荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究
2015-06-01孙小丽康萨如拉张庆常昌明马文静牛建明
孙小丽,康萨如拉,张庆,2,常昌明,马文静,牛建明,2*
(1.内蒙古大学生命科学学院,内蒙古 呼和浩特 010021;2.中美生态、能源及可持续性科学研究中心,内蒙古 呼和浩特 010021)
荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究
孙小丽1,康萨如拉1,张庆1,2,常昌明1,马文静1,牛建明1,2*
(1.内蒙古大学生命科学学院,内蒙古 呼和浩特 010021;2.中美生态、能源及可持续性科学研究中心,内蒙古 呼和浩特 010021)
物种多样性对生态系统功能的影响是生态学领域的核心问题之一。本研究利用内蒙古荒漠草原80个野外植被、土壤调查样地资料,结合气候数据,探讨物种多样性、生产力与气候因子和土壤养分的关系,为进一步认识多样性-生产力关系提供参考。结果表明,1)物种多样性、生物量与年平均降水量均呈线性正相关,与年平均温度均呈线性负相关,与干燥指数均呈线性正相关。2)物种丰富度与土壤全磷含量呈线性负相关(R2=0.072,P<0.05),与速效氮含量呈线性正相关(R2=0.183,P<0.01),Shannon-Wiener指数与速效氮也呈线性正相关(R2=0.061,P<0.05);生物量与土壤全氮、速效氮和有机碳含量都呈线性正相关,相关系数分别为0.150,0.177,0.089(P<0.01)。3)群落生物量与物种多样性呈线性正相关关系。4)干燥指数是影响多样性和生物量的主要环境因子。
荒漠草原;生产力;物种多样性;土壤养分;水热因子
物种多样性对生态系统功能的影响是生态学领域的核心科学问题之一[1-2]。生产力水平的高低是生态系统功能的重要表现形式,所以探讨物种多样性与生产力之间的关系对于研究物种多样性对生态系统功能的影响具有重要意义[3-4]。
物种多样性不仅能够度量群落的组成结构和功能的复杂性,而且也能指示环境[5]。近年来,围绕生物多样性、生产力与环境因子的关系,国内外开展了一系列讨论[6-9],其中较多的是关于物种多样性、生产力与纬度和海拔导致的光照和水热因子变化的关系[6-7]。尤其是在干旱、半干旱地区,多样性和生产力极大地受到降水条件的影响[10]。植物的生长发育与土壤因子的关系也十分密切,土壤理化性质的不同以及土壤母质条件等,都可能影响群落结构、物种多样性和生产力水平[11-12]。关于土壤养分与物种多样性的关系已有一些报道,然而由于土壤养分对多样性的影响复杂多样,研究结果缺乏一致的规律性。目前关于多样性、生产力与土壤养分关系的研究多涉及森林[13-14]、草原[15-16]、高寒草甸[17-18]等,但在本研究区,荒漠草原的理论探讨还不够全面。
内蒙古荒漠草原是草原向荒漠过渡的旱生性最强的草原生态系统[19]。前人在该地区积累了大量的研究成果及文献资料,如张庆等[20]对群落分布格局与环境因子关系的探讨,杨雪栋等[21]对小针茅(Stipaklemenzii)草原土壤养分化学计量学特征的研究,马文静等[22]阐述了短花针茅(Stipabreviflora)草原物种多样性、功能多样性与生产力之间的关系。但是该地区尚缺乏联系气候因子和土壤养分等环境因子的多样性、生产力研究。本研究通过对内蒙古荒漠草原进行植被与土壤调查,结合气候资料,在群落水平上探讨内蒙古荒漠草原物种多样性、生产力及其与气候因子和土壤养分的关系,为进一步认识物种多样性-生产力关系提供参考。
1 材料与方法
1.1研究区概况
研究区位于内蒙古中温型荒漠草原区,地处107°18′-115°35′ E,41°11′-45°5′ N,海拔905~1725 m。该地区年平均降水量为150~250 mm,多集中在夏季;年平均温度2~5℃,≥10℃年积温2200~2500℃。地带性土壤为棕钙土[19,23],主要植被类型为小针茅荒漠草原和短花针茅荒漠草原。
图1 样点分布图
1.2数据获取
植被调查选择在群落生物量高峰期进行,于2012年7月下旬至8月中旬完成。共设置了80个具有代表性的样地(图1)。植被调查采用的是样方法,每个样地在10 m×10 m的范围内,随机设置3个1 m×1 m测产样方,记录样方中出现的每一物种名、株丛数,并按物种将植株地上部分齐地面刈割,称取鲜重,然后分别用报纸包好保存,带回实验室置于65℃烘箱烘干至恒重,称取干重。另做7个频度样方,记录样方中出现的物种名。同时,用土钻钻取0~30 cm混合土样,3次重复。带回实验室风干后过1 mm和0.15 mm筛后用于测定土壤基本指标。测试项目为土壤的全氮(total nitrogen,TN)(硒粉、硫酸铜、硫酸钾消化-蒸馏法[24])、全磷(total phosphorus,TP)(碱熔-钼锑抗比色法[24])、速效氮(available nitrogen,AN)(凯氏定氮法[24])、速效磷(available phosphorus,AP)(碳酸氢钠浸提-钼锑抗比色法[24])和有机碳(organic carbon,OC)(重铬酸钾氧化外加热法[24])。群落生产力(productivity)用3个测产样方地上生物量干重的平均值代替。群落的物种丰富度由10个样方中出现的物种数决定。
年平均降水量(mean annual precipitation, MAP)和年平均温度(mean annual temperature, MAT)数据采用WORLDCLIM网站(http://www.worldclim.org/)下载的当前(1950-2000年)监测数据,数据空间分辨率为arc 30 s。干燥指数(aridity index, AI)数据源于CGIAR-CSI GeoPortal网站(http://www.csi.cgiar.org)的数据产品,这一产品是根据WORLDCLIM网站的数据建模生成的[25]:
AI=MAP/MAE
式中,AI为干燥指数,MAP为年平均降水量,MAE(mean annual potential evapo-transpiration)为年平均潜在蒸散量。AI值越大代表气候条件越湿润,AI值越小代表气候条件越干旱。
1.3植物群落物种多样性的指标
物种丰富度指数:R=S
式中,R为群落物种丰富度指数,S为群落中的总物种数。
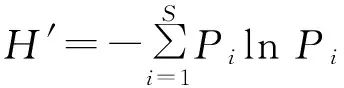
式中,H′为Shannon-Wiener指数,Pi为种i的相对重要值=(相对生物量+相对多度+相对频度)/3,S为种i所在样地所有物种数。
1.4数据处理
采用线性回归方法分析物种多样性、生物量与气候因子和土壤养分之间的关系以及物种多样性与生物量之间的关系。通过多元逐步回归分析确定气候因子及土壤因子对群落丰富度、多样性和生物量变异的贡献。将引入变量的P值定为0.05,剔除变量的P值定为0.10,判定系数(R2)揭示了回归模型中自变量能够解释的因变量变异比例。
使用SPSS 21.0软件进行统计分析,采用SigmaPlot 12.5绘散点图制。
2 结果与分析
2.1物种多样性、生物量与气候因子的关系
根据研究区80个样地的物种丰富度、Shannon-Wiener指数和生物量与气候因子建立的回归模型(图2)可以看出,物种丰富度、Shannon-Wiener指数和生物量均随着年平均降水量的增加而显著上升(图2A,B,C),其中物种丰富度和生物量与年平均降水量呈极显著的线性正相关;Shannon-Wiener指数和生物量与年平均温度均呈显著的线性负相关(图2E,F),物种丰富度与年平均温度相关性不显著(图2D);物种丰富度、Shannon-Wiener指数和生物量均随着干燥指数的增大呈显著的上升趋势(图2G,H,I)。

图2 物种丰富度、多样性和生物量与水热因子和干燥指数的关系
2.2物种多样性、生物量与土壤养分的关系
由表1可知,物种丰富度与土壤全磷含量呈线性负相关(P<0.05),物种丰富度、Shannon-Wiener指数与土壤速效氮含量具有显著的线性正相关关系(P<0.01、P<0.05),生物量与土壤全氮、速效氮以及有机碳含量均呈极显著的正相关(P<0.01)。其他土壤养分与物种丰富度、Shannon-Wiener指数和生物量关系不显著。

表1 物种丰富度、多样性和生物量与土壤养分含量的关系Table 1 The regression analyses of species richness, species diversity, and biomass with soil nutrient content
NS 代表关系不显著;PL代表正线性关系;NL 代表负线性关系。
NS indicates non-significant relationship; PL indicates positive linear relationship; NL indicates negative linear relationship.
2.3物种多样性与生物量的关系
群落生物量与物种丰富度和Shannon-Wiener指数呈显著的线性正相关,相关性系数分别为0.076(P<0.05)和0.074(P<0.05),即物种多样性对生产力存在正效应(图3)。

图3 物种丰富度、多样性与生物量的关系
2.4影响物种多样性、生物量的主导环境因子
通过多元逐步回归分析环境因子对物种丰富度的影响发现,该模型引入了AI、TP和AN三个变量,剔除了其他变量,且这3个因子与物种丰富度均存在显著相关关系,解释了物种丰富度的30.9%,其中AI作为第一解释变量进入方程,解释了总变异的21.8%,成为影响群落物种丰富度最主要的环境因子(表2)。另外,MAT作为最重要的因子被引入解释Shannon-Wiener指数的方程中,解释率为9.9%,TP和AN同样作为第二和第三因子被引入,说明MAT是对Shannon-Wiener指数影响最大的环境因子。环境因子对生物量影响的多元逐步回归分析,AI作为唯一因子被引入方程,解释了群落生产力的27.2%。
2.5干燥指数对多样性-生产力关系的影响
为进一步探讨干燥指数对物种多样性-生产力关系的影响,按联合国环境规划署基于干燥指数的全球气候分类标准[26]将研究区划分为干旱、半干旱两种干湿气候类型。研究区80个样地,其中25个分布在干旱区,55个分布在半干旱区。利用线性回归方法分析两种不同气候类型(干旱区、半干旱区)下多样性-生产力的关系,依据R2分析干燥指数对两者关系的影响(图4)。发现无论物种丰富度还是Shannon-Wiener指数,当干燥指数高时(半干旱区)对生产力的贡献,均比干燥指数低时(干旱区)大。
3 结论与讨论
3.1水热因子对物种多样性、生产力的重要性

表2 多元逐步回归分析结果Table 2 Results of stepwise regression analysis
AI:干燥指数 Aridity Index; TP:全磷 Total phosphorus;AN:速效氮 Available nitrogen;MAT:年平均温度 Mean annual temperature.
内蒙古草原处于干旱半干旱区,其生物多样性和生产力深受降水条件的限制,大量研究表明,随着降水量的增加,生物多样性和生产力呈显著增加的趋势[27-28]。有学者对美国[29]、委内瑞拉[30]、阿根廷[31]干旱区草原生态系统的研究均证明物种丰富度与降水量呈显著的正相关关系。本研究中物种多样性和生产力与年平均降水量呈显著正相关,与年平均温度呈负相关,与干燥指数呈显著正相关。物种丰富度、Shannon-Wiener指数和生产力与水热因子相关性的比较结果显示,水热因子与群落生产力的相关性最高,可以解释其变异的13.0%~28.1%,降水与物种丰富度的相关性高于Shannon-Wiener指数,可以解释其变异的22.1%,3个气候要素中, 作为水热综合因子的干燥指数同物种丰富度和初级生产力的相关性最高。白永飞等[8]对锡林河流域植物群落的研究也表明物种丰富度、多样性和生产力与年降水量正相关,与年平均气温负相关,而水热因子可以解释生产力的变异高达71.31%~77.58%,可以解释物种丰富度变异的40.2%~61.39%;Kang等[28]对整个内蒙古草原的研究发现地上生物量与年平均降水量的相关系数高达65%;然而马文静等[22]对短花针茅荒漠草原的研究显示年平均降水量对物种丰富度和生产力的解释率也不高,分别为12.6%和10.5%。造成荒漠草原这种解释率偏低的原因可能与其独特的生境特征有关,该区年平均降水量普遍偏低,导致研究区内降水梯度不明显;此外土壤、放牧等因素也可能起重要作用。在本研究区内,存在着随年平均温度升高,年平均降水量显著降低的特点,二者呈极显著的负相关,因此,物种多样性和生产力与年平均温度呈负相关,极有可能是由于水分的限制,导致植物无法利用这些能量进行光合作用,从而呈随着能量增加群落物种多样性和生产力降低的关系。
3.2土壤养分对物种多样性、生产力的重要性
在土壤-植被体系中,土壤和植被是两个相互依存的因子,植被影响土壤,土壤制约植被[32]。探讨土壤养分与物种多样性的关系,对了解生态系统的功能与过程具有重要意义。目前二者之间关系的研究结果缺乏一致的规律性。多数氮素添加实验研究发现,增加氮素会导致物种多样性下降[33-34],Perroni-Ventura等[12]对热带半干旱生态系统的研究显示物种丰富度随着土壤氮含量的增加而升高。Olsen等[35]则认为在贫瘠的土壤里增加氮含量有利于提高物种丰富度,在养分丰富的土壤增加氮含量会使物种丰富度降低。究其原因,对退化的、资源贫瘠的群落添加氮素可以减轻氮素的限制,从而提高物种丰富度;相反,对相对成熟稳定的群落添加氮素可能会破坏原有生态系统的共存机制,导致物种丰富度下降[36]。就磷元素而言,张林静等[37]认为土壤全磷含量与物种多样性没有显著相关性,Jouany等[38]对欧洲温性草原和Perroni等[39]对墨西哥的草原-荒漠疏林生态系统研究均表明物种丰富度随着土壤中全磷含量的升高而降低。本研究发现在荒漠草原这一生态系统类型中,物种多样性沿着速效氮含量增加的梯度呈升高的趋势,沿着土壤全磷含量增加的梯度呈降低的趋势。地上生物量与有机碳、全氮和速效氮含量呈显著的线性正相关,这与左小安等[5]对科尔沁沙地的研究是一致的。土壤中的养分,在空间和时间上的分布是具有异质性的,作为对土壤养分异质性的反应,植物在养分丰富的局部环境中能选择性地改变其根系的生长,从而增加养分的吸收[40]。土壤中养分含量的高低直接影响群落的生产力,土壤养分越丰富,群落生产力越高[41]。由表1可知,速效氮与物种多样性和生产力的相关系数均最高,这可能是因为草地生态系统普遍缺氮,物种多样性和生产力对氮含量的增加反应比较敏感,这在一定程度上支持了Vitousek等[42]的观点,即氮素是草地生态系统净初级生产力的限制因子,同时也是衡量草地生产力的重要指标。Tilman等[43]认为高的植物多样性能够更完全地利用土壤中的限制性养分,减少生态系统中N素的淋溶损失。同时,多样性高的生态系统能够保持更高比例的限制性养分,使土壤养分的总储量增加,从而使生产力增加。有学者指出氮素对草地生产力的有效性受到水分的限制,尤其在干旱区,水分的缺乏会导致氮素的有效性偏低[44]。因此,降水的缺乏极有可能是导致荒漠草原氮素对生产力贡献(R2=0.177)低于典型草原和草甸草原(R2=0.65)[8]的主要原因。
3.3水热因子是影响物种多样性和生产力的主要环境因子
为进一步确定影响荒漠草原物种多样性和生产力的主导环境因子,建立了多元逐步回归模型,发现干燥指数作为解释物种丰富度的第一因子和解释群落生产力的唯一最重要的因子被引入,而且都呈显著的相关关系,年平均温度作为解释Shannon-Wiener指数的第一因子被引入模型。干燥指数是水分与温度综合作用的体现,这充分说明了水热因子在内蒙古荒漠草原区对物种多样性和生产力作用的主导地位。白永飞等[8]发现作为水热综合因子的干燥度与锡林河流域植物群落多样性和生产力的相关系数是最高的;郑晓翾等[45]对呼伦贝尔草甸草原、典型草原和荒漠草原3种草原类型的研究也证明水热因子是影响物种丰富度的主要环境因子。但是同时也发现,这种线性关系的相关系数并不高,这可能与研究尺度和放牧利用等因素有关。在大的空间尺度下,气候因子的梯度变化更为突出,稀释了局地尺度人类活动的作用,因此气候对物种多样性分布格局解释量相对较高,在小的空间尺度下,气候因子的梯度变化不明显,气候解释量反而下降[46-47]。此外,放牧是荒漠草原区的主要利用方式,牲畜对牧草的采食和土壤的践踏以及牲畜体内营养物质的归还和转化,对草地的微环境和土壤条件会造成显著影响[48],进而影响植物群落物种组成[49]、丰富度[50]、多样性[51]等。本研究区内群落结构较为简单,更易受到人类活动的干扰。目前,人类活动很可能是影响内蒙古荒漠草原多样性和生产力格局的一个重要因素。
3.4物种多样性与生产力之间的关系受到环境因子的制约
许多研究证实物种多样性与生产力主要存在4种关系:正相关[43];负相关[52];单峰曲线[53];不相关[54]。其中呈单峰曲线和正相关关系的结论居多。就荒漠草原而言,物种多样性与生产力之间呈线性正相关,这一结论与马文静等[22]对内蒙古短花针茅荒漠草原的研究相一致。在不同尺度下,物种多样性和生产力的关系存在差异。在单一的群落类型内,多样性与生产力之间的关系常常并不显著[55],而当研究区包括几个群落类型的时候,二者之间的关系常常表现出单峰曲线的形式[56]。Ni等[57]在不同尺度上对内蒙古不同类型草原物种多样性与生产力关系的研究结果也发现在局地尺度上,物种丰富度与生物量呈线性正相关,随着尺度的增大,物种丰富度与生产力的关系可能会出现单峰曲线关系。
干燥指数、年平均温度、全磷和速效氮进入了解释物种丰富度、Shannon-wiener指数的方程中,表明水热因子和土壤养分对物种多样性有重要影响,进而推测,荒漠草原多样性-生产力关系同时受到上述主要气候因子与土壤养分的调控。探讨干燥指数对物种多样性-生产力关系的影响,发现无论物种丰富度还是Shannon-Wiener指数,当干燥指数高时(半干旱区)对生产力的贡献,均比干燥指数低时(干旱区)大(图4)。He等[58]研究了在土壤养分N、P、K含量存在差异的情况下, 物种丰富度和生产力之间的关系,结果表明,在土壤N、P、K含量高的群落中,物种丰富度和生产力呈显著的正相关关系,而在土壤养分N、P、K含量低的处理中, 两者之间没有明显的关系。出现上述现象的原因可能是:植物在资源利用上具有互补性,不同的植物具有不同的资源利用形式,资源供给水平高可以使得物种间对资源的利用差异充分体现出来,多样性的作用表现更加充分,这样就加强了多样性与生产力之间正相关关系的表现。
[1] Loreau M, Naeem S, Inchausti P,etal.Biodiversity and ecosystem functioning:Current knowledge and future challenges.Science, 2001, 294:804-808.
[2] Hooper D U, Chapin F S III, Ewel J J,etal.Effects of biodiversity on ecosystem functioning:A consensus of current knowledge.Ecological Monographs, 2005, 75(1):3-35.
[3] Naeem S.Ecosystem consequences of biodiversity loss:The evolution of a paradigm.Ecology, 2002, 83(6):1537-1552.
[4] Tilman D, Reich P B, Knops J,etal.Diversity and productivity in a long-term grassland experiment.Science, 2001, 294:843-845.
[5] Zuo X A, Zhao X Y, Zhao H L,etal.Changes of species diversity and productivity in relation to soil properties in sandy grassland in Horqin sand land.Environmental Science, 2007, 28(5):945-951.
[6] Wilson J B, Lee W G, Mark A F.Species diversity in relation to ultramafic substrate and to altitude in southwestern New Zealand.Vegetatio, 1990, 86:15-20.
[7] Zhang Q, Hou X Y, Li F Y H,etal.Alpha, beta and gamma diversity differ in response to precipitation in the Inner Mongolia Grassland.PLoS ONE, 9(3):e93518.doi:10.1371/journal.pone.0093518.
[8] Bai Y F, Li L H, Wang Q B,etal.Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River Basin, Inner Mongolia.Acta Phytoecologica Sinica, 2000, 24(6):667-673.
[9] Zhang Z Z, Wu G L, Wang D,etal.Plant community structure and soil moisture in the semi-arid natural grassland of the Loess Plateau.Acta Prataculturae Sinica, 2014, 23(6):313-319.
[10] Abramsky Z, Rosenzweig M L.Tilman’s predicted productivity-diversity relationship shown by desert rodents.Nature, 1984, 309:150-151.
[11] Long W X, Yang X B, Li D H.Patterns of species diversity and soil nutrients along a chronosequence of vegetation recovery in Hainan Island, South China.Ecological Research, 2012, 27(3):561-568.
[12] Perroni-Ventura Y, Montana C, Garcia-Oliva F.Relationship between soil nutrient availability and plant species richness in a tropical semi-arid environment.Journal of Vegetation Science, 2006, 17(6):719-728.
[13] An S Q, Wang Z F, Zhu X L,etal.Effect of soil factors on species diversity in secondary forest communities.Journal of Wuhan Botanical Research, 1997, 15(2):143-150.
[14] Firn J, Erskine P D, Lamb D.Woody species diversity influences productivity and soil nutrient availability in tropical plantations.Oecologia, 2007, 154(3):521-533.
[15] Kahmen A, Perner J, Audorff V,etal.Effects of plant diversity, community composition and environmental parameters on productivity in montane european grasslands.Oecologia, 2005, 142(4):606-615.
[16] Ceulemans T, Stevens C J, Duchateau L,etal.Soil phosphorus constrains biodiversity across european grasslands.Global Change Biology, 2014, 20(12):3814-3822.
[17] Liu M X, Wang G.Responses of plant community diversity and soil factors to slope in alpine meadow.Chinese Journal of Ecology, 2013, 32(2):259-265.
[18] Wang C T, Long R J, Cao G M,etal.The relationship between soil nutrients and diversity-productivity of different type grasslands in alpine meadow.Chinese Journal of Soil Science, 2008, 39(1):1-8.
[19] Inner Mongolia-Ningxia jiont inspection group of Chinese Academy of Sciences.Vegetation of Inner Mongolia[M].Beijing:Science Press, 1985.
[20] Zhang Q, Niu J M, Buyantuyev A,etal.Ecological analysis and classification ofStipabrevifloracommunities in the Inner Mongolia region:the role of environmental factors.Acta Prataculturae Sinica, 2012, 21(1):83-92.
[21] Yang X D, Sun W G, Baoyin T.Characteristics of soil stoichiometry ofStipaklemeziicommunity in desert steppe of Inner Mongolia.Chinese Journal of Grassland, 2012, 34(5):30-34.
[22] Ma W J, Zhang Q, Niu J M,etal.Relationship of ecosystem primary productivity to species diversity and functional group diversity:evidence fromStipabrevifloragrassland in Nei Mongol.Chinese Journal of Plant Ecology, 2013, 37(7):620-630.
[23] Wei Z J, Han G D, Zhao G,etal.Research on Desert Grassland Ecosystem in China[M].Beijing:Science Press, 2013.
[24] Zhang G L, Gong Z T.Soil Survey Laboratory Methods[M].Beijing:Science Press, 2012.
[25] Trabucco A, Zomer R J, Bossio D A,etal.Climate change mitigation through afforestation/reforestation:A global analysis of hydrologic impacts with four case studies.Agriculture, Ecosystems and Environment, 2008, 126:81-97.
[26] Middleton N J, Thomas D.World Atlas of Desertification[M].London:Arnold, United Nations Environment Programme, 1997.
[27] Han B, Fan J W, Zhong H P.Grassland and biomass of communities along gradients of the Inner Mongolia grassland transect.Journal of Plant Ecology, 2006, 30(4):553-562.
[28] Kang M, Dai C, Ji W,etal.Biomass and its allocation in relation to temperature, precipitation, and soil nutrients in Inner Mongolia Grasslands, China.PLoS ONE, 8(7):e69561.Doi:10.1371/journal.pone.0069561.
[29] Daubenmire R.Steppe vegetation of Washington.Washington Agricultural Experiment Station Technical Bulletin, 1970, 62:1-131.
[30] Sarmiento G.Patterns of specific and phenological diversity in the grass community of the venezuelan tropical savannas.Journal of Biogeography, 1983, 10:373-391.
[31] Jobbagy E G, Paruelo J M, Leon R J C.Vegetation heterogeneity and diversity in flat and mountain landscapes ofPatagonia(Argentina).Journal of Vegetation Science, 1996, 7(4):599-608.
[32] Tilman D.Causes, consequences and ethics of biodiversity.Nature, 2000, 405:208-211.
[33] Zhang J Q, Li Q, Ren Z W,etal.Effect of nitrogen addition on species richness and relationship between species and aboveground productivity of alpine meadow of Qinghai-Tibetan Plateau, China.Chinese Journal of Plant Ecology, 2010, 34(10):1125-1131.
[34] Cornwell W K, Grubb P J.Regional and local patterns in plant species richness with respect to resource availability.Oikos, 2003, 100(3):417-428.
[35] Olsen S L, Sandvik S M, Totland O.Influence of two N-fixing legumes on plant community properties and soil nutrient levels in an Alpine.Arctic Antarctic and Alpine Research, 2013, 45(3):363-371.
[36] Bai Y F, Wu J G, Clark C M,etal.Trade offs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning:Evidence from Inner Mongolia Grasslands.Global Change Biology, 2010, 16(1):358-372.
[37] Zhang L J, Yue M, Gu F X,etal.Coupling relationship between plant communities’ species diversity and soil factors in ecotone between desert and oasis in Fukang, Xinjiang.Chinese Journal of Applied Ecology, 2002, 13(6):658-662.
[38] Jouany C, Cruz P, Daufresne T,etal.Biological phosphorus cycling in grasslands:Interactions with nitrogen.In:Bunemann E K, Oberson A, Frossard E.Phosphorus in Action:Biological Processes in Soil Phosphorus Cycling[M].Berlin:Springer, 2011, 275-294.
[39] Perroni Y, Garcia-Oliva F, Souza V.Plant species identity and soil P forms in an oligotrophic grassland-desert scrub system.Journal of Arid Environments, 2014, 108:29-37.
[40] Grime J P.Biodiversity and ecosystem function:The debate deepens.Science, 1997, 277:1260-1261.
[41] Tilman D.Secondary succession and the pattern of plant dominance along experiment nitrogen gradients.Ecological Monographs, 1987, 57:189-214.
[42] Vitousek P M, Aber J D, Howarth R W,etal.Human alteration of the global nitrogen cycle:Sources and consequences.Ecological Applications, 1997, 7(3):737-750.
[43] Tilman D, Knops J, Wedin D,etal.The influence of functional diversity and composition on ecosystem processes.Science, 1997, 277:1300-1302.
[44] Lee M, Manning P, Rist J,etal.A global comparison of grassland biomass responses to CO2and nitrogen enrichment.Philosophical Transactions of the Royal Society B-Biological Sciences, 2010, 365:2047-2056.
[45] Zhen X X, Jin T T, Mu L F,etal.The relationship between plant species richness in Hulunbeier grassland and biomass and environment factors.Chinese Journal of Grassland, 2008, 30(6):74-81.
[46] Field R, Hawkins B A, Cornell H V,etal.Spatial species-richness gradients across scales:A meta-analysis.Journal of Biogeography, 2009, 36(1):132-147.
[47] Whittaker R J, Nogues-Bravo D, Araujo M B.Geographical gradients of species richness:A test of the water-energy conjecture of hawkinsetal.(2003) using european data for five taxa.Global Ecology and Biogeography, 2007, 16(1):76-89.
[48] Tasker E M, Bradstock R A.Influence of cattle grazing practices on forest understory structure in north-eastern New South Wales.Austral Ecology, 2006, 31(4):490-502.
[49] Milchunas D G, Lauenroth W K.Quantitative effects of grazing on vegetation and soils over a global range of environments.Ecological Monographs, 1993, 63(4):327-366.
[50] Pykala J.Effects of restoration with cattle grazing on plant species composition and richness of semi-natural grasslands.Biodiversity and Conservation, 2003, 12(11):2211-2226.
[51] Proulx M, Mazumder A.Reversal of grazing impact on plant species richness in nutrient-poor vs.nutrient-rich ecosystems.Ecology, 1998, 79(8):2581-2592.
[52] Thompson K, Askew A P, Grime J P,etal.Biodiversity, ecosystem function and plant traits in mature and immature plant communities.Functional Ecology, 2005, 19(2):355-358.
[53] Kassen R, Buckling A, Bell G,etal.Diversity peaks at intermediate productivity in a laboratory microcosm.Nature, 2000, 406:508-512.
[54] Grace J B, Michael Anderson T, Smith M D,etal.Does species diversity limit productivity in natural grassland communities.Ecology Letters, 2007, 10(8):680-689.
[55] Moore D R J, Keddy P A.The relationship between species richness and standing crop in wetlands the importance of scale.Vegetatio, 1988, 79(1-2):99-106.
[56] Waide R B, Willig M R, Steiner C F,etal.The relationship between productivity and species richness.Annual Review of Ecology and Systematics, 1999, 30:257-300.
[57] Ni J, Wang G H, Bai Y F,etal.Scale-dependent relationships between plant diversity and above-ground biomass in temperate grasslands, south-eastern Mongolia.Journal of arid environments, 2007, 68(1):132-142.
[58] He J S, Bazzaz F A, Schmid B.Interactive effects of diversity, nutrients and elevated CO2on experimental plant communities.Oikos, 2002, 97, 337-348.
参考文献:
[5] 左小安, 赵学勇, 赵哈林, 等.科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系.环境科学, 2007, 28(5):945-951.
[8] 白永飞, 李凌浩, 王其兵, 等.锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究.植物生态学报, 2000, 24(6):667-673.
[9] 张志南, 武高林, 王冬, 等.黄土高原半干旱区天然草地群落结构与土壤水分关系.草业学报, 2014, 23(6):313-319.
[13] 安树青, 王峥峰, 朱学雷, 等.土壤因子对次生森林群落物种多样性的影响.武汉植物学研究, 1997, 15(2):143-150.
[17] 刘旻霞, 王刚.高寒草甸植物群落多样性及土壤因子对坡向的响应.生态学杂志, 2013, 32(2):259-265.
[18] 王长庭, 龙瑞军, 曹广民, 等.高寒草甸不同类型草地土壤养分与物种多样性——生产力关系.土壤通报, 2008, 39(1):1-8.
[19] 中国科学院内蒙宁夏综合考察队.内蒙古植被[M].北京:科学出版社, 1985.
[20] 张庆, 牛建明, Buyantuyev, 等.内蒙古短花针茅群落数量分类及环境解释.草业学报, 2012, 21(1):83-92.
[21] 杨雪栋, 孙卫国, 宝音陶格涛.内蒙古荒漠草原小针茅群落土壤养分的化学计量学特征.中国草地学报, 2012, 34(5):30-34.
[22] 马文静, 张庆, 牛建明, 等.物种多样性和功能群多样性与生态系统生产力的关系——以内蒙古短花针茅草原为例.植物生态学报, 2013, 37(7):620-630.
[23] 卫智军, 韩国栋, 赵钢, 等.中国荒漠草原生态系统研究[M].北京:科学出版社, 2013.
[24] 张甘霖, 龚子同.土壤调查实验室分析方法[M].北京:科学出版社, 2012.
[27] 韩彬, 樊江文, 钟华平.内蒙古草地样带植物群落生物量的梯度研究.植物生态学报, 2006, 30(4):553-562.
[33] 张杰琦, 李奇, 任正炜, 等.氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响.植物生态学报, 2010, 34(10):1125-1131.
[37] 张林静, 岳明, 顾峰雪, 等.新疆阜康绿洲荒漠过渡带植物群落物种多样性与土壤环境因子的耦合关系.应用生态学报, 2002, 13(6):658-662.
[45] 郑晓翾, 靳甜甜, 木丽芬, 等.呼伦贝尔草原物种多样性与生物量、环境因子的关系.中国草地学报, 2008, 30(6):74-81.
Relationship between species diversity, productivity, climatic factors and soil nutrients in the desert steppe
SUN Xiao-Li1, KANG Sarula1, ZHANG Qing1,2, CHANG Chang-Ming1, MA Wen-Jing1,NIU Jian-Ming1,2*
1.CollegeofLifeSciences,InnerMongoliaUniversity,Hohhot010021,China; 2.Sino-USCenterforConservation,EnergyandSustainabilityScienceinInnerMongolia,Hohhot010021,China
The effect of species diversity on ecosystem function is one of the core issues in ecology.Our objective was to explore the relationship between species diversity, productivity, climatic factors, and soil nutrients in the desert steppe, to enhance understanding of the diversity-productivity relationship.Species richness and Shannon-Wiener index were used to measure species diversity.Climatic, vegetation and soil data were collected for 80 sampling sites across the desert steppe of Inner Mongolia.Regression analysis and stepwise regression analysis were used for determining the relationship between environmental factors and species diversity and productivity.The effects of environmental factors on species diversity and productivity were evaluated, based on the coefficient of determination from regression analysis.It was found that species diversity and biomass were both positively and linearly correlated with mean annual precipitation and aridity index.However, Shannon-Wiener index and biomass were both negatively and linearly correlated with mean annual temperature.Species richness was negatively and linearly correlated with soil total phosphorus content (R2=0.072,P<0.05), but positively and linearly correlated with soil available nitrogen content (R2=0.183,P<0.01); Shannon-Wiener index was also positively and linearly correlated with soil available nitrogen content (R2=0.061,P<0.05), and biomass was positively and linearly correlated with total nitrogen, soil available nitrogen, and soil organic carbon content.The coefficients of determination were 0.150, 0.177, 0.089 (P<0.01), respectively.Biomass increased significantly with the increase of species richness and Shannon-Wiener index.These results indicate that aridity index is the most influential factor affecting species diversity and biomass of desert steppe.
desert steppe; productivity; species diversity; soil nutrient; precipitation and temperature
10.11686/cyxb2015014
http://cyxb.lzu.edu.cn
2015-01-08;改回日期:2015-04-08
国家重点基础研究发展计划(973)项目(2012CB722201),国家科技支撑计划课题(2011BAC07B01)和国家自然科学基金(31060320)资助。
孙小丽(1988-),女,内蒙古赤峰人,在读硕士。 E-mail:sxl890325@163.com*通信作者Corresponding author.E-mail:jmniu2005@163.com
孙小丽,康萨如拉,张庆,常昌明,马文静,牛建明.荒漠草原物种多样性、生产力与气候因子和土壤养分之间关系的研究.草业学报, 2015, 24(12):10-19.
SUN Xiao-Li, KANG Sarula, ZHANG Qing, CHANG Chang-Ming, MA Wen-Jing, NIU Jian-Ming.Relationship between species diversity, productivity, climatic factors and soil nutrients in the desert steppe.Acta Prataculturae Sinica, 2015, 24(12):10-19.
