花魔芋与多叶花魔芋生物学性状比较分析
2015-06-23段龙飞陈国爱郭邦利
段龙飞,陈国爱,郭邦利,刘 婷
(安康市农业科学研究所,陕西安康725021)
花魔芋与多叶花魔芋生物学性状比较分析
段龙飞,陈国爱,郭邦利,刘 婷*
(安康市农业科学研究所,陕西安康725021)
通过对花魔芋及多叶花魔芋生物学性状进行比较分析,发现多叶花魔芋与花魔芋在叶生物量、叶柄颜色、株高、叶柄长度、叶柄直径及小裂叶数上存在显著差异.多叶花魔芋的第二叶光合总色素显著高于花魔芋.多叶花魔芋的单株球茎鲜重比花魔芋提高了17.8%,而球茎葡甘聚糖含量、水分含量无明显差异.该结果可为进一步深入研究多叶花魔芋提供参考.
花魔芋;多叶花魔芋;农艺性状;光合色素;葡甘聚糖
0 前言
魔芋(Amorphophallus konjac)为天南星科魔芋属多年生宿根草本植物,球茎中富含葡甘聚糖,具有降血脂、降血糖、减肥等多种功效,广泛应用于食品、医药、化工及生物等领域[1].随着人们对魔芋特性的不断认识和科学技术的迅猛发展,魔芋新的用途在不断地被开发和利用,国际市场对魔芋需求量快速增大[2].
魔芋属块茎作物,其地上部由球茎顶芽发生一个粗壮叶柄及多次分裂的复叶构成.魔芋为单叶植物,与同科其他芋不同,其叶为一大型复叶.通常情况下,魔芋一个生长周期只出现一片叶,一年中没有新叶的更新替换[3].然而,在花魔芋群体中也发现多叶魔芋,即魔芋在当年主芽抽出第一叶柄后,在主芽基部可再生长第二、第三甚至是多叶.人们对魔芋多叶现象的机理研究表明:魔芋多叶现象是一个很复杂的生理过程,是基因、激素等多层次因素共同作用的结果.已有研究表明,辐照诱变能引起花魔芋的多叶现象.多叶现象的出现是环境主导还是遗传决定未可知[4].由于魔芋的多叶特性可以提高魔芋产量,因此魔芋研究人员采用了多种方法以期获得多叶魔芋.恩施农科院陈永波等[5]利用外源植物激素对魔芋进行诱导发现,多叶的形成主要是由于激素(6-BA)的作用;张盛林等采用辐射诱变技术分别处理了白魔芋和花魔芋,推测魔芋球茎生长过程中的多叶现象与核畸变有关[6-7].但总体来说,对魔芋的多叶研究还处于起步阶段,其研究结果离实际应用差距较大.
目前,对花魔芋和多叶花魔芋形态、生理等的比较分析鲜见报道.基于此,本研究对花魔芋及多叶花魔芋光合色素含量、农艺性状、产量及品质等进行了比较分析,以期为魔芋新品种选育提供参考.
1 材料与方法
1.1 材料
花魔芋根状茎(芋鞭),平均10 g左右.
1.2 方法
试验于2014年4月至2014年10月在陕西省安康市农业科学研究所种芋繁育基地进行.精选优质魔芋根状茎,种植于20*20营养钵中,连栋温室大棚内培养.待魔芋展叶后,分别选择50株魔芋和多叶魔芋用于观察分析.
1.3 测定指标
1.3.1 农艺性状比较分析
待魔芋地上部分定型后,分别选择50株花魔芋和多叶花魔芋进行调查,测定叶生物量、株高、叶柄长度、叶柄直径、小裂叶数.
叶生物量:魔芋地上部分鲜重.叶柄长度:地面到叶柄分叉处的长度.叶柄直径:叶柄基部紧贴地面直径[8].
1.3.2 光合色素含量比较分析
魔芋光合色素含量测定:选择魔芋生长最旺盛季节(球茎膨大期:2014-08-20),采集新鲜叶片,用直径0.9 cm 的打孔器取0.4 g( 避开主叶脉) ,在50 mL容量瓶避光浸提10~12 h,光合色素提取液为丙酮∶乙醇∶蒸馏水= 4∶5 ∶1( 体积比);分别在波长665 mn、649 nm、470 nm和652 nm下测定浸提液的吸光度值,计算叶绿素a、b、类胡萝卜素和总色素的含量.每处理重复5次, 结果取平均值[9].
1.3.3 产量与质量比较分析
收获时各选取花魔芋与多叶花魔芋50株,计算单株球茎鲜重、含水量、硒含量以及葡甘聚糖含量.
魔芋含水量测定:取20株鲜魔芋,分别称重后,在80 ℃恒温下置入烘箱内烘烤,每隔6个小时称重,直至后次称重和前次称重不变为止,记下最后一次重量作为烘干后重量.魔芋含水量计算公式为: 魔芋含水量(%)=(烘前鲜魔芋重-烘后魔芋样重)/烘前鲜魔芋重*100%.
魔芋球茎硒含量测定采用按国家标准GB5009.93-2010, 采用HNO3-HClO4消化-HCl还原-氢化物原子荧光光谱法(Atomic Fluorescence Spectrometry,AFS法), 测定果实硒元素含量[10~11];魔芋葡甘聚糖测定采用3-5二硝基水杨酸法[12].
1.4 数据处理
用DPS(Version 7.05)对试验数据进行统计分析.
2 结果与分析
2.1 花魔芋与多叶花魔芋主要农艺性状比较分析
表1 花魔芋与多叶花魔芋主要农艺性状比较
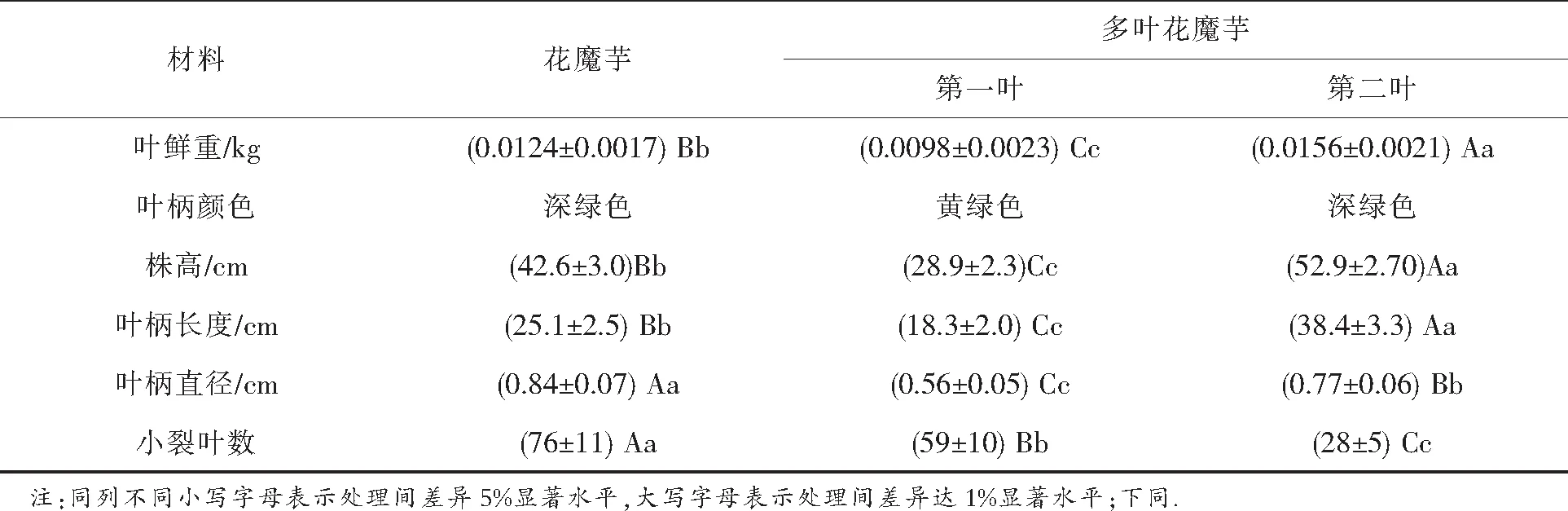

图1 花魔芋与多叶花魔芋
由表1可知,多叶花魔芋叶生物量(地上部分生物量)约为花魔芋的2倍;多叶花魔芋第二叶的株高和叶柄长度明显高于第一叶和花魔芋叶,而多叶花魔芋第一、二叶的叶柄直径、小叶裂片数小于花魔芋.多叶花魔芋第一叶和第二叶的叶柄颜色不同(如图1-D),多叶花魔芋的第二叶和花魔芋的叶柄颜色基本一致,为深绿色,而其第一叶叶柄颜色为黄绿色.
由以上结果表明,花魔芋与多叶花魔芋在叶生物量、叶柄颜色、株高、叶柄长度、叶柄直径以及小裂叶数等农艺性状上存在明显差异.
2.2 花魔芋与多叶花魔芋光合色素含量比较分析
表2 花魔芋与多叶花魔芋光合色素含量比较(mg/g)

由表2可知,花魔芋与多叶魔芋在光合色素含量上也存在显著性差异.具体表现为,多叶花魔芋的第二叶光合总色素、叶绿色a、叶绿色b以及类胡萝卜素高于第一叶.花魔芋叶片叶绿色a和叶绿色b含量均大于多叶花魔芋的第一叶,小于第二叶,类胡萝卜素含量差异不显著.
以上结果表明,多叶花魔芋光和色素合成与积累能力优于花魔芋.
2.3 花魔芋与多叶花魔芋球茎鲜重及品质比较分析
表3 花魔芋与多叶花魔芋球茎鲜重与品质比较

由表3可知,花魔芋与多叶花魔芋的球茎鲜重和膨大倍数存在显著性差异.多叶花魔芋的单株球茎鲜重比花魔芋提高了17.8%,膨大倍数提高了约2.3.多叶花魔芋和花魔芋的水分含量、葡甘聚糖含量差异不显著.多叶花魔芋球茎中硒含量要小于花魔芋,但是两者富硒能力均较低.由此可见,多叶花魔芋叶面积成倍增加,进而有利于叶片同化产物的积累,为球茎的膨大提供了充足的养分,提高了多叶魔芋的鲜重质量,而对魔芋葡甘聚糖含量、水分含量以及硒含量影响甚微.
3 讨论
本研究表明,多叶花魔芋与花魔芋从植株形态上比较发现,多叶花魔芋地上部分生物量明显高于花魔芋,这对于地下球茎干物质的积累有明显的促进作用.同时,多叶花魔芋与花魔芋在株高、叶柄长、叶柄直径、小裂叶数以及叶柄颜色都存在显著性差异.此外,通过多叶多花魔芋与花魔芋光合色素含量分析发现,多叶花魔芋第二叶光合总色素含量显著高于第一叶和花魔芋.因此,笔者认为,由于光合色素含量的差异,从而导致多叶花魔芋第一叶和第二叶叶柄颜色存在差异.多叶花魔芋与花魔芋球茎鲜重和品质的分析结果发现,多叶花魔芋的单株球茎鲜重明显于花魔芋,而葡甘聚糖含量差异不显著.由此说明,多叶花魔芋光合作用合成的同化物质可能没有直接转化为葡甘聚糖.
随着魔芋种植年限的延长和面积的扩大,软腐病病害严重[13],叶片受到创伤感病后会迅速腐烂,导致魔芋失去光合作用的器官,严重影响魔芋光合作用和地下球茎的生长发育.显而易见,魔芋多叶现象不仅成倍增加叶面积,有利于叶片同化产物的积累,为地下球茎的膨大提供了充足的养分[14],而且多叶现象在一定程度上可能会降低病害的发生.因此,花魔芋的多叶现象在生产中应为一个优良的特性.
目前研究表明,花魔芋的多叶现象可能受到基因、激素等多层次因素共同的调节.花魔芋的多叶现象是内在基因变异导致的结果,还是受外界环境的影响还未可知.我们在大田魔芋栽培中发现,未经任何激素、诱变处理的魔芋根状茎大约有1/3出现了双叶现象.笔者认为,由于花魔芋群体种混杂,多年无性繁殖加上自然选择,可能使其性状发生变异,这有可能是群体中产生多叶的一个重要原因.因此,需要魔芋育种工作者进一步从所收集到的当地资源中淘汰单叶类型,保留多叶型材料,并纯化种源,从而选育出多叶花魔芋品种,这将是培育多叶的魔芋品种最直接、最有效的办法[15].
[1] 刘佩英. 魔芋学[M]. 北京:中国农业出版社, 2004.
[2] 牛义, 张盛林,王志敏等. 中国魔芋资源的研究与利用[J]. 西南农业大学学报(自然科学版),2005,27(5):635-637.
[3] 李川. 魔芋“多叶”现象的研究进展[J]. 长江蔬菜(学术版),2008,11b:4-6.
[4] 石新龙.人工调控对魔芋产生多叶影响的研究[D].西南农业大学, 2011.
[5] 陈永波,钟刚琼,赵清华等.外源激素对魔芋多叶、多球茎形成的影响[J].氨基酸和生物资源,2005,27(1):35-36.
[6] 张盛林,李川,刘佩瑛等.60Co-γ 射线辐射对花魔芋性状影响初探[J].中国农学通报,2004,20(5):183-184.
[7] 黄训端,周立人,何家庆等.60Co-γ 射线辐照花魔芋球茎的早期诱导效应研究[J].激光生物学报,2004,13(4):306-313.
[8] 许宁国,杨朝柱,刘二喜等. 不同育龄花魔芋主要农艺形状比较研究及其在栽培学中的应用[J]. 湖北民族学院学报( 自然科学版),2013,31(3):306-308.
[9] 萧浪涛,王三根. 植物生理学实验技术[M]. 北京:中国农业出版社,2005.
[10] NixonD.E.,NeubaueK.R.,EekdahlS.J.etal. Evaluation of a tunable bandpass reaction cell inductively coupled plasma mass spectrometer for the determination of selenium in serum and urine[J]. Spect.Acta.A.Mo.I.Biomol.Speet.,2003,58:97-110.
[11] Worbel k., CuarsoJ.A.,etal. Perteratment porcedures for characterization of arsenie and Selenium Species in complex samples utilizing coupled teehniques with mass spectrometric detection[J]. Analy.Bioanaly.Chem.,2005,381(2):317-331.
[12] 欧阳华学,韩梅,刘佩英等. 分光光度法测定魔芋葡甘露聚糖含量[J].西南农业学报,2002, 15(1):109-111.
[13] 丁自立,万中义,矫振彪等.魔芋软腐病研究进展和对策[J].中国农学通报,2014,30(4):238-241.
[14] 刘海利, 牛义, 石新龙等. 植物生长物质处理对花魔芋产生多叶的影响[J]. 西南农业大学学报(自然科学版),2012,34(12):40-42.
[15] 周富忠,吴卓耕,田祚旭等. 一种新的多叶花魔芋资源[J]. 长江蔬菜(学术版),2010,20:16-17.
Comparative Study on Biological Characters of Konjac and Multi-leaf Konjac
DUAN Long-fei, CHEN Guo-ai, GUO Bang-li, LIU Ting
(Agricultural Science Research Institute of Ankang of Shannxi Province, Ankang 725021, China)
The comparative study on the biological characters of konjac and multi-leaf konjac was conducted in the study. The results show that there were significant differences in leaf biomass, petiole color, height, petiole length, petiole diameter and number of leaves between multi-leaf konjac and konjac. The second of multi-leaf konjac photosynthetic pigments were significantly higher than konjac; The Corm fresh weight of Multi-leaf konjac increased by 17.8%, and the glucomannan, moisture content had no significant difference between them. This results can provide reference for further study of multi-leaf konjac.
Konjac;Multi-leaf Konjac; Agronomic traits; Photosynthetic pigment; KGM
2015-05-20
陕西省科学技术研究发展计划项目(2013K01-24).
*
段龙飞(1987—),男,陕西西安人,硕士,助理农艺师,主要从事魔芋栽培技术及育种研究.
S632.9
A
1009-2102(2015)02-0021-04
