棉属三元杂种的合成及细胞遗传学鉴定
2015-06-18荣二花李灵娇杨娜陈弟吴玉香
荣二花,李灵娇,杨娜,陈弟,吴玉香*
(1.山西农业大学 实验教学中心,山西 太谷 030801;2.中国农业科学院 棉花研究所,河南 安阳 455000;3.山西农业大学 农学院,山西 太谷 030801)
棉属三元杂种的合成及细胞遗传学鉴定
荣二花1,李灵娇3,杨娜3,陈弟2,吴玉香3*
(1.山西农业大学 实验教学中心,山西 太谷 030801;2.中国农业科学院 棉花研究所,河南 安阳 455000;3.山西农业大学 农学院,山西 太谷 030801)
棉属有丰富的种质资源,野生棉具有栽培棉种所缺乏的许多优良性状,为了将野生澳洲棉和达尔文棉的优良性状导入陆地棉,以期改良栽培种陆地棉。本研究通过陆地棉和C组野生澳洲棉远缘杂交获得杂种F1,并对其进行染色体加倍,将加倍成的异源六倍体与野生四倍体达尔文棉杂交,获得陆地棉、澳洲棉、达尔文棉的三元杂种,并对该杂种进行形态学和细胞遗传学鉴定。形态学观察结果表明:三元杂种有三个亲本的遗传特性。花粉母细胞减数分裂结果表明后期存在异常行为,主要表现在65%的花粉母细胞减数分裂后期染色体不能均等分离,使得末期形成大量的多分体,这些多分体会进一步发育为败育花粉粒,统计结果表明正常花粉粒只占18.3%,导致该杂种高度不育。本研究不仅合成了综合三个亲本特性的三元杂种,也为该杂种不育提供了细胞遗传学证据,并为棉花新种质的创制提供了理论依据和中间材料。
陆地棉;澳洲棉;达尔文棉;三元杂种;形态学;减数分裂
棉花属双子叶植物(Dicotyledons)、锦葵科(Malvaceae)、棉属(Gossypium)。棉属有4个亚属,8个组,9个亚组,51个种,其中46个为二倍体棉种,5个为四倍体棉种[1]。棉属有四个栽培品种即亚洲棉、草棉、陆地棉、海岛棉,其余47个均为野生棉种[2]。由于栽培棉种对病虫害及非生物胁迫较为敏感[3],而野生棉种有多种多样优异的农艺性状,如抗干旱、盐碱,抗病虫,抗寒,种子无棉酚而植株具棉酚,以及纤维品质优异等。通过远缘杂交,期望将不同棉种所特有的有益性状进行遗传重组,创造出品种间杂交无法得到的新种质,是棉花遗传育种的重要途径[4]。
陆地棉为四个栽培棉种之一,是世界90%以上的栽培棉种,也是最重要的纤维作物。澳洲棉和达尔文棉均为野生棉种,澳洲棉具有抗病虫及抗干旱、抗短期低温等优良特性,并能提高衣分和强力[5]。利用野生棉改良陆地棉国内外都有很多成功的报道:如早在60年代 Muramoto[6]将澳洲棉和陆地棉杂交,F1加倍,再对六倍体进行选择,第四代具备比陆地棉和海岛棉更大的耐寒性,抗根腐病菌,且改进了纤维品质。梁理民等人[7]通过陆地棉和斯特提棉种间杂交育成新品种秦远号,同时选育出抗病优质、抗病高强、抗病大铃、抗病高衣分,抗枯黄萎病的棉花新种质。迄今为止,我国已获得陆地棉与19个野生棉种及其他栽培种的高代种间杂种(4代以上)[8]。
本研究通过远缘杂交试图将野生澳洲棉和达尔文棉的优良性状导入陆地棉,合成综合野生棉和陆地棉优良性状的新种质,并对杂种进行形态学鉴定和减数分裂行为探讨,以期寻求杂种不育的直接原因,并进一步探讨育性恢复的有效方法,为棉花遗传改良及新种质创制提供理论依据和有价值材料。
1 材料和方法
1.1 实验材料
本实验所用材料有:陆地棉(G.hirsutum)(2n=4x=52)、C组野生澳洲棉(G.australe)(2n=2x=26)、四倍体野生达尔文棉(G.darwinii)(2n=4x=52)。野生棉材料来源于国家种质三亚野生棉圃,其染色体组及地理分布见表1。

表1 供试材料及其地理分布Table 1 The materials and geographic distribution
三元杂种合成过程如下:
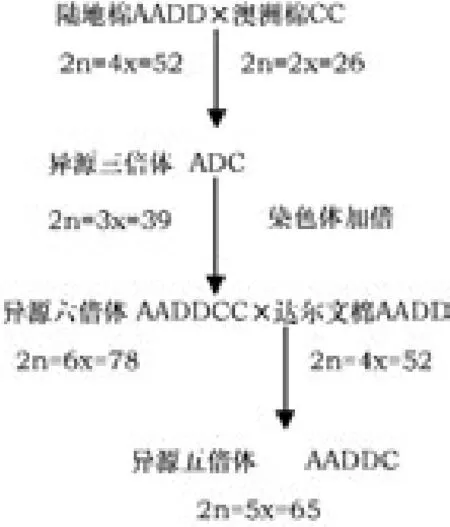
1.2 三元杂种的形态学观察和鉴定
对三元杂种植株和其三个亲本进行形态学观察鉴定并比较,分别选取同一节位(第五节)的三个亲本叶片和杂种叶片进行比较,并选取同一节位(第五节)的三个亲本花朵和杂种花朵进行比较,包括花瓣、雌雄蕊等性状差异较大的部位分别进行比较,判断亲本的遗传物质及形态性状在杂种中的表现。
1.3 三元杂种的花粉母细胞减数分裂制片及观察
花粉母细胞减数分裂观察制片和照相采用李懋学等[9]的方法。挑选各个时期分散好的片子用OLYMPUS DP71显微镜观察并拍照,分别选取60个视野,观察减数分裂后期染色体分离状况,记录多分体出现的种类及数目,并计算其各自所占比例。
1.4 三元杂种的花粉粒制片及观察
花粉粒观察制片和照相也采用李懋学等[9]的方法。挑选花粉粒分散好的片子用OLYMPUS DP71显微镜观察并拍照,选取40个视野观察统计花粉粒类型和出现的频率,并计算其各自所占比例。
2 结果与分析
2.1 三元杂种的形态学观察和鉴定
三元杂种植株长势良好,表现出了强的杂种优势。选取同一节位叶片及花朵进行比较(图1)。从图1中可以看出:杂种叶片为阔心型,有异源六倍体中间亲本的特性,综合亲本陆地棉叶片阔和澳洲棉椭圆形状的特征。花朵淡粉色,比异源六倍体中间亲本淡,但比亲本澳洲棉深,亲本澳洲棉紫色,杂种没有明显花斑,不像亲本澳洲棉,雌蕊柱头像母本陆地棉,雄蕊遗传了陆地棉黄色花药和野生达尔文棉亲本花丝短的特性,但更像中间亲本异源六倍体。

图1 杂种叶片及花朵和亲本比较Fig.1 Leaf and flower comparison with its parents
2.2 花粉母细胞减数分裂的观察
正常花粉母细胞减数分裂在末期Ⅱ会形成由4个细胞核组成的四分体,最终产生4个子细胞。而微核是由染色体后期Ⅱ向多极不均等分离形成的,一个花粉母细胞内可能形成了5个或5个以上大小不等的核,三元杂种的花粉母细胞,在减数第二次分裂后期,染色体向多极不均等分离,观察到不同数目的微核;即多分体,本研究选取60个视野中200个多分体进行统计分析,发现四分体70个,比例为35%,多分体及有微核出现的多达130,频率为65%。不同多分体出现的个数和比例的统计如表2。

表2 杂种花粉母细胞减数分裂后期多分体出现情况Table 2 Number of multispores in telophase II of hybrid in pollen mother cells

图2 三元杂种减数分裂后期出现的各种多分体Fig.2 Various multispores in meiosis of telophase II in hybrid
2.3 三元杂种花粉粒的观察
三元杂种植株在自然授粉情况下不结铃,大量杂交、自交不结种子,说明三元杂种不育,导致这种不育的原因之一是性细胞减数分裂异常。探究其花粉粒形状和饱满度可以为其育性恢复提供理论依据。
三元杂种花粉粒发育不整齐,大小不一,形状各异,既存在发育饱满的大花粉粒,也存在极小的败育花粉粒,正常花粉粒大小均匀,形状规则均为饱满的球形,如图3所示左边为正常花粉粒,即完整的饱满球形花粉粒,右边为无刺突花粉粒以及不完整的破裂花粉粒或干瘪的椭圆状花粉粒。对40个视野中的230个花粉粒的统计结果如表3,表明三元杂种正常的花粉粒只占18.3%,大部分为异常花粉粒。这是三元杂种不育的直接原因。
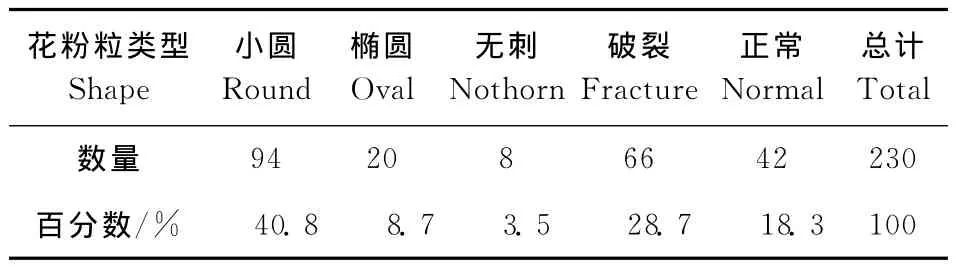
表3 三元杂种花粉粒类型及出现频率Table 3 The number and frequency of different pollen grains in trispecific hybrid
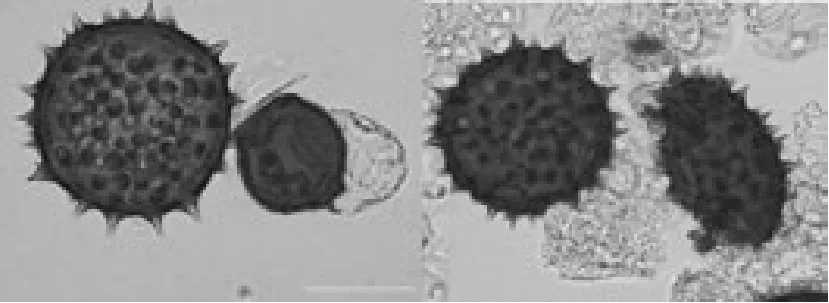
图3 三元杂种花粉粒类型Fig.3 Pollen grain type in hybrid.
3 讨论
3.1 三元杂种F1不育原因及育性恢复方法探讨
杂种不育在种间杂种中普遍存在,部分或完全阻断了不同种间遗传信息的交换。原因在于异种间遗传性的不协调,在棉属中常见的有细胞核不育和细胞质不育。前者指的是双亲染色体结构上的差异,染色体组型之间的不相适应,即不具有或缺少同源性,减数分裂时不能正常配对或配对后不能正常分裂,产生畸形花粉粒和畸形雌雄配子,导致杂种不育。细胞质不育性则是由于异种间的细胞质和细胞核不协调而产生[10]。本研究中三元杂种高度不育的直接原因在于减数分裂紊乱,染色体分离不均等,向多极移动产生多分体,最终形成的配子得不到全套染色体,花粉粒无活力。三元杂种F1不育性主要体现在花粉没有生活力。有望通过以下途径加以克服:强制自交和大量回交;控制营养和环境条件;染色体加倍等。Benbouza等[11]报道通过远缘杂交获得雷蒙地棉、斯特提棉、陆地棉三元杂种,通过人工授粉使三元杂种与亲本陆地棉大量回交以及自交,成功获得了可育的稳定遗传后代。采用三元杂种与栽培棉亲本回交也可获得很好的效果[12]。吴玉香等[`13]研究证明延长种间杂种植株的生长时间有利于协调不同来源的染色体组之间的关系,使染色体组趋于平衡,育性得到大幅度恢复。本研究中的三元杂种有望通过以上方法恢复其育性,选育出可育的杂种遗传后代,为棉花遗传育种及新种质创新提供理论依据和有价值材料。
3.2 克服远缘杂交不亲和的方法
杂交不亲和是远缘杂交育种的首要难题。主要分为受精前障碍和受精后障碍两类[14]。克服受精前障碍的方法有:(1)改良授粉方式,如混合授粉、授 蒙 导 花 粉、蕾 期 授 粉 和 重 复 授 粉[15,16],Sundaram等[17]探究了三叶棉、戴维逊氏棉、瑟伯氏棉、辣根棉四种野生棉在海岛棉雌蕊上的花粉管行为,认为蕾期授粉、试管内授粉可有效促进授粉成功。(2)应用激素类物质如赤霉素、萘乙酸或秋水仙素等涂抹或喷洒柱头[15],通过吸引花柱中促进花粉管生长的物质促进花粉管生长,抑制花的脱落,提高受精率[18]。山西省农科院[19]在培育特早熟抗蚜棉花新品种晋棉51号过程中,对获得的陆地棉×异常棉的远缘杂种后代采用杂交铃喷(滴)GA3和NAA、离体培养杂交当代幼胚技术,成功克服了杂交当代不亲和性。(3)体细胞杂交技术使得不兼容物种间的基因融合以及细胞核或细胞质性状的转移成为可能[16],孙玉强等[20]通过体细胞杂交技术成功地将野生三裂棉和陆地棉原生质体融合,将三裂棉优良性状转入陆地棉,杂种植株具有比双亲更强的光合能力及营养器官。(4)在授粉前对花粉、花柱进行一定的物理处理,如适当加热、电磁辐射、紫外线和γ射线照射等,可以提高受精率,对克服受精前障碍有积极帮助[21]。受精后障碍的克服常采用幼胚拯救技术,包括子房培养、胚珠培养和幼胚培养,这在菊科杂交种获得成功[22,23]。此外,染色体数目不同的双亲杂交,染色体数目多的一方做母本成功率较大;多倍体起到基因转移的载体作用和基因渐渗的媒介作用,可以把野生种中简单遗传的抗病基因转移到栽培种中[24],Robinson等[25]通过六倍体做桥梁获得陆地棉、长萼棉和辣根棉三元杂种,该杂种能抗线虫,大大解弱了美国棉花受线虫的危害。本研究以染色体多的陆地棉做母本,和二倍体澳洲棉杂交制成杂种三倍体,再对其进行染色体加倍,获得了可育的异源六倍体杂种,但该六倍体杂种与达尔文棉杂交后获得不育三元杂种,有待进一步寻找恢复育性的有效方法。
[1]Fryxell P A.A revised taxonomic interpretation of Gossypium L.(Malvaceae)[J].Pheedea,1992,2(2):108-165.
[2]王坤波,杜雄明,宋国立.棉花种质创新的现状与未来[J].植物遗传资源学报,2004,5(增刊):23-28.
[3]Yuqiang Sun,Shoumei Liu,Yu Wang,et al.An interspeci?c somatic hybrid between upland(G.hirsutumL.cv.ZDM-3)and wild diploid cotton(G.klotzschianumA.)[J].Plant Cell and Organ Culture,2011,106(2):425-433.
[4]梁正兰.棉花远缘杂交的遗传与育种[M].北京:科学出版社,1999:1-2.
[5]梁正兰,孙传渭.棉花远缘杂交[M].北京:科学出版社,1982:32.
[6]Muramoto H.Hexaploid cotton:some plant and fiber properties[J].Crop Sci,1969,9(1):27
[7]梁理民,刘有良,王增信,等.陆地棉×斯特提棉种间杂交创造抗枯黄萎病新种质[J].西北农业学报,2002,11(4):16-18.
[8]庞朝友,杜雄明,马峙英.具有野生棉外源基因的陆地棉特异种质创造与利用进展[J].棉花学报,2005,17(3):171-177.
[9]李懋学,张赞平.作物染色体及其研究技术[M].北京:中国农业出版社,1996:63-66.
[10]中国农业科学院棉花研究所.中国棉花遗传育种学[M].济南:山东科学技术出版社,2003:270
[11]H Benbouza,J M Lacape,J M Jacquemin,et al.Introgression of thelow-gossypolseed&high-gossypolplanttrait in upland cotton:Analysis of[(Gossypiumhirsutum×G.raimondii)2×G.sturtianum]trispecific hybrid and selected derivatives using mapped SSRs[J].Molecular Breeding,2010,25(2):273-286.
[12]Ganesh S Newaskar,Vivek P Chimote,Subhash S Mehetre,et al.Interspecific hybridization inGossypiumL.:characterization of progenies with different ploidy-confirmed multigenomic backgrounds[J].Plant Breeding,2013,132(2):211-216.
[13]吴玉香,陈崇乾,高燕会,等.活体保存对栽培棉种间四元杂种育性和染色体构型的影响[J].棉花学报,2007,19(6):461-466.
[14]邓衍明,叶晓青,佘建明,等.植物远缘杂交育种研究进展[J].华北农学报,2011,26(增刊):52-55.
[15]汤访评.菊属与四个近缘属植物远缘杂交研究[D].南京:南京农业大学,2009.
[16]李玉晖,陈学森,郑洲.果树远缘杂交育种研究进展[J].山东农业大学学报,2003,34(1):139-143.
[17]Sundaram Ganesh Ram V,Thiruvengadam S,Hari Ramakrishnan,et al.Investigation on pre-zygotic barriers in the interspecific crosses involvingGossypiumbarbadenseand four diploid wild species[J].Euphytica,2007,159:241-248.
[18]孟金陵.植物生殖遗传学[M].北京:科学出版社,1997.
[19]郭宝德,黄穗兰,冀丽霞.抗蚜棉花新品种晋棉51号的选育及高产栽培技术[J].山西农业科学,2011,39(9):933-935,971.
[20]X S Yu,B J Chu,R E Liu,et al.Characteristics of fertile somatic hybrids ofG.hirsutumL.andG.Trilobumgenerated via protoplast fusion[J].Theoretical and Applied Genetics,2012,125(7):1503-1516.
[21]Atanassov II,Atanassova SA,Dragoeva AI,et al.A new CMS source in Nicotiana developed via somatic cybridization betweenN.tabacumandN.alata[J].Theor Appl Genet,1998,97:982-985.
[22]Henn HJ,Wingender R,Schnabl H.Regeneration of fertile interspecific hybrids from cell fusion betweenHelianthusannuusL.and wildHelianthusspecies[J].Plant Cell Rep 1998,18:220-224.
[23]Cheng X,Chen S M,Chen F D,et al.Interspecific hybrids betweenDendranthemamorifolium(Ramat.)Kitamura andD.nankingense(Nakai)Tzvel.Achieved using ovary rescue and their cold tolerance characteristics[J].Euphytica,2010,172:101-108.
[24]孙敏红,张蜀宁.多倍体育种在园艺作物中的应用[J].江苏农业科学,2004(1):68-72.
[25]Robinson A F.Reniform in U.S.cotton:when,where,why,and some remedies[J].Ann Rev Phytopathol,2007,45:111-125.
Synthesis and Cytogenetic Identification of Trispecific Hybrid in Cotton
Rong Erhua1,Li Lingjiao3,Yang Na3,Chen Di2,Wu Yuxiang3*
(1.Experimentalandteachingcenter,ShanxiAgriculturalUniversity,TaiguShanxi030801,China;2.CottonResearchInstituteofChineseAcademyofAgriculturalScience,AnyangHenan455000,China;3.CollegeofAgriculture,ShanxiAgriculturalUniversity,TaiguShanxi030801,China)
There are rich germplasm resources inGossypiumand wild cotton species possess many valuable agronomic traits which cultivated cotton always lacks.In order to introgress the useful traits from wild speciesG.australeandG.darwiniiinto cultivated species,tetraploidG.hirsutumwas crossed directly with C-genome speciesG.australeas male parent,creating a triploid hybrid.Chromosome doubling of this triploid hybrid leads to a allohexaploid which was then crossed with wild tetraploid speciesG.darwinii,resulting in trispecific hybrid(G.hirsutum,G.australe,G.darwinii).The morphology characteristic and cytogenetic identification of trispecific hybrid was further investigated in this study.The hybrid morphology performed combining characteristics from three parents.Results indicated that the meiosis of trispecific hybrid was abnormal,and 65%abnormal multispores appeared in the telophase II,which finally developed into abortive pollen grains.The result also showed that there were only 18.3%normal pollen grains in this hybrid by statistic analysis.It leads to the sterility of trispecific hybrid F1.This study not only creats new cotton germplasms fromG.hirsutum,G.australe,G.darwiniiwith three parents characteristic.It also provides the main reason of sterility for this hybrid.
G.hirsutum;G.australe;G.darwiniiWatt;Trispecific Hybrid;Morphology;Meiosis
S334.3
A
1671-8151(2015)05-0449-05
10.13842/j.cnki.issn1671-8151.2015.05.001
2015-06-03
2015-07-09
荣二花(1974-),女(汉),山西太谷人,博士,研究方向:作物信息技术。
*通讯作者:吴玉香,教授,博士。Tel:15034664402,E-mail:yuxiangwu2009@hotmail.com
国家自然科学基金(No.31171599,31301364);山西农业大学科技创新基金(2014YZ04);博士基金(2013YJ45);博士后基金(125375)
(编辑:邢国芳)
