.30~0.33倍。而生长季5-7月生态系统白天的碳吸收强度主要受控于光合有效辐射,二者表现出良好的直角双曲线关系(NEE=+0.304,R2=0.73),而夜间则通过拟合出Vants Hoff方程(公式2)参数,来估算草甸生态系统的呼吸速率,本文利用已获试验数据经拟合确定Q10为2.63,从而建立起碳通量与土壤温度的指数拟合关系(Reco=0.05eIn(2.63)(Ts-10)/10,R2=0.67)。
4 结论
1)科尔沁草甸生态系统碳通量在全年呈现“两峰一谷”的变化规律,草甸生态系统碳吸收总量218.52 g CO2/m2。非生长季主要表现为碳源,净CO2日排放速率均值0.11 mg/(m2·s),生长季主要表现为碳汇,净CO2吸收速率均值-0.17 mg/(m2·s),较北美大草原的-0.88 mg/(m2·s)值偏小[27],这可能与海拔及叶面积指数的差异有关[17]。NEE日变化呈明显的“单峰型”,且日出、日落前后达到碳收支平衡状态。7月12日,10月9日分别出现CO2日吸收(-0.89 mg/m2·s)和日释放(0.56 mg/m2·s)的峰值。
2)叶面积指数对草甸草原生长季NEE的变化影响明显,LAI<0.97时,草甸CO2吸收量随LAI的增加而线性增长,当0.97<LAI<3.08时,CO2吸收量又随LAI的增加而渐进饱和,PAR与LAI的交互作用显著(P<0.01),叶面积指数越大,净碳交换NEE对PAR的响应也越明显。
3)光合有效辐射也是影响NEE的重要环境因子。低光强下,生长季的NEE-PAR呈显著线性相关(R2=0.924),生长季表观量子效率均值为0.0015μmol CO2/μmol PAR,而在PAR>1200μmol/(m2·s)时,NEE对光强的响应表现为渐进饱和型;同时草甸生态系统的碳汇功能受到高VPD值的抑制;短暂强降雨增加了表层土壤水分含量,但削弱了到达地面的辐射强度,降低了昼间土壤呼吸速率,但对夜间NEE无明显影响。而持续降雨使表层土壤含水量维持在较高水平,虽引起昼间碳交换能力的降低,但激发了夜间土壤呼吸速率的增加。
[1] Rank A B,Dugas W A.Carbon dioxide fluxes over a northern,semiarid,mixed grass prairie.Agricultural and Forest Meteorology,2001,108:317-326.
[2] Dugas W A,Heuer M L,Msyeux H S.Carbon dioxide fluxes over Bermudagrass,native prairie,and sorghum.Agricultural and Forest Meteorology,1999,93:121-139.
[3] Shaver G R,Rastetter E B,Salmon V,et al.Pan-Arctic modeling of net ecosystem exchange of CO2.Philosophical Transactions of the Royal Society B-biolonical Sciences,2013,368:1-13.
[4] Fu Y L,Yu G R,Wang Y F,et al.Impact of water stress on photosynthesis and respiration over Leymus chinensis ecosystem in Inner Mongolia.Science in China(Ser.D)Earth Sciences,2006,36(Supp,I):183-193.
[5] Hao Y B,Wang Y F,Sun X M,et al.Seasonal variation in carbon exchange and its ecological analysis over Leymus chinensis steppe in Inner Mongolia.Science in China(Ser.D)Earth Sciences,2006,36:174-182.
[6] Zhao Y P,Zhang X Z,Shi P L,et al.Impact of drought stress on net CO2exchange above an alpine grassland ecosystem in the Gentral Tibetan Plateau.Journal of Resources and Ecology,2013,4(4):327-336.
[7] Li Y Q,Zhao H L,Zhao X Y,et al.Responses of soil respiration to air temperature from June to September in different sanddunes,Horqin sandy land.Journal of Desert Research,2008,28(2):249-254.
[8] Chen N N,Guan D X,Jin C J,et al.Characteristics of soil respiration on Horqin meadow.Chinese Journal of Grassland,2011,33(5):82-87.
[9] LI-COR Biosciences,Eddy Pro 4.0:Help and User’s Guide.Lincoln[EB/OL].[2013-12-02].http://envsupport.licor.com/help/EddyPro4/Default.htm.
[10] Burba G G,McDermitt D K.Addressing the influence of instrument surface heat exchange on the measurements of CO2flux from open-path gas analyzers.Global Change Biology,2008,14:1854-1876.
[11] Li Y J,Xu Z Z,Wang Y L,et al.Latent and sensible heat fluxes and energy balance in a maize agroecosystem.Chinese Journal of Plant Ecology,2007,31(6):1132-1144.
[12] Falge E,Baldocchi D D,Olson R J,et al.Gap filling strategies for defensible annual sums of net ecosystem exchange.Agricultural and Forest Meteorology,2001,107:43-69.
[13] Raich J W,Schlesinger W H.The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate.Tellus,1992,44:81-99.
[14] Xu L J,Tang H J,Yang G X,et al.Variation of net ecosystem carbon flux and its impact factors on Stipa baicalensis steppe in the growing season.Acta Prataculturae Sinica,2011,20(6):287-292.
[15] Shi P L,Sun X M,Xu L L,et al.Net ecosystem CO2exchange and controlling factors in a Steppe-Kobresia meadow on Tibetan Plateau.Science in China(Ser.D),2006,36(Supp,I):194-203.
[16] Suyker A E,Verma S B,Burba G G,et al.Gross primary production and ecosystem respiration of irrigated maize and irrigated soybean during a growing season.Agricultural and Forest Meteorology,2005,131(3/4):180-190.
[17] Chen J L,Wang J,Wang Q,et al.LAI and light transmittance measurement of isolated Vitex shrub using LAI-2200 PCA.Chinese Journal of Agrometeorology,2014,35(4):373-379.
[18] Xu L L,Zhang X Z,Shi P L,et al.Photoionization efficiency in response to temperature under low pressure of alpine meadow on Tibetan Platean.Science in China(Ser.D),2006,36(Supp,II):204-209.
[19] Xu D Q,Li D Y,Shen Y G,et al.On midday depression of photosynthesis of wheat leaf under field conditions.Acta Phytophysiologia Sinica,1984,10:269-276.
[20] Bunce J A.Afternoon inhibition of photosynthesis in maize.Field Crop Research,1990,24:261-271.
[21] Fu Y L,Yu G R,Sun X M,et al.Depression of net ecosystem CO2exchange in semi-arid Leymus chinensis steppe and alpine shrub.Agricultural and Forest Meteorology,2006,137:234-244.
[22] Lawerence B,Flanagan,Linda A,et al.Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland.Global Change Biology,2002,8:599-615.
[23] Zhang F W,Li Y N,Zhao X Q,et al.Effect of one precipitation process on CO2flux and themal transportation in alpine meadow of Qinghai-Tibetan plateau.Chinese Journal of Ecology,2008,27(10):1685-1691.
[24] Zhang F W,Li Y N,Cao G M,et al.CO2fluxes and their driving factors over alpine meadow grassland ecosystems in the northern shore of Qinghai Lake,China.Chinese Journal of Plant Ecology,2012,36(3):187-198.
[25] Wang W,Wang T,Peng S S,et al.Review of winter CO2efflux from soils:a key process of CO2exchange between soil and atmosphere.Chinese Journal of Plant Ecology,2007,31(3):394-402.
[26] Chen J,Cao J J,Wei Y L,et al.Effect of grazing exclusion on soil respiration during the dormant in alpine meadow grassland ecosystems on the northern shore of Qinghai Lake,China.Acta Prataculturae Sinica,2014,23(6):78-86.
[27] Verma S B,Kim J,Clement R J,et al.Water vapor and carbon dioxide exchange at a centrally located prairie site during FIFE.Journal of Geophysics Research,1992,97:18629-18639.
参考文献:
[4] 付玉玲,于贵瑞,王艳芬.水分胁迫对内蒙古羊草草原光合和呼吸作用的影响.中国科学D辑,地球科学,2006,36(增刊I):183-193.
[5] 郝彦宾,王艳芬,孙晓敏,等.内蒙古羊草草原碳交换季节变异及其生态学解析.中国科学,2006,36:174-182.
[7] 李玉强,赵哈林,赵学勇,等.科尔沁沙地夏秋(6-9月)季不同类型沙丘土壤呼吸对气温变化的响应.中国沙漠,2008,28(2):249-254.
[8] 陈妮娜,关德新,金昌杰,等.科尔沁草甸草地土壤呼吸特征.中国草地学报,2011,33(5):82-87.
[11] 李祎君,许振柱,王云龙,等.玉米农田水热通量动态与能量闭合分析.植物生态学报,2007,31(6):1132-1144.
[14] 徐丽君,唐华俊,杨桂霞,等.贝加尔针茅草原生态系统生长季碳通量及其影响因素分析.草业学报,2011,20(6):287-292.
[15] 石培礼,孙晓敏,徐玲玲,等.西藏高原草原化嵩草草甸生态系统CO2净交换及其影响因子.中国科学D辑,地球科学,2006,36(增刊I):194-203.
[17] 陈景玲,王静,王谦,等.植物冠层分析仪测定荆条孤立冠层叶面积指数和透光率的分析.中国农业气象,2014,35(4):373-379.
[18] 徐玲玲,张宪洲,石培礼,等.青藏高原低气压条件下高寒草甸群落光量子效率对温度的响应.中国科学D辑,地球科学,2006,36(增刊I):204-209.
[19] 许大全,李德耀,沈允刚,等.田间小麦叶片光合作用“午睡”现象的研究.植物生理学报,1984,10:269-276.
[23] 张法伟,李英年,赵新全,等.一次降水过程对青藏高原高寒草甸CO2通量和热量输送的影响.生态学杂志,2008,27(10):1685-1691.
[24] 张法伟,李英年,曹广民,等.青海湖北岸高寒草甸草原生态系统CO2通量特征及其驱动因子.植物生态学报,2012,36(3):187-198.
[25] 王娓,汪涛,彭书时,等.冬季土壤呼吸:不可忽视的地气CO2交换过程.植物生态学报,2007,31(3):394-402.
[26] 陈骥,曹军骥,魏永林,等.青海湖北岸高寒草甸草原非生长季土壤呼吸对温度和湿度的响应.草业学报,2014,23(6):78-86.
Net ecosystem carbon exchange and its main driving factors in meadow grassland ecosystems in the Horqin Sand Lands
WANG Jing1,LIU Ting-Xi1*,LEI Hui-Min2*,ZHANG Sheng-Wei1,DUAN Li-Min1
1.Water Resources and Hydropower Survey and Design Institude,Inner Mongolia Agricultural University,Hohhot 010018,China;2.Department of Hydraulic Engineering,Tsinghua University,Beijing 100084,China
The objectives of this study were to investigate the main drivers of ecosystem carbon dynamics in meadow grasslands in the Horqin Sand Lands,and to understand the relationship between net ecosystem CO2exchange(NEE)and key meteorological factors in this region.We continuously measured(NEE)and environmental factors in meadow grasslands in the Horqin Sand Lands from October 2013 to September 2014 using the eddy covariance method.The results showed that the diurnal variation in NEE differed significantly among seasons,especially among vegetation growing seasons.The highest seasonal diurnal absorption rate of CO2was in July,followed by August,September,June,and then May.The NEE was controlled by photosynthetically active radiation(PAR)and the leaf area index(LAI)during vegetation growing seasons.The relationship between NEE and PAR was expressed by a rectangular hyperbolic function.The simulated apparent initial average light-use efficiency was 0.0015μmol CO2/μmol PAR,and the maximum photosynthetic rate(Pmax)was 0.65μmol CO2/(m2·s).The NEE was influenced by LAI,which was expressed by piecewise functions.When the LAI was greater than 3.08,NEE-PAR showed a gradual saturation-type relationship.The higher the LAI,the stronger the response of net carbon exchange to the amount of PAR.A light saturation response occurred when 1.5<VPD<2 k Pa,and NEE was significantly inhibited under high VPD.The day-time net CO2exchange was inhibited by heavy rainfall(cumulative rainfall>40 mm/day),while sustained low-intensity rainfall(rainfall length>15 h)stimulated soil respiration at night.The night-time respiration rate of the ecosystem(Reco)increased exponentially as the soil temperature(Ts)increased,and the temperature-sensitive coefficient Q10was 2.63.
net ecosystem exchange(NEE);meteorological factors;eddy covariance;Horqin sand
10.11686/cyxb2014530 http://cyxb.lzu.edu.cn
王婧,刘廷玺,雷慧闽,张圣微,段利民.科尔沁草甸生态系统净碳交换特征及其驱动因子.草业学报,2015,24(11):10-19.
WANG Jing,LIU Ting-Xi,LEI Hui-Min,ZHANG Sheng-Wei,DUAN Li-Min.Net ecosystem carbon exchange and its main driving factors in meadow grassland ecosystems in the Horqin Sand Lands.Acta Prataculturae Sinica,2015,24(11):10-19.
2014-12-19;改回日期:2015-03-10
国家自然科学基金重点资助项目(51139002),教育部创新团队发展计划(IRT13069)和内蒙古农业大学寒旱区水资源利用创新团队(NDTD2010-6)资助。
王婧(1989-),女,内蒙古阿拉善人,在读硕士。E-mail:lebishu1234@163.com
*通讯作者Corresponding author.E-mail:txliu1966@163.com,leihm@mail.tsinghua.edu.cn
