不同施氮水平苹果矮化中间砧幼树光合产物的周年分配利用
2015-06-15姜远茂魏绍冲葛顺峰李洪娜门永阁
李 晶, 姜远茂, 魏绍冲, 葛顺峰, 李洪娜, 门永阁, 周 乐
(山东农业大学园艺科学与工程学院/作物生物学国家重点实验室,山东泰安 271018)
不同施氮水平苹果矮化中间砧幼树光合产物的周年分配利用
李 晶, 姜远茂*, 魏绍冲, 葛顺峰, 李洪娜, 门永阁, 周 乐
(山东农业大学园艺科学与工程学院/作物生物学国家重点实验室,山东泰安 271018)
【目的】光合产物在树体内的利用、分配状况直接影响着果树的产量形成,是果树优质、丰产、稳产的重要因素。氮肥的不合理施用易导致树体C/N失衡,造成树体旺长或早衰,直接影响果树的产量、品质形成。因此,研究矮化中间砧苹果在不同氮水平下的光合产物利用、分配特性,为合理协调光合产物在树体内的利用、分配以保证果树稳健生长又及时成花结果打下理论基础。【方法】以生产上最常用的2年生烟富3/M26/平邑甜茶幼树为试材进行盆栽试验。设置不施氮肥(N0)、适宜施氮肥(N100)和过量施氮肥(N200) 3个氮素水平,分别于春梢生长期、春梢停长期、秋梢生长期进行13C标记,标记72 h后取样,整株解析为叶、一年生枝条、主干、中间砧、根系,测定了其13C丰度,玉米素核苷(ZR),脱落酸(ABA),可溶性淀粉含量,并测定了叶面积和叶绿素含量。【结果】与N0相比,不同物候期适宜施氮肥(N100)和过量施氮肥(N200)处理均显著促进树体生物量的增加,提高叶片面积和叶绿素含量,N100处理对树体生长的促进作用随着氮肥施入时间的延长逐渐显现。春梢生长期和春梢停长期,N100处理细根生长量最高,其次是N200处理,N0处理最低;至秋梢生长期,N0处理细根生物量迅速升高至最高且显著高于N200处理。N0处理在不同生长期叶片淀粉含量均显著高于N100和N200处理。氮肥施入初期,叶片ZR含量为N200>N100>N0,施肥30天后,N100处理叶片仍保持较高的ZR含量,但N200处理ZR含量显著下降。氮肥施入初期各处理ABA含量无显著差异,随着生育期延长差异性逐渐显著,施肥后30天,N0处理的叶片ABA含量达到最高并保持较高水平至生长后期。不同施氮处理树体根冠比和光合产物分配规律在不同生长期差异显著。氮肥施入至春梢生长期,N100和N200处理根系13C分配率分别是N0处理的285.35%和217.98%,而N0处理树体会将更多的光合产物用于地上部生长;至春梢停长期N100和N200处理仍保持较高根冠比和根系13C分配率;至秋梢生长期,N0处理根系光合产物分配率升高,而N100和N200处理根系13C分配率分别降低至N0处理根系13C分配率的71.98% 和41.26%,表明生长后期N0处理生长中心逐渐向根系转移。【结论】施氮水平对苹果矮化中间砧幼树生长及光合产物利用方式和分配规律的显著影响与玉米素核苷和脱落酸的合成变化密切相关。施氮通过促进ZR大量合成显著促使光合产物向根系大量分配,周年尺度上表现为树体根冠比和根系生物量显著升高,树体地上部快速生长。整个生长期内低氮条件下树体光合产物转化为淀粉在叶片中大量贮存是由ABA的合成差异所造成。
矮化苹果砧木; 氮肥水平; 光合产物; 利用分配
目前我国苹果生产上由于施肥不合理,所谓的高产园也常因缺素或肥料施用不平衡,存在产量不稳定、果品质量差的问题[1]。氮素营养与树体营养生长关系密切,碳水化合物在地上部累积是花芽分化的基础,直接影响着果树的产量、品质形成。栽培过程中氮素偏少树体C/N较高,利于从营养生长向生殖生长转化,但生长缓慢,落花落果严重,树体早衰;过量施氮易引起地上部过旺生长,C/N失调,营养生长与生殖生长争夺养分,落花落果严重[2]。氮肥不足和超量并存等问题[3]导致树体C/N失衡直接影响着果实产量和品质形成,如何通过合理施肥既保证树体稳健生长又及时成花结果是苹果栽培管理的核心。目前对矮砧苹果的碳氮特性研究较少,关于苹果矮化中间砧在不同氮水平下光合产物的年周期利用、分配规律研究尚未见报道。本研究以2年生烟富3/M26/平邑甜茶幼树为试材,采用13C同位素示踪法研究不同施氮水平对苹果矮化中间砧幼树生长及光合产物利用、分配规律的影响,以明确苹果矮化中间砧幼树在不同施氮水平下不同生长期对光合产物的利用及分配规律,以期通过氮肥施用量对树体生长节奏加以调节,维持树体C/N平衡,协调营养生长与生殖生长,在早果丰产的基础上保持优质稳产,为苹果矮化中间砧高产优质栽培配套技术提供理论依据。
1 材料与方法
1.1 试验设计
试验于2011年在山东农业大学园艺试验站进行。试材为定植在园艺试验站的2年生烟富3/M26/平邑甜茶,不同处理间设置隔离树。供试土壤为粘质壤土,有机质10.13 g/kg、 碱解氮76.63 mg/kg、 速效磷27.28 mg/kg、 速效钾184.99 mg/kg、 硝态氮37.95 mg/kg、 铵态氮16.17 mg/kg、 pH 6.7。于2011年4月15日各选取9株生长势基本一致、无病虫害的植株,试验设氮(N)施用量:0、100、200 kg/hm2(以下简称N0、 N100、 N200)三个水平。施肥后立即浇水,进行常规管理,各处理生长条件和其他栽培管理保持一致。分别于2011年6月1日(春梢生长期)、7月1日(春梢停长期)、8月15日(秋梢生长期)进行13C标记,72 h后破坏性采样。各处理均取3株,单株重复3次。
1.2 取样与测定
玉米素核苷(ZR)和脱落酸(ABA)含量的测定采用酶联吸附免疫测定法(ELISA)[4],ELISA试剂盒由中国农业大学提供,在中国农业大学进行。可溶性淀粉的测定采用蒽酮法[5]。叶面积采用手持式YMJ-B活体叶面积仪测定。叶绿素含量采用SPAD502叶绿素仪测定。13C标记72 h后取样,整株解析为叶、一年生枝条、主干、中间砧、根系。样品按清水→洗涤剂→清水→1%盐酸→3次去离子水顺序冲洗后,80℃下烘干至恒重,粉碎后过0.25 mm筛,混匀后装袋备用,13C丰度在DELTAplusXP型质谱仪中测定。
1.3 计算及统计方法
13C计算公式为[6-7]:
Atom%13C =(δ13C + 1000) × RPBD/[(δ13C + 1000) ×RPBD+1000]×100
式中:Atom%为13C丰度;RPDB为碳同位素的标准比值,RPDB=0.0112372。
Ci=干物重(g)×C%
式中:Ci为各器官所含的碳量。
13Ci(mg)= Ci×[(Atom13C%)标记丰度-(Atom13C%)自然丰度]×1000
式中:Ci为各器官所含的碳量;13Ci为各器官13C量。
13Ci%=13Ci/13C净吸收×100
式中:13Ci%为该器官的13C量占树体净吸收13C量的百分比。
所有数据均采用DPS 7.05软件进行单因素方差分析,LSD法进行差异显著性比较,应用Microsoft Excel 2003进行图表绘制。
2 结果与分析
2.1 施氮水平对树体生长的影响
氮肥施入至春梢生长期,施氮肥增加树体生物量,但不同处理间差异不显著;至春梢停长期,N100和N200处理对树体生长的促进作用逐渐显著,N100处理(196.2 g)和N200处理(209.8 g)树体生物量显著升高至N0处理(151.63 g)的125.2%和133.8%,但二者之间无显著性差异;至秋梢生长期,N100处理(351.4 g)和N200处理(323.0 g)仍保持较高树体生物量,分别为N0处理的144.0%和132.3%,二者之间虽无显著性差异,但N100处理对树体生长的促进作用随着氮肥施入时间的延长逐渐显现(图1)。
2.2 施氮水平对根系生长的影响
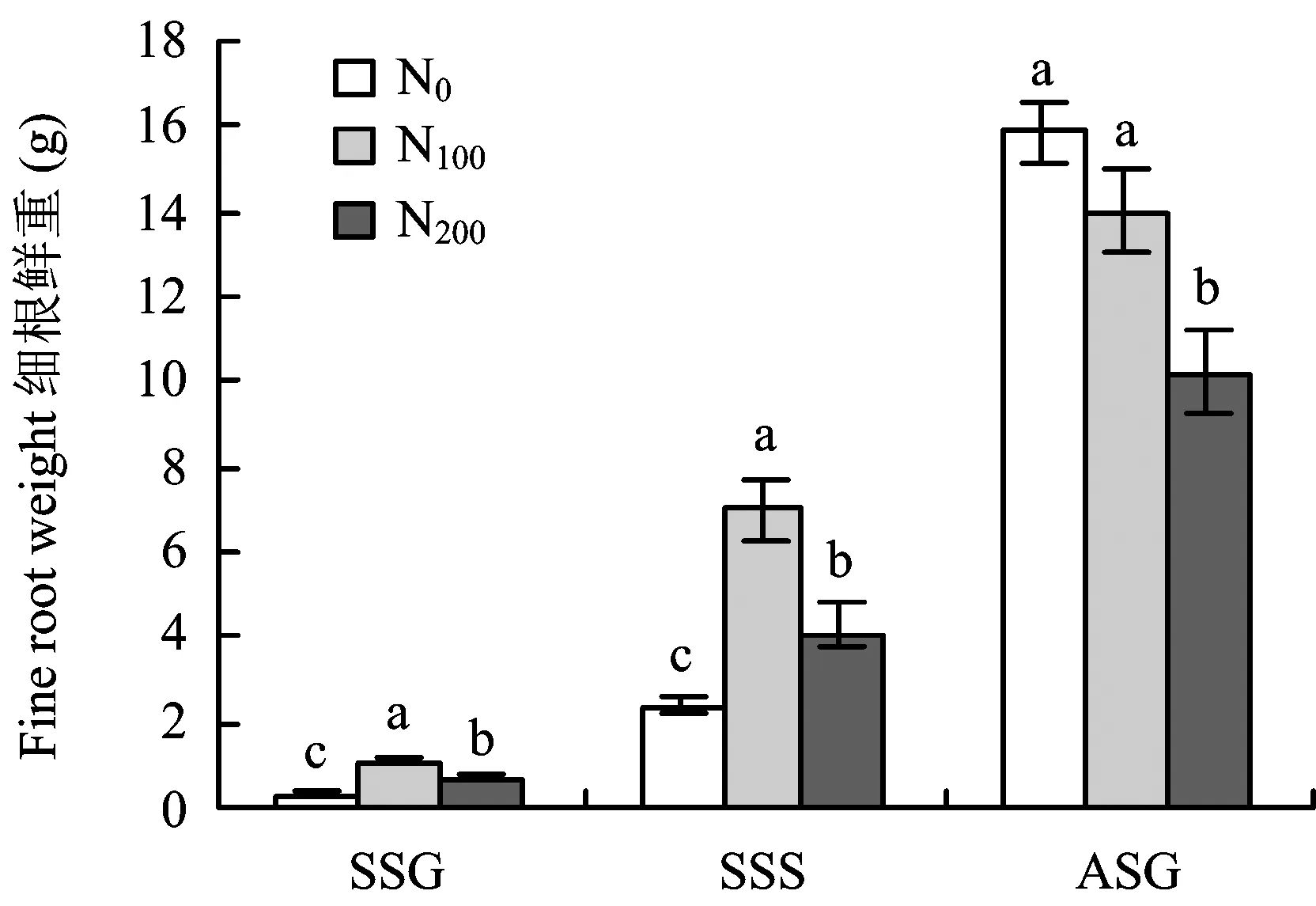
图2 施氮水平对细根生长的影响Fig.2 Effects of different N rates on fine roots growth[注(Note): SSG—春梢生长期 Spring shoot growing, SSS—春梢停止生长期Spring shoot stop growing; ASG—秋梢生长期Autumn shoot growing. 柱上不同字母表示同一时期处理间差异达到5%显著水平Different letters above the bars mean significant differences at the same stage at the 5% level.]
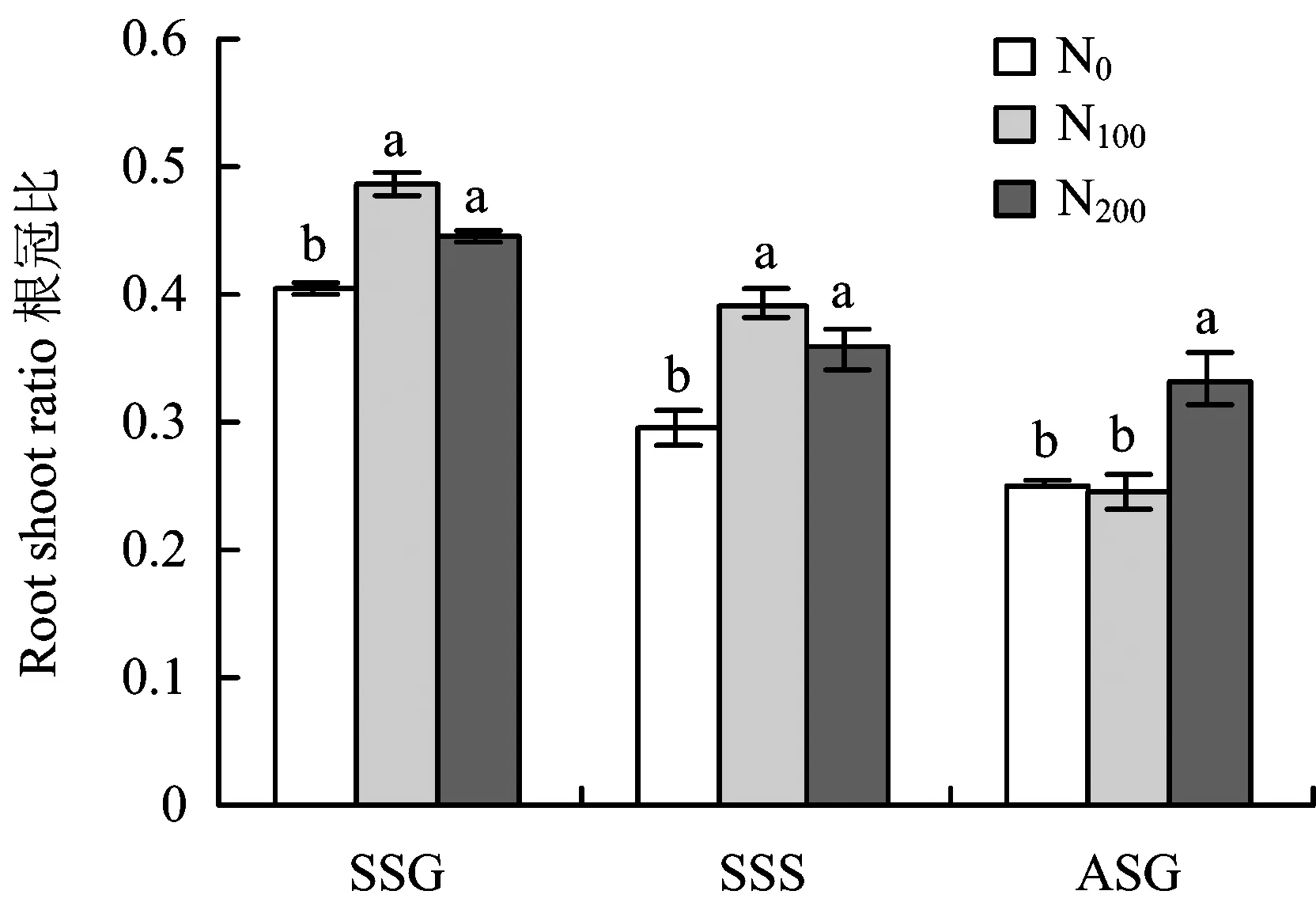
图3 施氮水平对根冠比的影响Fig.3 Effects of different N rates on root to shoot ratio[注(Note): SSG—春梢生长期 Spring shoot growing, SSS—春梢停止生长期Spring shoot stop growing; ASG—秋梢生长期Autumn shoot growing. 柱上不同字母表示同一时期处理间差异达到5%显著水平Different letters above the bars mean significant differences at the same stage at the 5% level.]
图2可见,施氮水平对苹果幼树细根生长影响显著。春梢生长期和春梢停长期,不同施氮处理树体细根生物量差异显著,N100处理细根生长量最高,其次是N200处理,N0处理最低,表明氮肥施入在较长时间内对细根生长量的增加有一定促进作用,且N100施氮水平相对于N200处理更利于细根生长。至秋梢生长期,N0处理细根生物量迅速升高至最高且显著高于N200处理,与N100处理无显著差异。施氮水平对根冠比年周期变化趋势无影响(图3),均表现为随着生长期的延长根冠比逐渐降低,施氮水平只改变不同生长期根冠比大小。氮肥施入初期,N100和N200施氮肥处理至春梢生长期根冠比显著升高至N0处理的119.04%和119.66%,表明施氮可促进根系快速生长,表现为根冠比的快速提高。随着氮肥施入时间和生长期的延长,至春梢停长期各处理根冠比逐渐降低,但N100和N200施氮处理根冠比仍显著高于N0,表明N100和N200氮水平对根系生长的促进作用仍显著高于N0。至秋梢生长期,随着N100处理地上部快速生长,根冠比则持续降低至与N0无显著差异。
2.3 施氮水平对叶片内源激素含量的影响
施氮对树体ZR含量影响显著(图4)。氮肥施入至第3天,叶片ZR含量为N200>N100>N0;生长期延长至施肥30天,N100处理叶片仍保持较高的ZR含量,且显著高于N200和N0处理;至施肥60天,不同施氮处理叶片ZR含量差异性逐渐降低,仍表现为N100>N200>N0;至施肥90天,各处理叶片的ZR含量无显著差异。氮肥施入初期各处理ABA含量无显著差异,随着氮肥施入时间的延长差异性逐渐显著(图5)。至施肥后30天,N0处理的叶片ABA含量呈现升高趋势并保持较高水平至生长后期。施肥30天N100和N200处理的叶片ABA含量显著低于N0,且两者间无显著差异,至施肥60天仍表现为N0>N200>N100。施肥时间延长至90天,叶片ABA含量改变为N0>N100>N200。
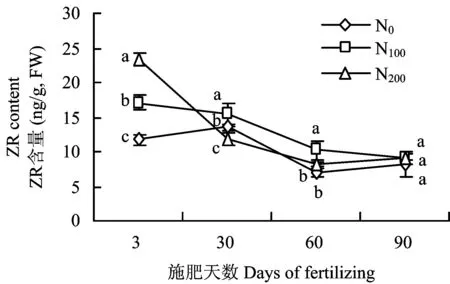
图4 施氮水平对叶片玉米素核苷ZR含量的影响Fig.4 Effects of different N rates on ZR contents in leaves[注(Note): 图中不同字母表示不同处理间在P<0.05水平差异显著 Different letters indicate a significant difference among treatments(P<0.05).]
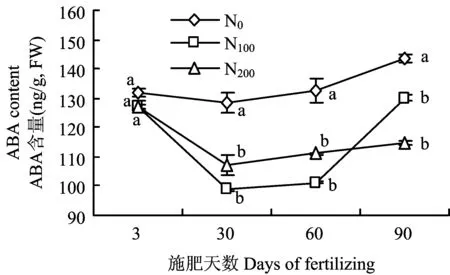
图5 施氮水平对叶片脱落酸ABA含量的影响Fig.5 Effects of different N rates on ABA contents in leaves[注(Note): 图中不同字母表示不同处理间在P<0.05水平差异显著 Different letters indicate a significant difference among treatments(P<0.05).]
2.4 施氮水平对叶片生长的影响
施氮水平对叶片生长影响显著。表1可见,至秋梢生长期,N100和N200处理叶面积分别为32.33 cm2和31.54 cm2,均显著高于N0处理(26.73 cm2),表明施氮利于叶面积的显著提高,但N100和N200处理的叶片面积差异不显著。氮作为叶绿素的组成成分,氮供应浓度与叶绿素含量密切相关。本次试验叶绿素含量(叶片SPAD值)在各物候期均表现为N200>N100>N0,以春梢停长期不同施氮处理间差异性最为显著,表明施氮利于叶绿素含量的提高,但随着施氮量的增加,对叶绿素含量的促进作用也逐渐降低。淀粉是多数植物非结构性碳水化合物的主要贮存形式,作为能量的主要贮存物质对植物的生长发育具有重要作用。氮肥施入至春梢生长期,不同施氮处理叶片淀粉含量差异性最为显著,N0处理叶片淀粉含量分别为N100和N200处理的174.9%和152.3%;随着生长期延长至春梢停长期,不同施氮处理叶片淀粉含量差异程度逐渐降低,但N0处理叶片淀粉含量仍是N100和N200处理的122.8%和148.4%;至秋梢生长期,不同施氮处理叶片淀粉含量的差异程度降低,但N0处理叶片仍表现为较高的淀粉含量,为N100和N200处理的112.4%和121.9%,表明N0处理更多的光合产物转化为淀粉进行贮藏。
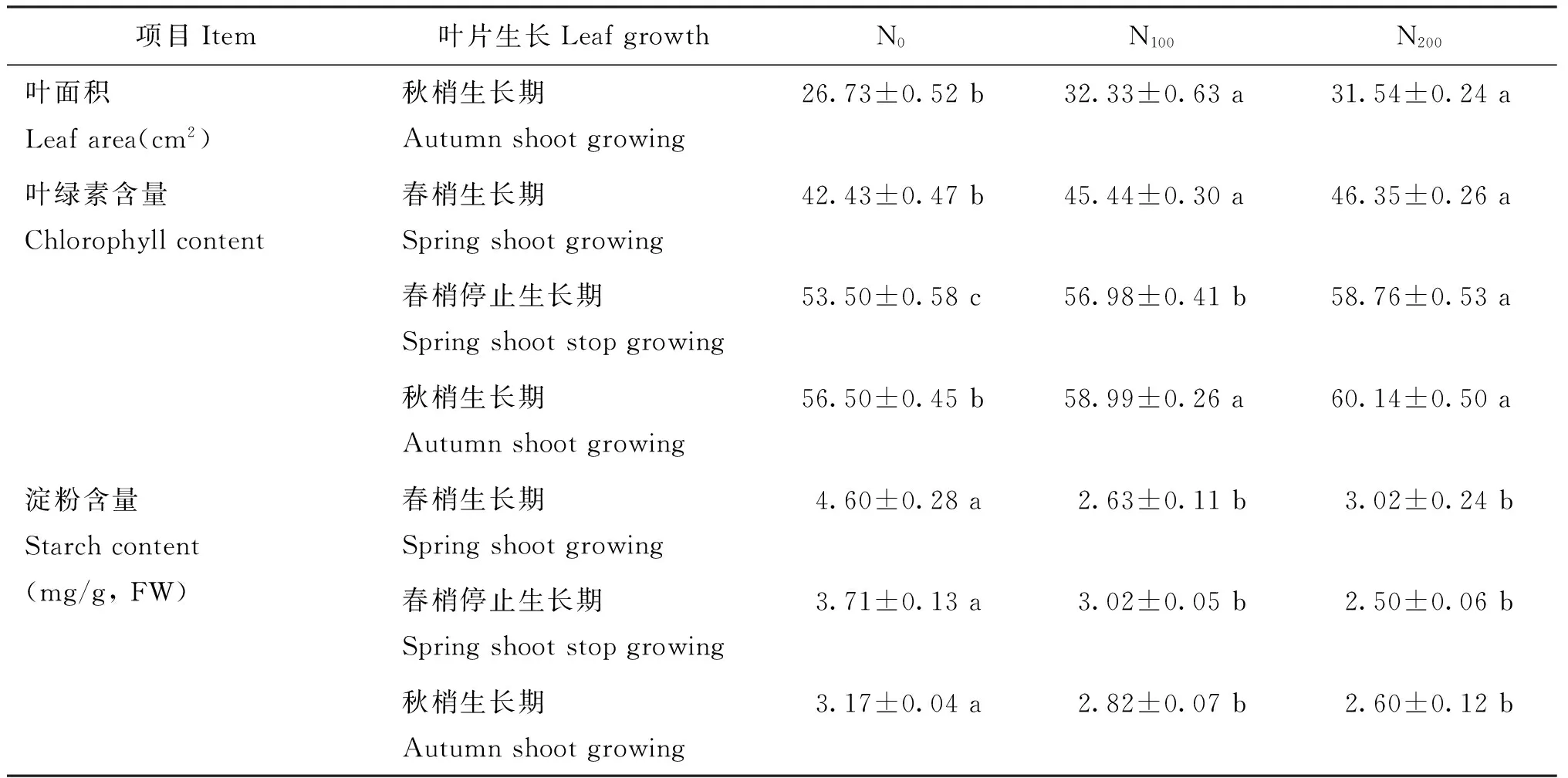
表1 施氮水平对叶片生长的影响
注(Note): 同行不同小写字母表示处理间差异为5%显著水平(n=3)Different letters mean significant differences among the treatments at the same stage at the 5% level.
2.5 施氮水平对年周期13C分配率的影响
13C同化产物分配到各个器官的比率与其竞争能力有关,竞争能力指代谢及生长旺盛部位从标记叶片中吸取13C的能力[7]。施氮水平对不同生长期光合产物在树体内的分配影响显著(表2)。春梢生长期至秋梢生长期,N0处理地上部13C分配率先高后低,根系13C分配率至生长后期显著增高;N100和N200处理则表现为地上部光合产物分配率的逐渐增强和根系光合产物分配率的逐渐降低。
氮肥施入至春梢生长期,低氮N0处理根系的13C分配率仅为10.40%,而N100和N200处理的根系13C分配率分别升高至N0处理的285.35%和217.98%,表明施氮可促进根系光合产物分配率的快速提高,利于光合产物向根系的快速转运,且N100施氮水平对根系光合产物分配率的促进作用较N200更为显著,低氮条件下春梢生长期树体会将更多的光合产物用于地上部生长。至春梢停长期,随着地上部旺盛生长,地上部光合产物分配率增高至81.36%,根系光合产物分配率(9.44%)仍显著低于N100和N200处理,根系13C分配率由高到低依然表现为N100>N200>N0,表明N100和N200处理依然显著促进光合产物向根系的大量运输,且N100处理对根系生长的促进作用较N200更为显著。秋梢生长期N0处理根系光合产物分配率升高至其春梢停长期光合产物分配率的179.91%,且显著高于N100和N200处理,地上部光合产物的分配率降低至与N100无显著差异,表明至生长后期N0处理生长中心逐渐向根系转移。
3 讨论与结论
施氮水平引起的树体生长及光合产物分配、利用差异均与内源激素的合成密切相关。在氮肥施入初期,适宜施氮肥N100处理和过量施氮肥N200处理根冠比(图3)和根系13C分配率(表2)在氮肥施入初期均显著高于N0处理,可能是由于施氮可诱导根中细胞分裂素合成酶基因IPT3快速表达和细胞分裂素大量合成[8-9],根系中细胞分裂素的大量合成对光合产物的运输产生加速和定向作用[10-12],促进了光合产物向根系的分配比率和根系生长。光合产物向根系的大量运输在短时间尺度上表现为显著增高的根系13C分配率(表2),在月或年的长时间尺度上则表现为不同器官光合产物分配积累量的改变[13-14],即根系生物量和根冠比的显著升高。
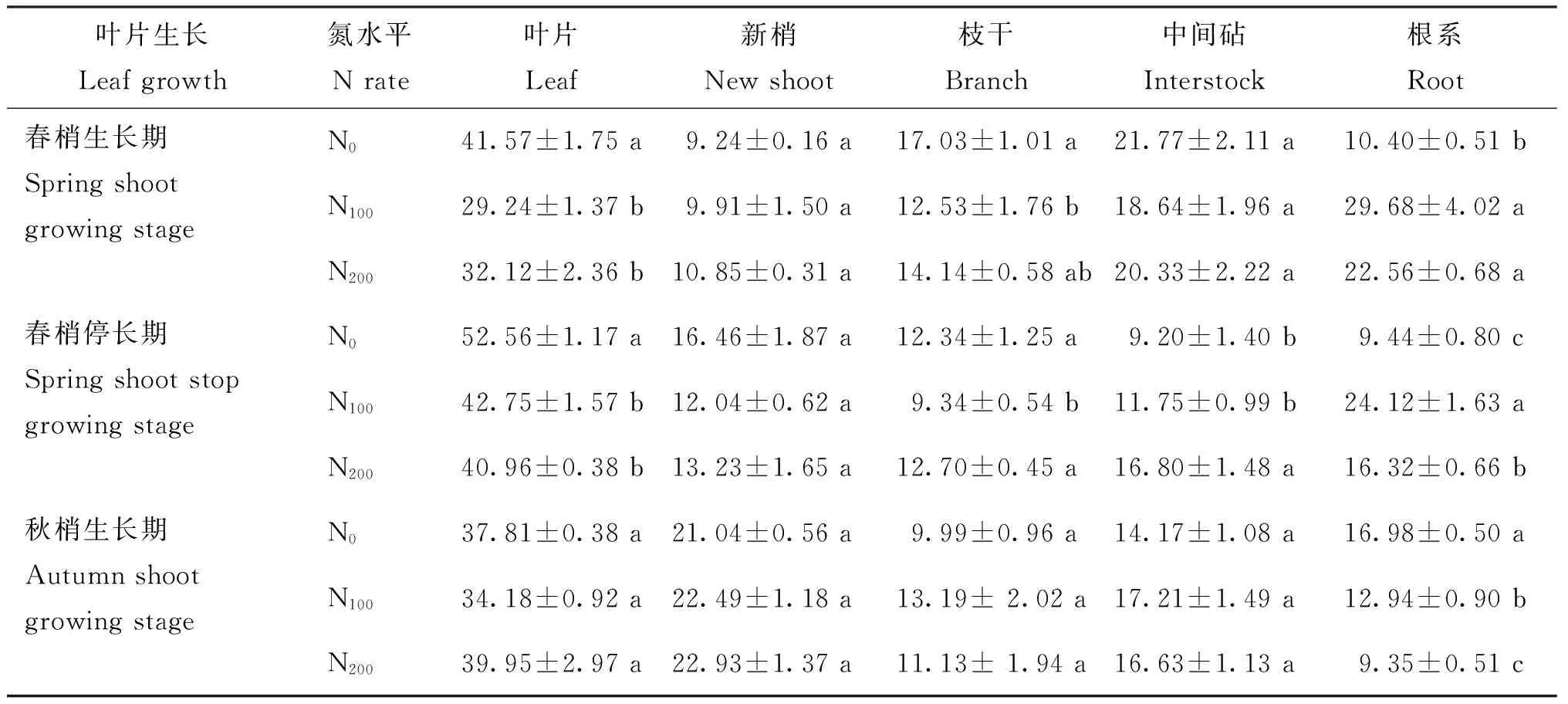
表2 施氮水平对年周期各器官13C分配率的影响
注(Note): 不同字母表示同一时期不同处理间差异为5%显著水平(n=3) Different letters mean significant differences at the 5% level among different treatments at the same stage.

淀粉累积量与果树成花显著相关。本试验整个生长期内,叶片淀粉含量均表现为不施氮肥条件下最高,同时不施氮肥条件下叶片ABA含量也表现为最高,不仅是由于根系生长缓慢降低光合产物极性运输至根系的比率,较高的ABA合成量(图5)对GA产生拮抗,抑制α-淀粉酶合成[20],可能是不施氮肥条件下光合产物以淀粉(表1)的形式在地上部大量累积的主要原因。因此不同氮水平下树体光合产物利用形态的差异是由ABA的合成差异所造成,可能是前人研究认为高淀粉含量和高ABA含量与成花有平行关系的主要原因[2,21]。
综合本次试验及前人研究结果表明,施氮水平对苹果矮化中间砧幼树生长的影响与施氮引起的ZR和ABA合成差异和光合产物的利用和分配差异密切相关。ZR是细胞分裂素(CTK)在木质部运输的主要形式,施氮对树体生长的促进作用是通过诱导ZR的大量合成促进根系光合产物分配率显著提高以促进快速根系生长,长时间尺度上则表现为根冠比和根系的显著升高,进而促进地上部快速生长。低氮条件下光合产物转化为淀粉在叶片中大量贮存是由ABA的大量合成造成。
[1] 丁平海, 郗荣庭, 张玉星, 等. 河北省主要苹果营养状况及施肥设计[J]. 河北农业大学学报. 1994, 17(3): 5-10. Ding P H, Xi R T, Zhang Y Xetal. The nutrition conditions and fertilizer application design of main apple varieties in Hebei Province[J]. Journal of Agricultural University of Hebei, 1994, 17(3): 5-10.
[2] 李文庆, 张民, 束怀瑞. 氮素在果树生理上的作用[J]. 山东农业大学学报(自然科学版), 2002, 33(1): 96-100. Li W Q, Zhang M, Shu H R. The physiological effects of nitrogen on fruit trees[J]. Journal of Shandong Agricultural University(Natural Science), 2002, 33(1): 96-100.
[3] 姜远茂, 张宏彦, 张福锁. 北方落叶果树养分资源综合管理理论与实践[M]. 北京: 中国农业大学出版社, 2007. Jiang Y M, Zhang H Y, Zhang F S. Concepts and practice of integrate nutrient resource management in northern deciduous fruit [M]. Beijing: China Agricultural University Press, 2007.
[4] 赵 平, 林克惠, 郑 毅. 氮钾营养对烟叶衰老过程中内源激素与叶绿素含量的影响[J]. 植物营养与肥料学报, 2005,11(3): 379-384. Zhao P, Lin K H, Zheng Y. Effect of N and K nutrition on chlorophyll content and endogenous hormones in the process of tobacco senescence[J]. Plant Nutrition and Fertilizer Science, 2005, 11(3): 379-384.
[5] 张志良. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2001. Zhang Z L. Plant physiology experiment [M].Beijing: Higher Education Press, 2001.
[6] Lu Y H, Watanabe A, Kimura M. Input and distribution of photosynthesized carbon in a flooded soil[J]. Global Biogeochemistry Cycles, 2002, 16: 321-328.
[7] 齐鑫, 王敬国. 应用13C脉冲标记方法研究不同施氮量对冬小麦净光合碳分配及其向地下输入的影响[J]. 农业环境科学学报, 2008, 27(6): 2524-2530. Qi X, Wang J G. Distribution and translocation of assimilated C pulse-labeled with13C for winter wheat (TrticumaestivumsL.), as affected by nitrogen supply[J]. Journal of Agro-Environment Science, 2008, 27(6): 2524-2530.
[8] Takei K, Sakakibara H, Taniguchi M, Sugiyama T. Nitrogen dependent accumulation of cytokinins in root and the translocation to leaf: implicaion of cytokinin species that induces gene expression of maize response regulator[J]. Plant and Cell Physiology, 2001, 42(1): 85-93.
[9] 彭静, 彭福田, 魏绍冲, 等. 氮素形态对平邑甜茶IPT3表达与内源激素含量的影响[J]. 中国农业科学, 2008, 51(11): 3716-3721. Peng J, Peng F T, Wei S C et al. Effect of nitrogen forms onIPT3 expression and hormone contents of Pingyitiancha(MalushupenensisRehd.)[J]. Scientia Agricultura Sinica, 2008, 51(11): 3716-3721.
[10] Shindy W W. Export of photosynthetic effected when leaves are pretreated with growth substances[J]. Nature, 1970, 277: 301-302.
[11] Weaver R J, Shindy W W, Kliewer W M. Growth regulator induced movement of photosynthetic products into fruits of Black Corinth grapes[J]. Plant Physiology, 1969, 44: 183-188.
[12] 彭良志, 胥洱. BA和GA3对华盛顿脐橙幼果14C-光合产物调配的影响. 园艺学报, 1990, 17(2): 111-116. Peng L Z, Xu E. The effects of BA and GA3on14C-assimilate partitioning in young fruits of Washington navel orange(Citrus Sinensis Osbeck)[J]. Acta Horticulturae Sinica, 1990, 17(2): 111-116.
[13] Cairney J W G, Alexander I J. A study of aging of spruce [Piceasitchensis(Bong.) Carr.] ectomycorrhizas. II. Carbohydrate allocation in agingPiceasitchensis/Tylosporafibrillosa(Burt.) Donk ectomycorrhizas[J]. New Phytologist, 1992, 122: 153-158.
[14] Domisch T, Finer L, Lehto T. Effects of soil temperature on biomass and carbohydrate allocation in Scots pine(Pinussylvestris) seedlings at the beginning of the growing season[J]. Tree Physiology, 2001, 21: 465-472.
[15] Walch L P, Ivanov I I, Filleur Setal. Nitrogen regulation of root branching[J]. Annals of Botany, 2006, 97: 875-881.
[16] Caba J M, Centeno M L, Fernandez Betal.Inoculation and nitrate alter phytohormone levels in soybean roots: differences between a supernodulating mutant and the wild type[J]. Planta, 2000, 211: 98-104.
[17] Tian Q Y, Chen F J, Liu J X, et al. Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots[J]. Journal of Plant Physiology, 2008(165): 942-951.
[18] Tian Q Y, Sun P, Zhang W H. Ethylene is involved in nitrate-dependent root growth and branching inArabidopsisthaliana[J]. New Phytologist, 2009(184): 918-931.
[19] Zhao D Y, Tian Q Y, Li L H, Zhang W H. Nitric oxide is involved in nitrate-induced inhibition of root elongation inZeamaysL[J]. Annals of Botany, 2007(100): 497-503.
[20] 郭建军, 叶庆生, 李玲. GA调节禾谷类α-淀粉酶基因表达的信号转导及分子机制[J]. 植物学通报, 2002, 19(1): 63-69. Guo J J, Ye Q S, Li L. Physiological and molecular mechanism of GA regulating α-amylase gene expression in cereal grains[J]. Chinese Bulletin of Botany, 2002, 19(1): 63-69.
[21] 沈德绪, 林伯年. 果树童期与提早结果[M]. 上海: 上海科技出版社, 1989. Shen D X, Lin B N. The juvenescent phase and early fruiting in fruit trees [M]. Shanghai: Shanghai Science and Technology Publishers, 1989.
Annual utilization and allocation of urea-13C byM.hupehensisRehd. under different N rates
LI Jing, JIANG Yuan-mao*, WEI Shao-chong, GE Shun-feng, LI Hong-na, MEN Yong-ge, ZHOU Le
(StateKeyLaboratoryofCropBiology,CollegeofHorticultureScienceandEngineering,ShandongAgriculturalUniversity,Tai’an,Shandong271018,China)
【Objectives】 Unreasonable use of nitrogen often causes imbalance of C/N, excessive vegetative growth, premature senescence of apple trees and decreases the yield and fruit quality as a result. Utilization and allocation characteristics of photosynthates under different nitrogen application levels should be studied to ensure steady growth and fruiting of apple trees. 【Methods】 A pot experiment was conducted with two-year-oldM.hupehensisRehd.(Yan Fu3/M26) seedlings as test materials. Urea was labeled with13C and applied in three levels of N 0, 100 and 200 kg/hm2. The plants were sampled after 72 h of13C labeling and then divided into roots, branches, interstocks, new shoots and leaves, then dried, grinded and weighed. The abundance of13C, the ZR(trans-Zeatin-riboside) and ABA(abscisic acid) contents, the soluble starch, leaf area and chlorophyll contents were determined. 【Results】 Compared in the N0treatment, both the rational application rate of nitrogen fertilizer(N100) and overuse rate(N200) promote the tree biomass, the area and chlorophyll contents of leaves, the promoting effect of N100is gradually appeared with the growth stages. At the spring shoot growing stage(SSG) and spring shoot stop growing stage(SSS), the number of fine root in N100treatment is the highest, followed by N200and N0treatments successively; at the autumn shoot growing stage(ASG), the fine roots of the N0treatment are increased significantly to the highest. The leaf starch content of the N0treatment is higher than those of the N100and N200treatments at all the growing stages. At the early stage of nitrogen application, the ZR contents of leaves are in order of N200>N100>N0, gradually, the ZR content of the N200treatment is decreased significantly. In comparison, no significant differences are found on the ABA contents among the treatments at the early fertilization stage, and the ABA content of the N0treatment is increased and maintains at high level until the late growing stage. Significant differences can be seen in the root to shoot ratio and photosynthate partition among different nitrogen treatments at different growing stages. The root13C partitions in the N100and N200treatments are significantly increased to 285.35% and 217.98%(compared with the N0treatment) at the spring shoot growing stage, more is distributed to the aboveground in the N0treatment. At the stage of autumn shoot growing, the distribution of photosynthate to roots in the N0treatment is improved, while those in the N100and N200treatments are decreased to 71.98% and 41.26% of the N0treatment. 【Conclusions】 The impact on the growth and the photosynthate partition under different nitrogen levels is closely related to the synthesis of ZR and ABA. The nitrogen application significantly improves the partition of photosynthate in roots by large amount syntheses of ZR, and the root to shoot ratio and root biomass are significantly increased at the long time scales. More photosynthates convert into starch under the no nitrogen treatment during the growing stage is caused by ABA synthetic differences.
apple interstock; N fertilization; photosynthate; utilization; distribution
2013-10-08 接受日期: 2014-07-17 网络出版日期: 2015-04-21
现代农业产业技术体系建设专项(CARS-28);公益性行业(农业)科研专项(201103003);山东省农业重大应用创新课题(201009)资助。
李晶(1983—),女,山东济宁人,博士,主要从事果树营养生理研究。E-mail:heavenlyking1010@163.com * 通信作者 Tel: 0538-8249778, E-mail:ymjiang@sdau.edu.cn
S661.1; S606+.2
A
1008-505X(2015)03-0800-07
