施肥及种植密度对大豆根瘤类菌体产生的影响
2015-06-15杨名方张崇玉付责中秦仕华
杨名方+张崇玉+付责中+秦仕华


摘要:通过大田试验,以探明大豆根瘤类菌体生长随播种时间的变化趋势,以及施用有机肥作为基肥、配施钼肥和铁肥与大豆种植密度等因素对大豆根瘤菌产生的影响。结果表明,在播种后40~90 d,大豆根瘤菌粒数、鲜质量和干质量均随播种时间呈先增加后降低的趋势,其最大值均出现在播种后70 d左右,此时大豆刚刚进入结荚期;有机肥作为基肥对根瘤菌的产生影响最大,其后依次是钼肥、种植密度、铁肥;根瘤菌产量最高的组合是不施有机肥与钼肥,铁肥施肥量为150 g/hm2,大豆撒播密度为1.5×107 株/hm2。
关键词:大豆;根瘤菌;播种时间;钼肥;铁肥;种植密度
中图分类号: S565.104;Q945.13 文献标志码: A
文章编号:1002-1302(2015)04-0105-03
收稿日期:2014-10-06
作者简介:杨名方(1987—),女,硕士,从事施肥原理与养分管理研究。E-mail:461810510@qq.com。
通信作者:张崇玉,博士,教授,从事植物营养与环境科学研究。E-mail:zhcy600116@sina.com。
生物固氮是指固氮微生物将空气中游离态的氮素转变为氨态氮,并能被植物吸收利用的过程。目前,人类对豆科植物的共生固氮利用空间还很大[1],如何使非豆科植物特别是主要粮食作物具有固氮作用更是人们正在努力探索的方向[2],原生态根瘤菌产品的工业化具有广阔的市场前景。对根瘤类菌体产生的适宜条件开展研究,可以增强人们对生物固氮的认识,对通过种植原生态大豆为微生物肥料生产提供大量优质的原始根瘤菌群具有重大意义[3]。
在大豆-根瘤共生体系中,根瘤为大豆提供了氮源,同时可从寄主植株吸收养分供自身生长。有研究表明,通过施肥等方式可使寄主植物地上部分营养供应增加、新陈代谢增强,但同时也减少了光合产物向地下部分运输,影响了根瘤的生长[4]。固氮酶能让固氮微生物在常温下将氮气分子转化为氨,并可进一步分化为钼铁蛋白和铁蛋白2个大小不同的深褐色蛋白,且只有在2个蛋白处于结合状态的时候才具有催化活性,钼铁元素对固氮酶具有十分重要的作用。氧气对固氮酶极易产生不可逆转的破坏,既破坏固氮酶的活性,又破坏固氮酶的生物合成,对生物固氮本身非常有害,固氮酶中的铁蛋白对氧尤其灵敏[5]。
在寄主豆科植物幼苗长出2~3片真叶时,根瘤菌首先进入根毛,经过1条“侵入线”进入皮层细胞并立即被质膜包围,在形成的包膜中繁殖、发育为成熟的类菌体。在这个系统中,豆血红蛋白、膜和末端氧化酶系统共同创造了一个既能保护固氮酶的活力,又能有效利用的低浓度氧环境,使生物固氮能够在类菌体中进行[6]。本试验通过施用有机肥作为基肥提供氮、磷、钾等主要营养元素,施用不同质量的钼肥和铁肥,设置不同的大豆种植密度以影响寄主植株的通风透光性等,观察和分析根瘤类菌体的产生情况,并通过正交试验,获得根瘤类菌体产生条件的优化组合。
1 材料与方法
1.1 试验地概况与试验材料
试验在贵州大学农学院松林坡试验地(SLP)大田进行,土壤pH值为7.65,全氮、有机质含量分别为1.97、18.26 g/kg,碱解氮、速效磷、速效钾含量分别为11.2、30.1、407.24 mg/kg;选取贵阳市花溪区农户自留的大豆种为试材,以撒播方式进行种植。大豆基肥为实验室专用有机肥,其养分含量为全氮0.92%、有效磷(P2O5)2.69%、有效钾(K2O)0.98%;钼酸铵试剂由洛阳欧瑞冶金科技有限公司生产,氧化钼含量为83.19%,用以配制钼肥;硫酸亚铁由天津市永大化学试剂有限公司生产,硫酸亚铁含量为99.0%,用以配制铁肥。
1.2 试验设计
1.2.1 根瘤菌收获时间的筛选 对根瘤菌收获时间的筛选采用单因素试验,不施用任何肥料,大豆种植密度为1.5×107 株/hm2。重复3次。
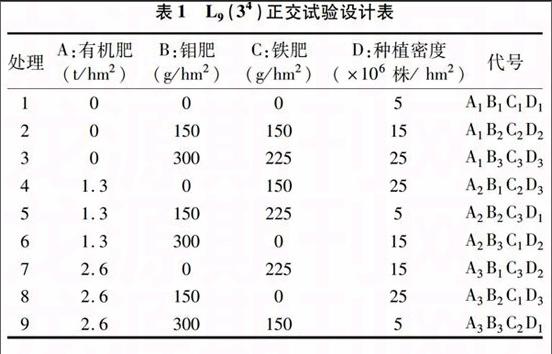
1.2.2 施肥因素与种植密度的筛选 根据大豆根瘤菌生长所需营养元素和大豆生长对种植密度的要求[7],选取有机肥(A)、钼肥(B)、铁肥(C)、种植密度(D)为考察因素,按照 L9(34) 正交表(表1)进行试验,共计9个处理,每处理重复3次。有机肥和钼肥在播种前撒施,铁肥于大豆播种25 d左右喷施于叶面上[8]。
1.3 试验处理
每1小区为1个处理,面积为3 m2(2 m×1.5 m)。大豆于4月20日播种,播后40 d开始采样,至播后90 d止,每隔4~5 d采样1次,采样时,每个小区随机选取大豆10株,根部随同泥土一起迅速带回实验室进行处理,以根瘤菌粒数、鲜质量、干质量作为测试指标,记录数据。正交试验以播后70 d的根瘤菌粒数和干质量各50%权重进行计算来作为根瘤菌的产量。
1.4 数据统计及分析
采用Excel 2007和DPS软件进行统计和数据分析。
表1 L9(34)正交试验设计表
处理 A:有机肥
(t/hm2) B:钼肥
(g/hm2) C:铁肥
(g/hm2) D:种植密度
(×106株/ hm2) 代号
1 0 0 0 5 A1B1C1D1
2 0 150 150 15 A1B2C2D2
3 0 300 225 25 A1B3C3D3
4 1.3 0 150 25 A2B1C2D3
5 1.3 150 225 5 A2B2C3D1
6 1.3 300 0 15 A2B3C1D2
7 2.6 0 225 15 A3B1C3D2
8 2.6 150 0 25 A3B2C1D3
9 2.6 300 150 5 A3B3C2D1
2 结果与分析
2.1 播种时间对大豆根瘤菌粒数和质量的影响
根瘤菌与大豆形成共生关系,其产生的瘤数、质量均会随大豆生理周期的变化而发生变化。由图1可知,随播种天数的增加,根瘤菌的粒数呈现先增加后降低的趋势;在播后40~50 d,根瘤菌粒数呈上升趋势,50~60 d,根瘤菌粒数变化比较平稳;60~70 d粒数再次增加,在70 d时达到最大值;70~90 d,结瘤数逐渐减少。
由图2可知,根瘤菌的质量随播种时间的变化趋势与粒数相似,在播种40~70 d不断增加并达到最大值,70~90 d逐渐下降;鲜质量的变化幅度大于干质量的变化幅度。
在大豆-根瘤共生体系中,大豆与根瘤菌的共生固氮作用所固定的氮素占大豆一生需氮量的50%~60%[9],而根瘤的生长需要依靠寄主植物提供养分才能完成。从大豆的生长周期来看,在播后70 d左右大豆处于结荚期,并从营养生长进入生殖生长阶段,此时,根瘤菌的活性逐渐降低;播后90 d,大豆种子进入成熟期,植株逐渐枯萎,根瘤体系也随之消亡[10]。随着活性的降低,根瘤菌群抵抗外界干扰的能力降低,会受到土壤中其他微生物的侵袭,出现大量坏烂和空壳的现象。因此,在大豆开始结荚以后,根瘤菌的数量和质量均呈现减少和下降的趋势。由此可见,为了获得较高产量的根瘤菌,需在大豆刚刚开始进入生殖生长的时期进行收集。
2.2 正交试验
2.2.1 正交试验各因素重要性分析 由表2、表3、表4可知,最优组合为A1B1C2D2,即不施用有机肥和钼肥,铁肥施肥量为150 g/hm2、大豆种植密度为 1.5×107株/hm2;试验因素主次顺序为:A>B>D>C,即影响因素最大的是有机肥作为基肥的施肥浓度,其次是钼肥、大豆种植密度,影响因素最小的是施用铁肥浓度。
表2 大豆施肥因素与种植密度等正交试验结果
小区代号
根瘤菌粒数(粒) 根瘤菌干质量(g)
重复1 重复2 重复3 重复1 重复2 重复3
A1B1C1D1 157 153 128 0.878 3 0.965 5 0.827 9
A1B2C2D2 145 163 149 0.773 9 1.321 5 0.989 2
A1B3C3D3 79 92 71 0.483 0 0.567 3 0.526 4
A2B1C2D3 81 80 72 0.470 3 0.576 6 0.331 7
A2B2C3D1 70 110 82 0.560 6 0.698 0 0.537 7
A2B3C1D2 46 42 62 0.280 2 0.294 7 0.475 1
A3B1C3D2 83 66 57 0.524 6 0.517 3 0.500 7
A3B2C1D3 27 51 34 0.294 7 0.361 4 0.337 9
A3B3C2D1 37 55 52 0.174 5 0.383 6 0.324 1
表3 不同施肥及种植密度下根瘤菌的产量与极差分析
试验小区 有机肥
(t/hm2) 钼肥
(g/hm2) 铁肥
(g/hm2) 种植密度
(株/hm2) 干质量
(g) 粒数
(粒) 综合
评分
A1B1C1D1 1 1 1 1 0.891 146 73.445
A1B2C2D2 1 2 2 2 1.028 152 76.581
A1B3C3D3 1 3 3 3 0.526 81 40.596
A2B1C2D3 2 1 2 3 0.460 78 39.063
A2B2C3D1 2 2 3 1 0.599 87 43.966
A2B3C1D2 2 3 1 2 0.350 50 25.175
A3B1C3D2 3 1 3 2 0.514 69 34.590
A3B2C1D3 3 2 1 3 0.331 48 24.166
A3B3C2D1 3 3 2 1 0.294 37 18.647
k1 63.541 49.033 40.929 45.353
k2 36.068 48.238 44.764 45.449
k3 25.801 28.139 39.717 34.608
R 37.740 20.894 5.047 10.841
注:统计分析使用LSD分析法。
表4 正交设计试验大豆根瘤菌产量方差分析
方差来源 偏差平方和 自由度 F值 P值
A:有机肥 2 284.441 2 2.687 >0.05
B:钼肥 841.106 2 0.989 >0.05
C:铁肥 41.640 2 0.049 >0.05
D:种植密度 232.963 2 0.274 >0.05
试验结果表明,不施用有机肥比施用1.3、2.6 t/hm2有机肥的根瘤菌产量分别高出76.2%和150.9%;施用 150 g/hm2 钼肥的大豆根瘤菌产量比不施用钼肥增加2.1%,比施用300 g/hm2钼肥产量增加32.9%;大豆种植密度为 15×107株/hm2 时根瘤菌产量比种植密度为5×106株/hm2时增产2.2%,比2.5×107株/hm2增产31.3%。铁肥在大豆播后25 d左右喷施时遭遇雨水天气,对正交试验结果存在较大的误差,没有对其进行分析。
3 结论与讨论
试验结果表明,大豆收获根瘤的最佳时间是播种后70 d左右,此时,大豆根瘤菌的粒数和质量均达到最大值。在实际生产中,由于大豆品种、生长特性、地理环境、气候等因素的不同而导致播种时间会有所不同,可以通过形态观察,在大豆植株刚开始进入结荚期、进入生殖生长时作为根瘤菌的最佳收获期。大豆仅靠自身的固氮作用不能满足丰产要求[11-12],需要通过施用有机肥为大豆生长补充氮、磷、钾等主要营养元素。
氮元素对豆科植物生物固氮的共生体系产生重大影响[13-21]。在供试土壤条件下,为获得较高产量的大豆根瘤菌,种植大豆过程不需要施用肥料,土壤中的营养物质可通过大豆吸收来满足根瘤菌的生长。施用有机肥可给大豆植株直接提供丰富的氮源,但使大豆对根瘤中固氮酶提供的氮源依赖性减小,根瘤与大豆的共生关系遭到破坏[10]。植物的磷吸收与根系形状、结瘤数量、籽粒产量等均有密切关系[22-23];钼、铁是生物固氮体系的重要元素,对根瘤生长具有重大的意义[5,8,24]。只有在根瘤菌群中固氮菌数量和活性均处于较好状态时,营养元素才能为根瘤的产量作出较大贡献。
试验结果表明,适宜大豆结瘤的最佳种植密度是 1.5×107株/hm2,约是通常大豆种植密度的1.5倍,这可能与根瘤菌群的需氧性有关。根瘤菌结瘤需要相对密封的状态,通过适当密植,可以减少植株间的通风透光性,为根瘤菌的产生提供适当封闭的空间。同时,不施肥与适当密植,都适当减少了地上部分光合作用和呼吸作用的强度,使根瘤菌能够从寄主植物获得更多的营养物质供给菌群的生长,而种植密度过大,影响了大豆植物的正常生长,不利于根瘤的产生。通过种植大豆收获根瘤的原始菌群,让大豆接近“野生状态”,利于根瘤菌类菌体的产生,可满足工业化生产的较大需求,且操作简便。
在本试验中,铁肥在喷施后受雨水天气影响,结果存在较大误差,有必要对铁肥因素进行进一步验证。同时,本研究只从根瘤菌的产量等相关方面进行了分析,还需要从根瘤菌的菌群种类和固氮菌所占比例及活性等进行分析比较,为筛选适宜根瘤菌生长的环境条件提供更全面的科学依据。
参考文献:
[1]贺学礼,赵丽莉. 陕西豆科固氮牧草的生态学研究[J]. 中国草地,1997(2):45-48.
[2]黄群策,陈启锋,李志真.生物固氮研究的前景[J]. 科技导报,1999(1):23-25.
[3]吴建峰,林先贵. 我国微生物肥料研究现状及发展趋势[J]. 土壤,2002,34(2):68-72.
[4]宋海星,王 萍,申斯乐,等. 大豆共生固氮与叶片全氮含量之间关系的研究[J]. 吉林农业科学,2000,25(6):9-11.
[5]刘 铮. 微量元素的农业化学[M]. 北京:科学出版社,1991:142-170.
[6]洪国藩.生物固氮[J]. 科学,1985,37(1):25-30.
[7]张立军,陈艳秋,孙旭刚,等. 营养元素对大豆氮素营养及产量的影响浅析[J]. 大豆科技,2013(4):59-63.
[8]王永锋,裴桂英,马赛飞,等. 铁肥在大豆上的施用效果 Ⅰ.大豆基施铁肥肥效研究[J]. 耕作与栽培,2006(2):33-36.
[9]Ohwaki Y,Sugahara P. Active extrusion of protons and exudation of carboxylic acids in response to iron deficiency by roots of chickpea(Cicer arietinum L.)[J]. Plant and Soil,1997,189(1):49-55.
[10]陈华癸,李阜棣,陈文新,等. 土壤微生物学[M]. 上海:上海科学技术出版社,1981.
[11]Demon C J,Sutherland P L. Soil-fertility requirement of soybeans with reference to irrigation[M]//Hurdy W H. Irrigated soybean production in arid and semi arid regions. University of Illinois,1979:276-352.
[12]Marscher H. Mineral nutrition of higher plant[M]. London:Academic Press,1986:674.
[13]Kennedy I R. Primary products of symbiotic nitrogen fixation. Ⅰ. Short-term exposures of serradella nodules to N2[J]. Biochem Biophys Acta,1966,130(2):285-294.
[14]Sinclair T R,Serraj R. Legume nitrogen fixation and drought[J]. Nature,1995,378(23):344-347.
[15]王树起,韩晓增,乔云发,等. 施氮对大豆根瘤生长和结瘤固氮的影响[J]. 华北农学报,2009,24(2):176-179.
[16]严 君,韩晓增,王守宇,等. 不同施氮量及供氮方式对大豆根瘤生长及固氮的影响[J]. 江苏农业学报,2010,26(1):75-79.
[17]房增国,左元梅,李 隆,等. 玉米-花生混作体系中不同施氮水平对花生铁营养及固氮的影响[J]. 植物营养与肥料学报,2004,10(4):386-390.
[18]Watababe T,Tabuchi K,Nakano H. Response of soybean to supplemental nitrogen after flowering[M]//Shanmugasundaram S,Sulzberger E W,Clean B T M. Soybean in tropical and subtropical cropping system. Shanhua,Taiwan,China:AVRDC,1986:308-310.
[19]甘银波,本佳婉.不同氮肥管理对毛豆共生固氮及产量的影响[J]. 中国油料,1996(1):34-37.
[20]Streeter J,Wong P P. Inhibition of legume nodule formation and N2 fixation by nitrate[J]. Critical Reviews in Plant Sciences,1988,7(1):1-23.
[21]Dakora D F. A functional relationship between leghaemoglobin and nitrogenase based on novel measurements of the two proteins in legume root nodules[J]. Annals of Botany,1995,75(1):49-54.
[22]Bhuiyan M M H,Rahman M M,Afroze F,et al. Effect of phosphorus,molybdenum and rhizobium inoculation on growth and nodulation of mungbean[J]. Journal of Soil Nature,2008,2(2):25-30.
[23]Murat E B,Yildirim,Enact T,et al. Effects of phosphorus application and rhizobium inoculation on the yield,nodulation and nutrient uptake in field pea(Pisum sativum sp. arvense L.)[J]. Journal of Animal and Veterinary Advances,2009,8(2):301-304.
[24]Kaiser B N,Moreau S,Castelli J,et al. The soybean NRAMP homologue,GmDMT1,is a symbiotic divalent metal transporter capable of ferrous iron transport[J]. The Plant Journal,2003,35(3):295-304.
