基因型×环境互作下小麦氮代谢相关性状的遗传和相关性分析
2015-06-15王琳琳王振林孙爱清王春微尹燕枰
王琳琳, 王 平, 王振林, 孙爱清, 杨 敏, 王春微, 尹燕枰*
(1山东农业大学农学院, 作物生物学国家重点实验室, 山东省作物生物学重点实验室,山东泰安 271018;2泰安市农业科学研究院,山东泰安 271000)
基因型×环境互作下小麦氮代谢相关性状的遗传和相关性分析
王琳琳1, 王 平2, 王振林1, 孙爱清1, 杨 敏1, 王春微1, 尹燕枰1*
(1山东农业大学农学院, 作物生物学国家重点实验室, 山东省作物生物学重点实验室,山东泰安 271018;2泰安市农业科学研究院,山东泰安 271000)
【目的】选育氮高效的小麦品种,可有效提高氮素利用效率和生产效率,对环境安全至关重要。本文分析了小麦氮代谢相关性状的遗传效应,为小麦氮高效品种选育提供理论依据。【方法】选用7个小麦品种及其组配的12个杂交组合,进行田间盆栽试验。设置3个氮水平,利用基因型与环境互作的加性-显性遗传模型,对氮代谢相关的10个性状进行遗传与相关性分析。【结果】株高、开花期和成熟期单茎干物重、开花期氮素积累量、籽粒氮素积累量和氮素吸收总量主要受加性效应控制,花后氮素同化量受显性×环境互作效应影响较大,氮素利用效率、氮素生理效率以加性×环境互作效应为主。10个性状狭义遗传力总体不高(平均值为0.56),广义遗传力总体较高(平均值为0.881)。互作广义遗传力均达到1%显著水平,表明不同的氮水平对遗传表达有较大影响。氮素利用效率、氮素生理效率和开花期氮素积累量的互作狭义遗传力较大,表明不同氮水平对这些性状的选择效果不同。通过加性效应预测值得出,亲本DK138和JN10的氮素利用效率和氮素生理效率的加性效应为显著正效应。大多数组合的显性主效应与不同氮水平下的显性×环境互作效应在方向上不尽一致,表明小麦氮高效杂交后代的选择宜考虑特定的氮水平条件。显性效应预测值表明,组合JN10×W9903的氮素生理效率显性效应值最大且达到显著水平,是氮素生理效率较高的组合。相关分析表明,两两性状间以加性遗传相关为主。氮素生理效率与株高呈加性正相关关系,达到10%显著水平。除株高和谷氨酰胺合成酶活性外,氮素利用效率与其他性状间以显性环境互作相关为主。氮素利用效率与氮素生理效率之间的显性×环境互作相关系数达到10%显著水平。氮素利用率与氮素生理效率的表现型和基因型相关系数为正值且达1%显著水平。【结论】通过性状分析表明,株高在一定程度上可以作为氮素生理效率的间接选择性状,氮素利用效率与氮素生理效率这两个性状进行协同改良。品种DK138和JN10可作为亲本以提高后代的氮素利用效率和氮素生理效率。杂交组合LM14×W9903表现出良好的后代选育利用潜力。
小麦; 氮代谢; 遗传; 加性-显性模型; 基因型×环境互作
氮是小麦生长发育所必需的大量营养元素之一。为了提高小麦产量,氮肥的实际施用量已远远超过最佳经济施用量,不仅造成经济浪费,还造成了环境污染。因此,在小麦生产中,提高氮素利用效率,选育氮高效的小麦品种至关重要。对于氮素利用效率的定义各有所解。对以收获籽粒的禾谷类作物而言,可采用经济利用效率,即植株中积累的单位氮所生产得到的籽粒干重[1]。刘强等[2]认为,在统一供氮条件下,可以作物籽粒产量与地上部氮素积累量之比来表征氮素利用效率。研究发现,玉米[3]、水稻[4]、小麦[5]等的不同基因型对氮素利用存在显著差异,且是由基因型与环境因素共同作用所致。因此,在基因型×环境互作下对氮代谢相关性状的研究显得十分必要。
Sinebo等[6]通过田间试验,研究了低氮条件下不同基因型大麦成熟期氮素利用相关的性状及产量的遗传变异。Barraclough等[7]对小麦氮效率基因型与环境变异的改良研究中,发现高秆型与矮秆型品种间籽粒氮素吸收总量和籽粒氮素利用效率差异显著,矮秆型品种间差异也显著;同时得出,籽粒氮含量与籽粒氮素利用效率间呈显著的负相关关系,籽粒氮含量高时,籽粒氮素利用效率低。王庆仁等[8]研究表明,在低氮水平时吸收效率和利用效率共同左右产量,但是利用效率的影响较大;而高氮水平时主要是吸收效率起作用,利用效率的影响较小。谷氨酰胺合成酶处于氮代谢中心且参与氮代谢的调节,其活性的提高可使氮代谢运转增强,促进氨基酸合成和转化[9-10]。因此,谷氨酰胺合成酶也在氮代谢中发挥着重要作用。周桃旺[11]在对玉米杂种一代遗传特性的研究中发现,组配耐低氮胁迫的玉米杂交种时,应选用耐低氮能力强的种质材料作为母本,为选育优良后代组合奠定基础。朴钟泽等[12]在两个氮水平下对水稻氮效率相关性状的配合力进行了分析,表明在两个氮素水平下,最高分蘖期和孕穗期的干物重与氮素利用效率的亲本加性效应均对F1起主导作用,收获期氮素利用效率在施氮条件下遗传变异主要来自加性效应,而在未施氮条件下主要来自非加性效应。
我国对小麦氮高效品种的筛选和对农艺性状的遗传效应研究相对较多[13-15],对不同氮水平下小麦氮代谢相关性状遗传效应的研究尚少。对小麦氮代谢的研究,如郝西[16]在两种环境下,于成熟期测定小麦籽粒、叶片和茎秆的全氮、全磷含量,利用Griffing[17]的双列杂交分析方法估算配合力效应值及配合力方差,未就更多的代谢相关性状做进一步分析。本研究在三种氮水平下,采用基因型与环境互作的加性-显性遗传模型,分析了小麦氮代谢相关性状的遗传效应与遗传相关性,以期减少亲本组配的盲目性,提高氮高效育种的遗传改良效率,对氮高效遗传育种的亲本选配和后代选择具有重要意义。
1 材料与方法
1.1 供试材料
选用豫麦35(YM35)、济南10号(JN10)、鲁麦14(LM14)做母本;大科138(DK138)、聊麦18(LM18)、临汾6010(LF6010)、潍9903(W9903)做父本,采用不完全双列杂交设计,组配成12个杂交组合。亲本选择时综合考虑了氮效率类型等因素。如亲本YM35为低氮高效品种,亲本JN10为氮高效品种,亲本LM14为双高效品种,亲本DK138为高氮高效品种,亲本W9903为氮低效品种。
1.2 试验方法
试验在山东农业大学教学试验基地进行。亲本于2011年10月种植,常规管理。2012年5月做杂交并于6月收获得到F1代杂交种子。2012年10月将亲本及其杂交F1代盆栽种植,随机区组设计,3次重复,每盆定植12株。设3个N水平: 0.1(N1)、1.5(N2)和2.5 g/pot(N3),相当于大田施N量16、240和400 kg/hm2。P2O5用量为0.7 g/pot,相当于大田施用P2O5115 kg/hm2;K2O用量0.4 g/pot,相当于大田施用K2O 65 kg/hm2。磷钾肥全部底施,氮肥50%底施,50%拔节期追施。试验用盆规格为直径30 cm,深度26 cm。试验盆埋入土壤,距盆口5 cm处与地面平齐,盆内土壤为过筛(0.85 mm)土。盆内起始土壤全氮含量为1.18 g/kg。
1.3 取样方法与测定
开花期取样一次,每处理组合取3盆,每盆选3株,每株取1个有代表性的带穗单茎,于105℃杀青30 min后80℃烘至恒重,用于单茎干物重、全氮含量测定。另取9个单茎的倒二叶用液氮速冻后放入-40℃冰箱中保存,用于谷氨酰胺合成酶活性测定。
成熟期田间测量株高,取样方法同开花期。取9个带穗单茎于80℃烘至恒重,用于单茎干物重、单穗粒重,单茎氮含量、籽粒氮含量测定。谷氨酰胺合成酶活性参照王月福等[9]的方法测定,植株氮含量采用浓H2SO4-H2O2消化,半微量凯氏定氮法[18]测定。
1.4 计算方法
氮素积累量=含氮量×干重
开花期氮积累量为开花期单茎、叶、鞘、穗氮积累量的总量;氮素吸收总量为成熟期单茎、叶、鞘、穗、籽粒氮积累总量。
花后氮素同化量(mg)=氮素吸收总量-开花期氮积累量
氮素利用效率(g/g)=单茎穗粒重(单茎籽粒产量)/氮素吸收总量[2]
氮素生理效率(g/g)=成熟期总干物重(单茎生物量)/氮素吸收总量[19]
1.5 统计方法
利用Microsoft Excel整理原始数据,并计算各性状的平均值和变异系数。数据分析参照加性-显性与环境互作的遗传模型[20-22]进行。采用QGA Station2.0分析软件估算各项方差分量、各性状间的遗传相关系数。以调整的无偏预测法(AUP法)[22]对各遗传效应值进行预测。
表型方差分量VP=VA+VD+VAE+VDE+Ve
2 结果与分析
2.1 亲本和F1代氮代谢相关性状特征值
由表1可见,就最小值、最大值和平均值而言,F1代株高与开花期单茎干物重在3种氮水平下均高于亲本;开花期氮素积累量、籽粒氮素积累量和氮素吸收总量在N1和N2水平下F1代高于亲本,在N3水平下小于亲本;谷氨酰胺合成酶活性和花后氮素同化量在N1水平下F1代高于亲本,在N2和N3水平下小于亲本;F1代成熟期单茎干物重和氮素利用效率在3种氮水平下与亲本相等或接近;F1代氮素生理效率小于亲本。从变异系数来看,亲本各性状在3个氮水平下的变异系数高于F1代,N3水平下的谷氨酰胺合成酶活性和N2水平下的开花期单茎干物重、开花期氮素积累量和氮素生理效率除外。除氮素生理效率外,其他性状的均值在低氮水平下F1代均高于亲本,而大多数性状均值在高氮水平下亲本高于F1。从氮代谢相关性状在低氮水平下后代优于亲本的这一表现得出,氮代谢相关性状适于在适量低氮水平下对优良后代进行选择。
2.2 氮代谢相关性状的遗传方差及遗传力分析
由表2可知,10个性状均存在加性效应且达1%显著水平。株高、籽粒氮素积累量、氮素吸收总量和氮素生理效率的加性效应和显性效应均达1%显著水平,但加性效应所占比重较大,表明以加性效应为主。成熟期单茎干物重加性效应达1%显著水平,显性效应达5%显著水平,表明主要受加性效应控制。这说明既可以通过选择加以固定培育出优良品种,也可以通过利用杂种优势选育氮高效后代。谷氨酰胺合成酶活性、开花期单茎干物重和氮素积累量、花后氮素同化量和氮素利用效率只检测到加性效应且加性方差达极显著水平。基因型×环境的互作对各性状的变异也有重要作用。所有性状的显性×环境互作均达显著或极显著水平,说明各氮代谢相关性状的后代优势因氮水平的不同而呈现较大差异。对于加性×环境互作,只有开花期氮素积累量、氮素利用效率和氮素生理效率的加性×环境互作效应方差达显著水平,其他性状均不存在加性×环境互作效应。
由表2可以看出,各性状狭义遗传力总体不高(平均值为0.56)。其中,最高的是开花期氮素积累量,其次是株高,谷氨酰胺合成酶活性的狭义遗传力最低,仅有0.07。氮素利用效率和氮素生理效率的互作狭义遗传力大于普通狭义遗传力,开花期氮素积累量以普通狭义遗传力为主,但也存在互作狭义遗传力且达到极显著水平。表明这些性状的遗传表达在不同氮水平表现不一致,在不同氮水平下对这些性状的选择效果不同。其他7个性状的互作狭义遗传力均为0,说明对这些性状进行选择时不受氮水平条件的影响。各性状广义遗传力总体较高(平均值为0.881),其中,开花期氮素积累量和株高最高(均为0.92),氮素生理效率最小(0.65)。10个性状的互作广义遗传力均达到1%显著水平,说明不同的氮水平对遗传表达有较大影响。
2.3 氮代谢相关性状的遗传效应预测值
2.3.1 加性效应预测值 从表3可以看到,亲本JN10的籽粒氮素积累量、氮素利用效率和氮素生理效率均最高且达到极显著水平,因此与之杂交的后代籽粒氮素积累量较多,氮素利用效率和氮素生理效率较高。亲本DK138氮素利用效率和氮素生理效率的加性效应表现为显著的正效应,可作为亲本以提高后代的氮素利用效率和氮素生理效率。LM14的开花期单茎干物重、成熟期单茎干物重、开花期氮素积累量、氮素吸收总量和花后氮素同化量均为最大值,说明以LM14作为亲本得到的杂交后代,上述性状均较优,利于优良品种选育。亲本LF6010的多数性状效应值为负值,说明其后代性状多出现衰退现象。

表2 小麦氮代谢相关性状遗传方差和遗传力

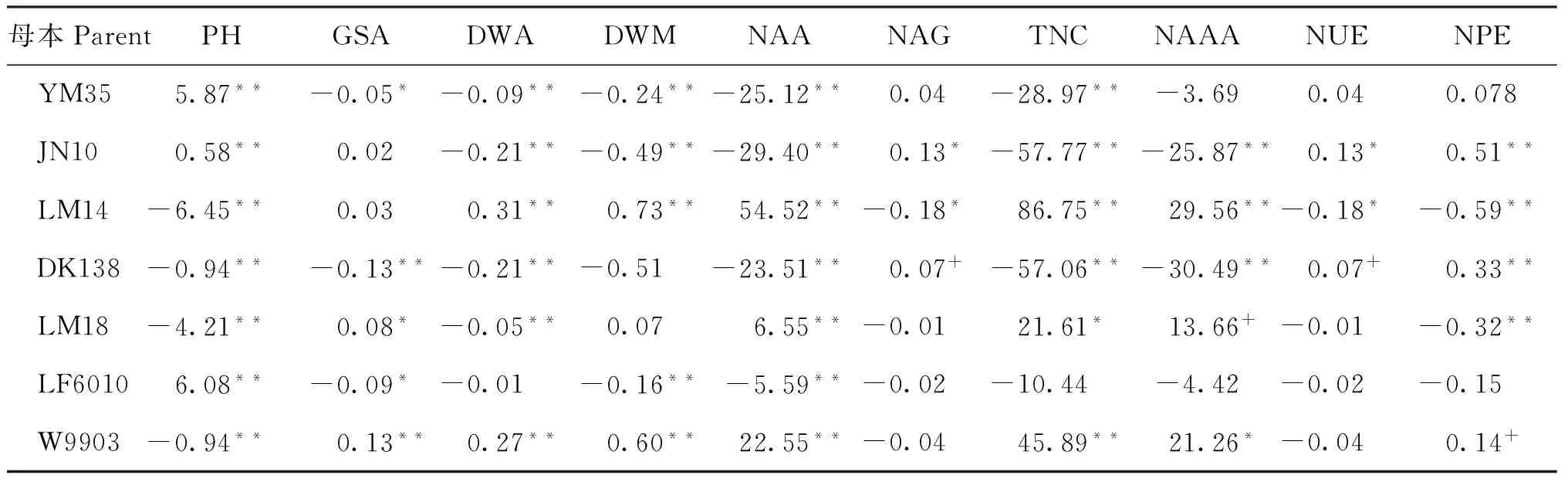
表3 基因型×环境互作下氮代谢相关性状的加性效应预测值
注(Note): PH—株高 Plant height; GSA—谷氨酰胺合成酶活性 Glutamine synthetase activity; DWA—开花期单茎干物重 Dry weight per stem at the anthesis; DWM—成熟期单茎干物重 Dry weight per stem at the maturity stage; NAA—开花期单茎氮素积累量 Nitrogen accumulation per stem at the anthesis; NAG—籽粒氮素积累量 Nitrogen accumulation in grain; TNC—氮素吸收总量 Total nitrogen content; NAAA—花后氮素同化量 Nitrogen assimilation amount after the anthesis; NUE—氮素利用效率Nitrogen use efficiency; NPE—氮素生理效率 Nitrogen physiology efficiency. “+”, “*”, “**”分别表示10%、5%和1%的显著水平Significant at the 10%, 5%, and 1% probability levels, respectively.
2.3.2 显性效应预测值 表4列出了12个杂交组合的显性效应预测值。组合LM14×W9903的成熟期单茎干物重、籽粒氮素积累量和氮素吸收总量的显性效应值均为最大,表明该组合利于这3个性状的后代选择。12个组合的成熟期单茎干物重显性效应值均未达到显著水平。组合YM35×DK138的籽粒氮素积累量显性预测值达5%显著水平。组合JN10×W9903的氮素生理效率显性效应值最大且达到显著水平,说明该组合氮素生理效率较高。

表4 小麦12个杂交组合部分性状的显性效应预测值
注(Note): PH—株高 Plant height; DWM—成熟期单茎干物重 Dry weight per stem at the maturity stage; NAG—籽粒氮素积累量 Nitrogen accumulation in grain; TNC—氮素吸收总量 Total nitrogen content; NPE—氮素生理效率 Nitrogen physiology efficiency. “+”, “*”, “**”分别表示10%、5%和1%的显著水平Significant at the 10%, 5%, and 1% probability levels, respectively.
2.3.3 显性×环境互作效应预测值 从10个性状的显性×环境互作效应预测值(表5)可知,对于株高性状,组合YM35×LF6010的显性主效应为显著的正效应(4.76),在N2和N3水平的显性×环境互作效应预测值也均为显著的正效应(4.08、1.00)。组合LM14×W9903表现负向显性主效应, 在N2和N3水平下具有负向显性互作效应且达到1%显著水平。该组合的株高在正常氮水平和高氮水平下可以稳定遗传。组合YM35×DK138籽粒氮素积累量的显性主效应为显著的正效应(1.24),在N1水平下的显性×环境互作效应预测值为正向效应且达10%显著水平(22.44)。组合LM14×LM18的氮素吸收总量表现正向显性主效应,在N2水平下具有极显著的正向显性互作效应,其他氮水平没有达到显著的显性互作效应。
2.4 氮代谢相关性状间的遗传相关分析
了解性状间的遗传相关性对于多个性状的同步改良或进行间接选择有重要意义。表6显示,谷氨酰胺合成酶活性与其他性状的表现型和基因型相关系数较小,都在0.16以下。开花期单茎干物重、成熟期单茎干物重、开花期氮素积累量、籽粒氮素积累量和氮素吸收总量两两之间的表现型和基因型相关系数均较高,为0.6以上且均达极显著水平。成熟期单茎干物重、籽粒氮素积累量、氮素吸收总量三个性状与花后氮素同化量的表现型和基因型相关系数也较高(0.6以上)。氮素利用效率与氮素生理效率之间的表现型和基因型相关系数较大(0.688、0.858),达到1%显著水平。
由表7可知,10个性状间的遗传相关以加性遗传相关为主。氮素生理效率与株高呈加性正相关关系,达到10%显著水平,与其他性状呈负相关。开花期单茎干物重、成熟期单茎干物重、开花期氮素积累量、籽粒氮素积累量、氮素吸收总量和花后氮素同化量之间的相关系数为正相关且相关系数较高。除株高和谷氨酰胺合成酶活性外,氮素利用效率与其他性状间以显性环境互作相关为主。氮素利用效率与氮素生理效率之间的显性×环境互作相关系数达到10%显著水平。
3 讨论
小麦氮代谢相关性状如株高、开花期氮素积累量和氮素利用效率等都属于数量性状,受微效多基因控制,并且易受不同环境条件的影响。Novaes等[24]的研究鉴定了两个氮水平下调节生长的基因位点,在对20个性状的分析中鉴定出了63个数量性状位点,发现多数数量性状位点对其中一个氮水平有显著的氮素依赖遗传控制。Xu等[25]研究了182个小麦重组自交系在6个环境下的数量性状QTLs,通过对无条件和有条件的田间试验分析氮磷肥对QTLs表达的影响,无条件分析检测到30个基因位点,有条件分析检测到117个基因位点,两种方法检测到的共同基因位点为21个。以往对氮代谢相关性状的遗传分析,如田松等[26]分析了7个茄子自交系氮素利用效率相关性状的配合力,通过计算一般配合力方差与特殊配合力方差的比值来分析性状主要受何种效应基因控制。但尚未更深入的分析氮代谢相关性状的遗传相关性。也有的利用Hayman[27]的双列杂交分析方法估算遗传方差分量和遗传力。对基因型×环境互作效应研究大多是对农艺性状[28-30]的分析,对氮代谢相关性状的研究尚少。本研究利用QGAstation2.0软件进行基因型×环境互作的模型分析,方便地计算出每个亲本氮代谢相关性状的加性效应预测值。亲本的加性效应能够在世代间稳定遗传,参照这些预测值,根据育种目标可以更好的进行亲本组配。同时得出广义遗传力和狭义遗传力及其与环境的互作效应,方便指导育种工作中对优良后代的选择。


表6 小麦氮代谢相关性状间的表现型和基因型相关系数
注(Note): PH—株高 Plant height; GSA—谷氨酰胺合成酶活性 Glutamine synthetase activity; DWA—开花期单茎干物重 Dry weight per stem at the anthesis; DWM—成熟期单茎干物重 Dry weight per stem at the maturity stage; NAA—开花期单茎氮素积累量 Nitrogen accumulation per stem at the anthesis; NAG—籽粒氮素积累量 Nitrogen accumulation in grain; TNC—氮素吸收总量 Total nitrogen content; NAAA—花后氮素同化量 Nitrogen assimilation amount after the anthesis; NUE—氮素利用效率Nitrogen use efficiency; NPE—氮素生理效率 Nitrogen physiology efficiency; 右上角为表现型相关系数(rP)Data in the upper right corner are the phenotypic correlation coefficient(rP); 左下角为基因型相关系数(rG)Data in the lower left corner are the genotypic correlation coefficient(rG). “+”, “*”, “**”分别表示10%、5%和1%的显著水平 Significant at the 10%, 5%, and 1% probability levels, respectively.
本研究采用了基因型×环境互作的加性-显性遗传模型[22],估算遗传方差的同时预测了遗传效应值,还分析了各性状的遗传效应与不同氮水平之间的互作效应。研究发现,氮代谢相关10个性状加性效应均达1%显著水平,以加性效应控制为主。氮素利用效率和氮素生理效率主要受加性×氮环境互作效应控制。与郝西[16]的研究基本一致。由于我国环境条件的多样性,对杂交后代的选择应注意结合特定的环境条件进行。氮代谢相关性状适于在适量低氮水平下对优良后代进行选择。朱军[22]提出将狭义遗传率划分为普通狭义遗传率和互作狭义遗传率。普通狭义遗传率是指有累加性遗传效应的方差占表现型方差的比率;互作狭义遗传率是指有累加性的基因效应与环境互作的方差占表现型方差的比率。对于选择而言,普通狭义遗传率和互作狭义遗传率都是有效的。不同性状的遗传力不同,总体广义遗传力较高,而狭义遗传力较低。开花期氮素积累量和株高的两种遗传力均较高,有利于早代选择。亲本LM14的多数性状加性效应预测值均较高,利于氮高效品种选育。杂交组合LM14×W9903除氮素生理效率外其他性状的显性效应值均最高,表现出良好的后代选育利用潜力。各性状的显性主效应及其在3种氮水平的互作效应在方向上表现不一致,说明小麦氮高效杂交后代的选择宜考虑特定的氮水平条件,在不同的氮水平条件下,小麦氮高效后代的选育效果会出现不同。

表7 小麦氮代谢相关性状间的遗传相关系数
注(Note): PH—株高 Plant height; GSA—谷氨酰胺合成酶活性 Glutamine synthetase activity; DWA—开花期单茎干物重 Dry weight per stem at the anthesis; DWM—成熟期单茎干物重 Dry weight per stem at the maturity stage; NAA—开花期单茎氮素积累量 Nitrogen accumulation per stem at the anthesis; NAG—籽粒氮素积累量 Nitrogen accumulation in grain; TNC—氮素吸收总量 Total nitrogen content; NAAA—花后氮素同化量 Nitrogen assimilation amount after the anthesis; NUE—氮素利用效率Nitrogen use efficiency; NPE—氮素生理效率 Nitrogen physiology efficiency. 右上角上行为加性相关系数(rA),下行为显性相关系数(rD)Data in the upper and lower line in the upper right corner are the additive correlation coefficient(rA) and dominance correlation coefficient(rD); 左下角上行为加性×环境互作相关系数(rAE),下行为显性×环境互作相关系数(rDE) Data in the upper and lower line in the lower left corner are the additive × environmental interaction correlation coefficient(rAE) and dominance × environmental interaction correlation coefficient(rDE). “+”, “*”, “**”分别表示10%、5%和1%的显著水平Significant at the 10%, 5%, and 1% probability levels, respectively.
由相关性分析发现,无论是低氮还是高氮条件,小麦氮素利用效率与籽粒氮浓度呈负相关[31]。本研究表明,小麦氮素利用效率与氮素生理效率的表现型和基因型为正相关且达1%显著水平,同时这两个性状的显性环境互作相关系数达到10%显著水平,因此可以对这两个性状进行协同改良,但在不同环境下改良结果有一定的差异。氮素利用效率与株高也呈正相关,相关系数较低,但达到5%显著水平,可以对这两个性状进行协同选择。而匡艺等[32]研究的小黑麦氮素利用效率与株高呈现1%显著的正相关性,结果基本一致。在杂交后代选择中可被固定和累加的是加性效应,因此加性相关有利于间接选择[33]。本研究中,两两性状间以加性遗传相关为主。氮素利用效率与氮素生理效率、株高、谷氨酰胺合成酶活性的加性相关系数为正值,但只有与氮素生理效率的相关性达到显著水平。通过相关系数分析,发现株高与氮素生理效率加性相关达到10%显著水平,一定程度上可以将株高作为氮素生理效率的间接选择性状,但是未发现可作为氮素利用效率间接选择的性状,有待进一步研究。
4 结论
株高、开花期单茎干物重、成熟期单茎干物重、开花期氮素积累量、籽粒氮素积累量和氮素吸收总量的遗传以加性效应为主,狭义遗传力较高,达到极显著水平,可在早代对这些性状进行选择。花后氮素同化量受显性×环境互作效应影响较大,狭义遗传力较低。氮素利用效率、氮素生理效率主要受加性×环境互作效应控制。品种DK138和JN10可作为亲本以提高后代的氮素利用效率和氮素生理效率。杂交组合LM14×W9903表现出良好的后代选育利用潜力。小麦氮高效杂交后代的选择宜考虑特定的氮环境条件,适量低氮水平适合优良后代的选择。遗传相关分析表明,两两性状间以加性遗传相关为主。一定程度上可以将株高作为氮素生理效率的间接选择性状。氮素利用效率与氮素生理效率的表现型和基因型相关系数为正值且达1%显著水平,可以对这两个性状进行协同改良。
[1] 张福锁. 土壤与植物营养研究新动态(第一卷)[A]. 北京: 北京农业大学出版, 1992. Zhang F S. New developments of soil and plant nutrition research(Volume 1) [A]. Beijing: Beijing Agricultural University Press, 1992.
[2] 刘强, 宋海星, 荣湘民, 等. 不同品种油菜子粒产量及氮效率差异研究[J]. 植物营养与肥料学报, 2009, 15(4): 898-903. Liu Q, Song H X, Rong X Metal. Studies on oilseed yield and nitrogen efficiency in different cultivars of oilseed rape(Brassicanapus)[J]. Plant Nutrition and Fertilizer Science, 2009, 15(4): 898-903.
[3] Mi G H, Liu J A, Zhang F S. Analysis on agronomic efficiency and its components of maize[J]. Journal of China Agricultural University, 1998, 3: 97-104.
[4] Broadbent F E, De Datta S K, Laureles E V. Measurement of nitrogen utilization efficiency in rice genotypes[J]. Agronomy Journal, 1987, 79: 787-791.
[5] Cox M C, Qualset C O, Rains D W. Genetic variation in nitrogen assimilation and translocation[J]. Crop Science, 1985, 25: 430-435.
[6] Sinebo W, Gretzmacher R, Edelbauer A. Genotypic variation for nitrogen use efficiency in ethiopian barley[J]. Field Crops Research, 2004, 85: 43-60.
[7] Barraclough P B, Howarth J R, Jones Jetal. Nitrogen efficiency of wheat: Genotypic and environmental variation and prospects for improvement[J]. European Journal of Agronomy, 2010, 33: 1-11.
[8] 王庆仁, 李继云, 李振声. 不同磷肥对石灰性土壤磷效率小麦基因型生长发育的影响[J]. 土壤通报, 2000, 31(3): 127-129. Wang Q R, Li J Y, Li Z S. The influence of development of wheat genotypes grown on calcareous soil with different phosphate fertilizer[J]. Chinese Journal of Soil Science, 2000, 31(3): 127-129.
[9] 王月福, 于振文, 李尚霞, 等. 氮素营养水平对冬小麦氮代谢关键酶活性变化和籽粒蛋白质含量的影响[J]. 作物学报, 2002, 28(6): 743-748. Wang Y F, Yu Z W, Li S Xetal. Effect of nitrogen nutrition on the change of key enzyme activity during the nitrogen metabolism and kernel protein content in winter wheat[J]. Acta Agronomica Sinica, 2002, 28(6): 743-748.
[10] 吕鹏, 张吉旺, 刘伟, 等. 施氮时期对高产夏玉米氮代谢关键酶活性及抗氧化特性的影响[J]. 应用生态学报, 2012, 23(6): 1591-1598. Lü P, Zhang J W, Liu Wetal. Effects of nitrogen application period on the nitrogen metabolism key enzymes activities and antioxidant characteristics of high-yielding summer maize[J]. Chinese Journal of Applied Ecology, 2012, 23(6): 1591-1598.
[11] 周桃旺. 低氮胁迫下玉米杂交子一代遗传特性的研究[J]. 安徽农学通报, 2013, 19(10): 24-25. Zhou T W. The study of generation genetic for maize hybrids under low nitrogen stress[J]. Anhui Agricultural Science Bulletin, 2013, 19(10): 24-25.
[12] 朴钟泽, 韩龙植, 高熙宗, 等. 水稻干物质量和氮素利用效率性状的配合力分析[J]. 中国水稻科学, 2005, 19(6): 527-532. Piao Z Z, Han L Z, Gao X Zetal. Analysis on combining ability of dry weight and nitrogen use efficiency in rice[J]. Chinese Journal of Rice Science, 2005, 19(6): 527-532.
[13] 李法计, 常鑫, 王宇娟, 等. 小麦重组自交系群体9个重要农艺性状的遗传分析[J]. 麦类作物学报, 2013, 33(1): 23-28. Li F J, Chang X, Wang Y Jetal. Genetics analysis of nine important agronomic traits in wheat population of recombinant inbred lines[J]. Journal of Triticeae Crops, 2013, 33(1): 23-28.
[14] 刘长友, 范保杰, 曹志敏, 等. 利用混合线性模型分析绿豆主要农艺性状的遗传及相关性[J]. 作物学报, 2012, 38(4): 624-631. Liu C Y, Fan B J, Cao Z Metal. Inheritance and correlation analysis of main agronomic traits in mungbean(VignaradiataL.) by mixed linear models[J]. Acta Agronomica Sinica, 2012, 38(4): 624-631.
[15] 左清凡, 朱军, 刘宜柏, 等. 非等试验设计植株农艺及产量性状的数量遗传分析方法[J]. 中国农业科学, 2000, 33(2): 30-33. Zuo Q F, Zhu J, Liu Y Betal. Methods of quantitative genetic analysis of plant agronomic traits and yield traits for unequal experiment design[J]. Scientia Agricultura Sinica, 2000, 33(2): 30-33.
[16] 郝西. 小麦氮磷效率相关性状遗传规律的研究[D]. 郑州: 河南农业大学硕士学位论文, 2004. Hao X. Studies on genetic principles of wheat traits relevant to nitrogen and phosphorus efficiency[D]. Zhengzhou: PhD Dessertaton of Henan Agricultural University, 2004.
[17] Griffing B. Concept of general and specific combining ability in relation to diallel crossing systems[J]. Australian Journal of Biological Sciences, 1956, 9: 463-493.
[18] 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000. Bao S D. Soil and agricultural chemistry analysis [M]. Beijing: China Agricultural Press, 2000.
[19] 宋海星, 彭建伟, 刘强, 等. 不同氮素生理效率油菜生育后期氮素再分配特性研究[J]. 中国农业科学, 2008, 41(6): 1858-1864. Song H X, Peng J W, Liu Qetal. Nitrogen redistribution characteristics of oilseed rape varieties with different nitrogen physiological efficiency during later growing period[J]. Scientia Agricultura Sinica, 2008, 41(6): 1858-1864.
[20] 朱军, 季道藩, 许馥华. 作物品种间杂种优势遗传分析的新方法[J]. 遗传学报, 1993, 20(3): 262-271. Zhu J, Ji D F, Xu F H. A genetic approach for analyzing intra-cultivar heterosis in crops[J]. Acta Genetica Sinica, 1993, 20(3): 262-271.
[21] Zhu J, Weir B S. Diallel analysis for sex-linked and maternal effects[J]. Theoretical and Applied Genetics, 1996, 92(1): 1-9.
[22] 朱军. 数量性状遗传分析的新方法及其在育种中的应用[J]. 浙江大学学报(农业与生命科学版), 2000, 26(1): 1-6. Zhu J. New approaches of genetic analysis for quantitative traits and their applications in breeding[J]. Journal of Zhejiang University(Agriculture & Life Sciences), 2000, 26(1): 1-6.
[23] 朱军. 遗传学[M]. 北京: 中国农业出版社, 2008. 316-324. Zhu J. Genetics [M]. Beijing: China Agriculture Press, 2008. 316-324.
[24] Novaes E, Osorio L, Drost D Retal. Quantitative genetic analysis of biomass and wood chemistry ofPopulusunder different nitrogen levels[J]. New Phytologist, 2009, 182: 878-890.
[25] Xu Y F, Wang R F, Tong Y Petal. Mapping QTLs for yield and nitrogen-related traits in wheat: influence of nitrogen and phosphorus fertilization on QTL expression[J]. Theoretical and Applied Genetics, 2013.Doi: 10.1007/s00122-013-2201-y
[26] 田松, 银婷, 王彦华, 等. 茄子氮素利用效率相关性状的配合力分析[J]. 植物遗传资源学报, 2011, 12(3): 481-485. Tian S, Yin T, Wang Y Hetal. Combination ability analysis of traits related to nitrogen use efficiency in eggplant[J]. Journal of Plant Genetic Resources, 2011, 12(3): 481-485.
[27] Hayman B I. The theory and analysis of diallel crosses[J]. Genetics, 1954, 39: 789-809.
[28] 肖炳光, 朱军, 卢秀萍, 等. 烤烟主要农艺性状的遗传与相关分析[J]. 遗传, 2006, 28(3): 317-323. Xiao B G, Zhu J, Lu X Petal. Genetic and correlation analysis for agronomic traits in flue-cured tobacco(NicotianatabacumL.)[J]. Hereditas, 2006, 28(3): 317-323.
[29] 王坚, 赵开军, 乔枫, 杨生龙.OsGA20ox2不同长度RNAi片段对水稻株高等农艺性状的遗传效应[J]. 作物学报, 2012, 38(4): 632-638. Wang J, Zhao K J, Qiao F, Yang S Letal. Genetic effects of different RNA interference fragments fromOsGA20ox2 on plant height and other agronomic traits in rice[J]. Acta Agronomica Sinica, 2012, 38(4): 632-638.
[30] 卢超, 高明博, 焦小钟, 等. 几个小麦亲本主要农艺性状的配合力评价及遗传力分析[J]. 麦类作物学报, 2010,30(6): 1023-1028. Lu C, Gao M B, Jiao X Zetal. Combining ability and heritability analysis of main agronomic traits in wheat[J]. Journal of Triticeae Crops, 2010, 30(6): 1023-1028.
[31] 李雁鸣, 张建平, 王焕忠, 等. 喷灌条件下高产冬小麦植株体内氮磷钾的时空分配特点[J]. 河北农业大学学报, 2002, 25(3): 1-5. Li Y M, Zhang J P, Wang H Zetal. Properties of spatial and temporal distribution of nitrogen, phosphorus and potassium in the plant of high yield winter wheat under the conditions of sprinkling irrigation[J]. Journal of Agricultural University of Hebei, 2002, 25(3): 1-5.
[32] 匡艺, 李廷轩, 张锡洲, 等. 小黑麦氮利用效率基因型差异及评价[J]. 植物营养与肥料学报, 2011, 17(4): 845-851. Kuang Y, Li T X, Zhang X Zetal. Variation of nitrogen use efficiency of triticale in genotype and its evaluation[J]. Plant Nutrition and Fertilizer Science, 2011, 17(4): 845-851.
[33] 吴吉祥, 朱军, 许馥华, 等. 陆地棉产量性状的遗传效应及其与环境互作的分析[J]. 遗传, 1995, 17(5): 1-4. Wu J X, Zhu J, Xu F Hetal. Analysis of genetic effect × environment interactions for yield traits in upland cotton[J]. Hereditas, 1995, 17(5): 1-4.
Inheritance and correlation analysis of genotype × environment interaction for nitrogen metabolism-related traits of wheat
WANG Lin-lin1, WANG Ping2, WANG Zhen-lin1, SUN Ai-qing1, YANG Min1, WANG Chun-wei1, YIN Yan-ping1*
(1CollegeofAgronomy,ShandongAgriculturalUniversity/StateKeyLaboratoryofCropBiology/ShandongKeyLaboratoryofCropBiology,Tai’an,Shandong271018,China; 2Tai’anAcademyofAgriculturalSciences,Tai’an,Shandong271000,China)
【Objectives】 Selection of wheat cultivars of high nitrogen utilization is essential for the improvement of nitrogen fertilizer use efficiency and environmental safety. In this study, the genetic effects of nitrogen metabolism-related traits of wheat were analyzed, to provided a theoretical basis for wheat breeding of high nitrogen use efficiency cultivars. 【Methods】 Seven wheat cultivars with different nitrogen use efficiencies and their twelve F1crosses were used as materials in a pot experiment. Three nitrogen levels were designed, a genetic model was used to calculate the additive and dominance effects and their interaction. 【Results】 The additive effects are the major genetic component for plant height, the dry weights in single stem at the anthesis stage and maturity stage, the nitrogen accumulation in anthesis stage, the nitrogen accumulation in grain and the total nitrogen contents. The total N assimilation amounts after the anthesis are mainly controlled by dominance × environment interaction effects, and the nitrogen use efficiency and nitrogen physiological efficiency mainly by additive × environment interaction effects. The heritabilities of ten traits in narrow-sense are generally small with a mean of 0.56, but are high in broad-sense with a mean of 0.881. The interaction between the heritability of ten traits in broad-sense is significant at 1%, showing a intense effect on the genetic expression under different nitrogen levels. The narrow-sense heritability interactions for the nitrogen use efficiency, nitrogen physiological efficiency and nitrogen assimilation amount after the anthesis are high, showing the different selection results of nitrogen levels to the wheat traits. The additive effect forecast shows the significant positive interaction of DK138 and JN10 in N use efficiency and N use physiological efficiency. The dominant effects in most crosses do not always show agreeable directions with those in the interactions of domestic × environmental factors, which indicate that the selection of wheat traits with high N efficiencies should consider the supply levels of nitrogen. The dominance effects of the nitrogen physiology efficiency on cross JN10×W9903 are maximum and reach significant level, showing high nitrogen physiology efficiency. The cross LM14×W9903 exhibits expecting potential for the selection of desirable progenies according the values of dominance effect. The correlation analysis indicates the predominance effects of additive correlation among traits. The nitrogen physiological efficiency and plant height show positive additive correlations which reach the significant level at 10%. Except the plant height and glutamine synthetase activity, the nitrogen use efficiency and other traits are given priority to dominant × environment interaction correlation. The correlations between nitrogen use efficiencies and phenotypic and genotypic nitrogen physiological efficiencies are positive significantly. 【Conclusions】 The plant height can be used as indirect selection traits of nitrogen physiological efficiency to a certain extent. Both the genotypic nitrogen use efficiency and nitrogen physiological efficiency traits could be improved simultaneously. The DK138 and JN10 can be used as parents to breeding cross progenies with high nitrogen use efficiency and nitrogen physiological efficiency. Cross LM14×W9903 has good potential of selecting progenies according to the values of dominance effect.
wheat(TriticumaestivumL.); nitrogen metabolism; inheritance; additive-dominance genetic model; genotype × environment interaction
2014-02-27 接受日期: 2014-08-09 网络出版日期: 2015-02-12
国家公益性行业(农业)科研专项(201203029, 201303002)资助。
王琳琳(1989—),女,河北邢台人,硕士, 主要从事作物遗传育种方面的研究。 E-mail: wanglinlin20120128@163.com * 通信作者 E-mail: ypyinsdau@sina.com
S512.03
A
1008-505X(2015)03-0549-12
