环境中的抗生素与毒物兴奋效应综述
2015-05-30葛会林等
葛会林等

摘 要 对毒物兴奋效应的出现、作用机制、普遍性及其对风险评估的影响,以及从细菌、藻类、植物、动物等不同水平对抗生素的毒物兴奋效应进行综述;并对该领域的研究进行总结和展望。
关键词 抗生素;毒物兴奋效应;风险评估;双相剂量-响应;混合物效应
中图分类号 X503.2 文献标识码 A
A Review on Hormesis and Antibiotics in the Environment
GE Huilin1, LUO Jinhui1, ZHI Xia2, LE Yuan1
1 Analysis and Testing Center, Chinese Academy of Tropical Agricultural Sciences/Hainan Provincial Key
Laboratory of Quality and Safety for Tropical Fruits and Vegetables, Haikou, Hainan 571101, China
2 College of Materials and Chemical Engineering, Hainan University, Haikou, Hainan 570228, China
Abstract The article reviewed the occurrence, mechanism of action, generalizability, and the impact on risk assessment of hormesis. The hormesis of antibiotics was reviewed from different levels including bacteria, algae, plants, and animals.
Key words Antibiotics; Hormesis; Risk assessment; Biphasci dose-response; Mixture effect
doi 10.3969/j.issn.1000-2561.2015.09.028
抗生素是由细菌、真菌、放线菌等微生物经培养而获得或用化学合成法制得的用于杀灭或抑制病原微生物的物质。我国是抗生素的生产和使用大国,如2003年我国青霉素产量达2.8万t,占世界总产量的60%,土霉素占世界总产量的65%[1]。目前,全球每年至少一半的抗生素用于畜牧业和水产养殖业,通过将抗生素添加到饲料中来预防动物疾病,或控制水产疾病。抗生素摄入后并不完全被动物所吸收,除少量抗生素残留在动物体内外,大部分则以原形化合物或代谢物的形式排泄出来,给环境微生物、土壤、水体等带来不良影响,并通过食物链影响植物和动物的正常生命活动,最终对人类的健康和生存造成不利影响[2]。抗生素在水及土壤等环境体系中都能检测到其存在,已成为一类新型环境污染物。目前关于抗生素对环境的污染及其生态毒性的认识大多来源于其急性毒性研究,对于抗生素的低剂量效应的了解仍然有限。具有低剂量促进高剂量抑制特征的毒物兴奋效应(Hormesis)[3]对抗生素的生态环境风险评估提出了新的挑战。
Hormesis一词源于希腊语“hormaein”,本意是“to excite(激活)”,目前在我国有将其译为“(毒物)兴奋效应”、“化学兴奋效应”、“激效作用”、“低促效应”、“赫米西斯”等[4]。本文采用“毒物兴奋效应”一词。我国传统医药学理论认为“药之效,毒为之”,药正是因为有毒才有了药效,即所谓“是药三分毒”;这与16世纪现代毒理学之父帕拉塞尔苏斯(Paracelsus)提出的“剂量决定毒物”,即所有物质都是有毒的,只有剂量才能区别毒物的观点不谋而合;这些观点可以认为是毒物兴奋效应概念的雏形。
1888年微生物学家Schulz发现低浓度的抗生素、氯化汞及苯酚等对酵母菌生长具有促进作用而在高浓度时表现出抑制作用[5];并于1912年在其著作中用已故同事Ardnt和自己的名字将这一现象命名为Ardnt-Schulz法则,即弱刺激加速生命力,中等强度刺激促进生命力,强刺激抑制生命力,很强刺激能致死[6]。1943年Southam和Ehrlich等在研究红雪松提取物对真菌的影响时,将观察到的双相剂量-效应关系正式命名为hormesis,首次发表在《Phytopathology》期刊上[7]。由于Ardnt-Schulz法则一直没有找到合理的机理解释,而且进入20世纪后着重强调消毒剂、杀虫剂、除草剂等在高剂量下的杀伤效应而不是关注它们的低剂量促进作用,20世纪30年代后毒物兴奋效应处于既没有追随者又没有资金资助的边缘化地位,毒物兴奋效应领域逐渐冷落下来[8]。20世纪80年代,美国环境保护署(USEPA)在评价化学物的致癌性时,将毒物兴奋效应列入考虑范围,以此来回答对于致癌物质“How clean is clean”;1982年Stebbing[9]发表毒物兴奋效应在植物生长过程中作用的综述,提出了公认的β曲线;毒物兴奋效应开始得到公认并被重视。Calabrese等对毒物兴奋效应进行了大量的研究与评述,2003年在《Nature》期刊上发表题为“Toxicology rethinks its central belief”的文章[10],至此,毒物兴奋效应进一步成为毒理学、环境科学、公共卫生等领域的研究热点。以“Hormesis”为主题词在Web of Science平台的所有数据库中进行检索,1950~1999年仅有200篇文献,而2000~2014年已有2 069篇文献。
1 毒物兴奋效应的作用机制
关于毒物兴奋效应的机理,目前有数种理论,但是这些理论尚缺乏足够的实验证据支持[11]。Calabrese[12]提出过度补偿(Overcompensation)机制,认为毒物兴奋效应是内稳态受到扰动后的响应,即生物体在最初的抑制反应之后会出现一个补偿反应,这个补偿反应会逐渐超过控制行为,从而导致一个净刺激效应。当毒物的抑制作用小于机体产生的补偿作用时,表现为低剂量促进作用;随着毒物剂量的增加,补偿作用逐渐被抵消,整体效果转变成高剂量抑制作用。Stebbing[9]提出矫正过度(Overcorrection)机制,认为毒物在高剂量时一般对生物产生抑制作用,刺激作用可能是生物对低剂量抑制的自我矫正反应,即生物体对毒物抑制作用的过度中和与抵抗。毒物低剂量时总体表现为促进;随着毒物剂量的增加,抵抗中和作用到达临界最大值后就不再增加,最终对生物体产生抑制作用。
上述2种机制都停留在定性的描述性层面,未从分子机制方面对对毒物兴奋效应做出合理的解释。因此有人提出了多亚型受体机制[13],认为毒物兴奋效应是由一类同工G蛋白偶联受体(GPCR)介导的,GRCP谱中每一个亚型激活其所对应的信号传导通路,选择性的控制生物反应。根据产生的效应不同,该同工受体谱分为3类,即兴奋受体、抑制受体和过渡受体。兴奋受体的亲和力较高,数量较少,能够激活有益的信号传导通路;抑制受体的亲和力较低,数量较多,能够激活不利的传导通路;过渡受体亲和力居中,数量不定,只是单纯与毒物结合,整体上起到调控兴奋效应向抑制效应转变的作用[14]。当少量的毒物作用于机体时,首先与兴奋受体相结合,激活有益的信号传导通路,产生正向效应;随着毒物剂量的增加,毒物逐步与过渡受体和抑制受体相结合,同时激活不利的信号传导通路,产生的负向效应逐渐将正向效应抵消,最后总体表现为抑制作用。在不同生物学模型中发现核转录因子E2相关因子2(Nrf2)、叉头转录因子(FOXO)、核转录因子(NF-κB)和热休克转录因子(HSF)等周期性的激活对于各种毒物兴奋的刺激效应是必要的[15]。
Conolly等[16]根据2个药理学数据集与2个毒理学数据集提出了4个解释双相剂量-效应曲线的生物学机制。(1)苯基异丙腺苷(PIA)对大鼠脑中腺苷酸环化酶活性的影响研究证实脑纹状体中环磷腺苷(cAMP)的形成具有非单调剂量-效应曲线,解释为腺苷酸环化酶在相反方向调控的2个腺苷受体亚型的拮抗作用,导致在配体亲和力与信号转导效能方面的一定差异。(2)在HepG2人肝癌细胞中,雄激素受体羟基氟他胺(HOF)介导的基因转录呈现非单调剂量-效应曲线,低浓度的HOF部分拮抗初始剂量的双氢睾酮(DHT)的效应,而进一步增加HOF浓度则表现为激动活性;解释为携带2个DHT或2个HOF配体的二聚体受体而非混合配体的二聚体有转录活性。(3)外源性致癌物诱导DNA修复形成DNA加合物的过程中,诱导修复能力不仅修复外源DNA加合物,而且修复背景DNA损伤。在低剂量时,修复诱导的有益效应能过度补偿增加的损伤。一旦修复诱导饱和,在高剂量外源物下观察到总DNA损伤(背景加外源)将高于背景水平,从而导致双相剂量-效应关系。(4)细胞分裂作为DNA损伤修复的一个调节因子,低水平的DNA损伤假定减缓细胞周期,以使复制前能进行更完全的DNA修复。而高水平的DNA损伤是有细胞毒性的,为此细胞分裂加速实现再生性增生。随着DNA损伤剂的剂量增加,细胞分裂速率与突变速率显示出双相响应。
2 毒物兴奋效应对风险评估的影响
传统上用于化学污染物风险评估的基本模型有两种,如图1所示,一种是线性阈值模型(Linear Threshold model,LT),它假设化学物在一定的暴露浓度下是安全的,即在阈剂量以下不存在风险,一般用于非致癌性物质的风险评估;另一种是线性非阈值模型(Linear Non-Threshold model,LNT),假设只有当剂量为零时产生的风险才为零,一般用于化学致癌物质及辐射的风险评估[17]。具有双相特征的毒物兴奋效应模型假设存在零等价剂量(Zero Equivalent Dose,ZED),ZED点以上为有害效应,ZED点以下则呈有益效应,毒物兴奋效应模型否定了低剂量效应能从高剂量数据进行外推的标准假设,这无疑对传统线性剂量-效应模型形成巨大挑战。Calabrese[18]认为应当将毒物兴奋效应模型作为毒理学、药理学与生态风险评价的中心法则。
一般在非致癌物健康风险评估中,人类健康阈值制定采用的是动物的无观察效应浓度(NOEC)除以安全系数获得,安全系数一般定为100(10×10,分别代表种间差异与种内差异),这样所得的阈值不仅可能低于人的NOEC,而且也可能低于对人类健康的最佳有益浓度[19]。为此,Calabrese[20]提出为充分发挥毒物兴奋效应的有益作用,应该将安全系数改为50。传统的风险评估外推是以线性模型为基础的, 而根据毒物兴奋效应模型可知在低剂量下的毒性反应并不遵循线性规律,提示以往所制定的卫生标准存在商榷之处,毒物兴奋效应模型将会对污染物残留卫生标准的制定带来一定影响[21]。
毒物兴奋效应强调了污染物存在阈值,这就引发这样一个讨论:环境应该治理到什么程度, 也就是说多清洁才算清洁? 公众心目中零污染是最安全的,但是如果遵循毒物兴奋效应规律,污染物在低剂量时存在毒物兴奋效应并不一定是一件坏事,管理部门可有的放矢地控制污染,既能有效控制风险,又能控制成本。毒物兴奋效应的双相剂量-效应模型比传统单调剂量-效应模型更具有预测性。接受毒物兴奋效应的理论,在进行低剂量化学物风险评估时,将可能放宽大气、水体、食品和土壤的污染物暴露标准,并可削减环境管理的费用。
毒物兴奋效应的观点将彻底改变向公众进行风险交流的策略[21]。风险交流是在风险分析全过程中,风险评估专业人员、风险管理行政人员、消费者、企业、学术界和其他利益相关方就某项风险、风险所涉及的因素和风险认知相互交换信息和意见的过程,内容包括风险评估结果的解释和风险管理决策的依据[22]。正如现在已知长期大量饮酒可增加患高血压、食管癌、肝硬化和肝癌的风险,而少量适宜饮酒可降低冠心病和脑中风的发生率;砒霜(三氧化二砷)被国际癌症研究中心(IARC)定义为一级致癌物,但低剂量的砒霜可用于治疗白血病。过去环境管理部门、公共卫生机构和公众只强调化学物带来的环境与食品安全风险,如果毒物兴奋效应观点被采纳,公共风险管理与交流的面貌将会完全改观。
在医学上,许多抗生素和抗肿瘤药物都表现出双相剂量-响应,一个剂量可能是临床有效的, 但另一剂量则可能是有害的。如抗肿瘤药苏拉明在高剂量下抑制癌细胞增殖,但其在人体中的残留时间长达40~50 d,这段时间内癌细胞将有可能处于苏拉明的毒物兴奋效应浓度区域而促进癌细胞增殖[23]。体内试验表明非常低剂量的常用抗生素如青霉素[24]和链霉素[25]可以增加感染伤寒埃氏杆菌的小白鼠的死亡率,而在较高剂量时却可以降低其死亡率。因此,毒物兴奋效应的存在不仅为完善临床治疗方案提供了新的机会,同时也提出了新的挑战。
在农业上,已发现一些农药对昆虫的活力能导致毒物兴奋效应,在使用农药特别是杀虫剂时值得关注。如吡虫啉对目标害虫桃蚜的生殖具有毒物兴奋效应,高剂量时显著抑制桃蚜生殖力,但低剂量时(0.000 1~0.1 mg/kg)能刺激桃蚜产卵行为[26]。低剂量(0.001~0.1 mg/kg)吡虫啉能显著提高瓢虫的捕食量及相对活力,从而增强其控害效能[27]。蓟马取食含有少量马拉硫磷的柑桔叶片后,雌虫产卵能力显著高于未处理的雌虫[28]。Morse[29]建议进行农药登记时,为全面了解农药的可能影响,需要考虑农药的毒物兴奋效应。
毒物兴奋效应中有益效应的评判应进行综合考虑,大量事实表明简单的刺激作用并不总是有利。细胞色素P450酶、谷胱甘肽-S-转移酶(GST)是动物体内的重要解毒酶系,一些毒物作用机体后会导致这些酶系活力的升高,这正是毒物发挥有害作用导致的;而且P450、GST酶活力的升高会增加致突变物损伤DNA的风险[30]。类似地,在某一剂量下毒物可能对生物体内某些指标有益, 而对另一些生物指标却不利。如低剂量2,3,7,8-四氯二苯并二噁英(TCDD)可使脑垂体、子宫、乳腺和胰腺的肿瘤发生率降低,却使肝、肺、舌和鼻咽部的肿瘤发生率增加[8]。危险度确定应根据剂量对健康的净作用而不是单一剂量对单一疾病终点的作用,如总的死亡率而不是特定的致死性疾病[31]。
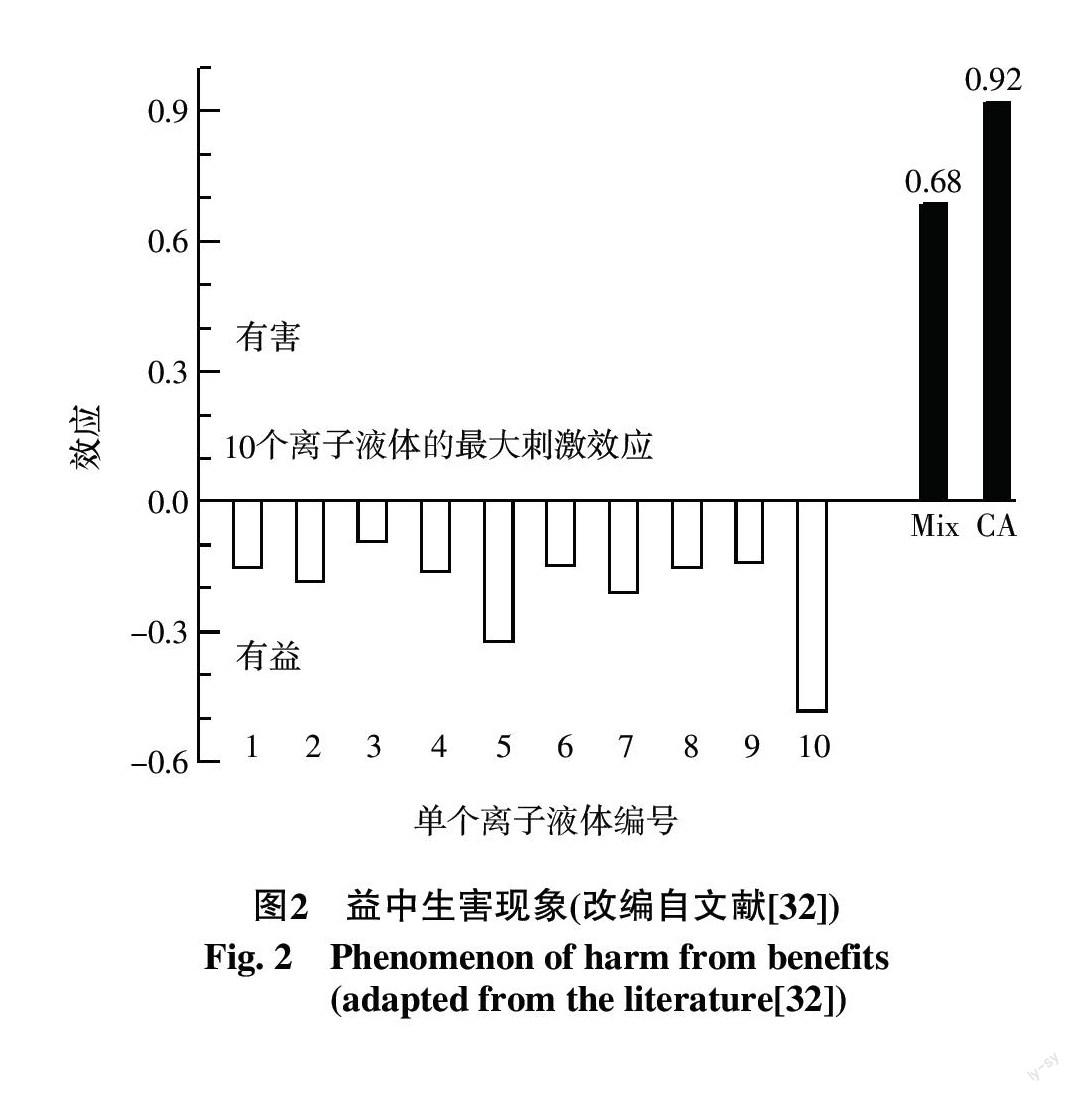
生物体一般都是暴露于多种环境污染物构成的混合物中。Ge等[32]应用浓度加和(CA)模型预测了10个离子液体混合物对荧光素酶的毒性,观察到离子液体组分均呈现最大刺激效应时,其混合物却表现出较高的抑制效应(图2)。这种"益中生害"的现象在单调剂量-效应模型框架下是不可理解和无法预测的,所以毒物兴奋效应中的有益效应还应放在混合物背景下进行评判,在毒物兴奋效应框架下建立污染物残留限量标准时应考虑混合暴露因素。关于混合物毒物兴奋效应,目前讨论的焦点有实践中是否需要使用双相剂量-效应模型进行有害效应风险评价;混合物毒物兴奋效应的刺激幅度和刺激范围是否能预测;混合物的毒物兴奋效应是否总能预测;双相剂量-效应模式下协同与拮抗如何定义等。由于混合物中组分的最大刺激效应存在差异,CA模型存在某些预测盲区,这是今后混合物毒物兴奋效应预测与评估中需要解决的关键问题。
3 抗生素的毒物兴奋效应
抗生素对动物的一些生理指标能导致毒物兴奋效应。Flaherty等[33]观察到36 μg/L氟西汀30 d慢性暴露能显着增加大型蚤产卵量,10 μg/L氯贝酸6 d急性暴露能显著增加其性别比例,解释为大型蚤繁殖力的增加可能是氟西汀慢性暴露导致体内平衡破坏后矫枉过正的反映,即在有限的时间内对低水平胁迫的适应性响应。丁君等[34]研究了四种抗生素对海胆抗氧化酶和脂质过氧化的影响,结果表明海胆体液超氧化物歧化酶(SOD)和过氧化氢酶(CAT)的活性随处理时间呈先增加后下降的趋势且在第9天达到最大活性;丙二醛(MDA)含量随处理时间呈上升趋势,毒物兴奋效应发生在实验早期阶段;处理15 d后,盐酸四环素和氨苄青霉素作用的MDA含量高于对照。环丙沙星对大鼠星形胶质细胞增殖[35]与人类成纤维细胞存活率[36]能导致毒物兴奋效应,分析可能是活性氧、细胞增殖和细胞活力之间复合的剂量-效应曲线所致。
抗生素对植物的生长与藻类的繁殖能产生毒物兴奋效应。浓度50~5 000 μg/L的氟甲喹对千屈菜暴露35 d后,能增加菜叶和次生根的平均数目和直径,在50 μg/L时刺激效应最大[37]。盐酸金霉素和土霉素对玉米的主根长度、地上部分长度、叶片数目能产生毒物兴奋效应[38]。恩诺沙星对黄瓜、莴苣、菜豆与萝卜的初生根、下胚轴和子叶的长度以及叶片的数目/长度都能导致毒物兴奋效应[39]。盐酸环丙沙星、盐酸土霉素和磺胺二甲嘧啶在高浓度(>10 μg/L)对芦苇的根系活力和叶片叶绿素具有毒性,而在低浓度(0.1~1 μg/L)则发生刺激效应[40]。诺氟沙星对芽期玉米的根和芽中MDA含量以及过氧化物酶(POD)、GST、SOD和CAT等抗氧化酶的活性具有毒物兴奋效应[41]。10 μg/L的四环素对浮萍的刺激效应达到26%,10 μg/L的布洛芬对蓝藻的刺激效应达到72%[42]。诺氟沙星对蛋白核小球藻的7-乙氧基异吩恶唑酮脱乙基酶(EROD)和CAT活性能产生毒物兴奋效应[43]。头孢拉定能刺激羊角月牙藻的生长,头孢氨苄和7-氨基头孢霉烷酸对羊角月牙藻具有明显的毒物兴奋效应[44]。链霉素对亚心形扁藻、新月菱形藻、小球藻、金藻8701、三角褐指藻和角毛藻的生长具有刺激效应[45]。研究表明刺激植物生长的分子靶位是烟酰胺腺嘌呤二核苷酸(NADH)氧化酶(NOX),一些溶剂如醋酸乙酯、丙酮、苯、二甲苯、氯仿和四氯化碳在低浓度时能增加与生长有关的NOX蛋白质的活性[46]。而NOX的活性与细胞生长速率密切相关,能够决定细胞的分裂速度[47]。
抗生素对微生物的生理活动也能产生毒物兴奋效应。抗生素的混合物对着生藻类能导致毒物兴奋效应[48]。磺胺甲恶唑、磺胺吡啶、磺胺嘧啶、磺胺异恶唑、磺胺间甲氧嘧啶和磺胺氯哒嗪对明亮发光杆菌能导致毒物兴奋效应[49]。磺胺甲恶唑、甲氧苄啶、磺胺甲氧哒嗪及其混合物对费氏弧菌能产生毒物兴奋效应[50]。土霉素对青海弧菌与费氏弧菌能导致毒物兴奋效应[51]。硫酸安普霉素和硫酸巴龙霉素对青海弧菌的24 h毒性呈现毒物兴奋效应[52],这反映了毒物兴奋效应的时间依赖性。四环素在远低于最小抑制浓度(4 μg/mL)的范围(0.015~0.03 μg/mL)对大肠杆菌呈现毒物兴奋效应菌落形成单位(CFU)相对空白上升到141%~121%[53]。磺胺嘧啶、磺胺甲恶唑和土霉素能抑制土壤细菌和放线菌生长,使土壤微生物的生物量明显下降,但能促进土壤真菌生物量增加[54]。Linares等[55]发现妥布霉素、四环素、环丙沙星对铜绿假单胞菌生物膜形成能力具有毒物兴奋效应,认为抗生素应当看作是调节微生物群落动态平衡的信号分子。Calabrese等[56]研究了1 888种抗菌剂在低于毒理学阈值以下的浓度对大肠杆菌生长的影响,发现处理的响应显著大于空白,符合毒物兴奋效应特征。
抗生素对动物、植物、微生物等均能导致毒物兴奋效应,毒物兴奋效应具有一定的普遍性。Calabrese[57]认为应将毒物兴奋效应模型视为剂量-效应关系的预设形式,污染物对生物体的毒物兴奋效应是普遍存在的,与所研究的生物模型、观察终点、受试化学物质的种类无关。但这并不意味着毒物兴奋效应在自然界与文献中出现的频率是一样的,已经检验的例子证实该数值在20%~25%或更低,而毒物兴奋效应在人体中令人信服的证据则更少[58]。此外,单终点的毒物兴奋效应不能用于解释多毒性终点、靶器官和所有机理,所以不应将毒物兴奋效应机理全能化,毒物兴奋效应只有与具体的毒性终点与分子机制结合起来讨论才有意义,否则毒物兴奋效应的普遍性将丧失建立的基础。
为了寻找化学物的毒物兴奋效应,应当应用优化试验设计的方法。正常情况下毒物兴奋效应的刺激范围窄,通常只有一个数量级;刺激幅度很小,只比对照组高30%~60%,因此需要有巧妙的实验方案设计,来避免错过毒物兴奋效应的最佳观测时机[5]。毒物兴奋效应的剂量通常比NOEC低10倍左右,要想得到理想的毒物兴奋效应曲线,需要按照剂量等比稀释等优化方法选择剂量范围,设计合适的剂量组数和剂量间距来进行试验。其次,暴露时间是一个重要影响因素。如一些抗生素在15 min发光菌毒性试验中没有毒物兴奋效应,而在24 h发光菌毒性试验中则出现毒物兴奋效应。再次,指示生物和毒性终点的选择也很重要。如双酚A对人乳腺癌细胞增殖具有毒物兴奋效应,而对发光菌发光则表现为抑制效应。磺胺二甲氧嘧啶对千屈菜的根、子叶、子叶叶柄总是呈现毒性作用,而对节间和叶片长度表现出剂量依赖的毒物兴奋效应[59]。
4 总结与展望
综上所述,尽管抗生素在环境中的浓度较低,但其对环境生态的威胁和人类健康的影响是巨大的。早在上世纪90年代欧盟各国已经开始了对环境中抗生素风险评估的研究,并对医用抗生素和兽用抗生素做了严格的限定;美国在2001年开始水体中抗生素残留的大范围调查和研究[60]。我国抗生素的生产及使用情况较欧盟和美国更甚,更需重视其在环境中的生物效应研究,将来应进一步关注以下几个方面。
(1)抗生素对有机体低浓度长期暴露的生物效应。目前对抗生素生物效应的了解多是来自高浓度抗生素对水生生物和微生物的急性毒性数据,这显然与实际环境中抗生素的低浓度长期残留状况存在较大差异,同时由于毒物兴奋效应的存在,由高浓度急性毒性数据外推至低浓度慢性暴露时需要十分小心。随着环境卫生安全标准的提高,研究抗生素低浓度长期暴露的生物效应已迫在眉睫。
(2)抗生素毒物兴奋效应的作用机制。毒物兴奋效应显示出剂量-效应关系从单调向双相模式的转变,对毒理学、药理学、临床医学和农药学等许多方面都产生影响,将可能改变传统的风险评估模式[61]。通过融入环境基因组学与毒理基因组学等方法,对抗生素毒物兴奋效应的作用机制将有更加完整的认识,从而对更加合理有效的使用抗生素做出指导。
(3)抗生素混合物的环境生态效应。环境中的抗生素主要是以混合物形式存在于水体或土壤中,抗生素间发生协同或拮抗的毒性相互作用现象也较普遍。因此,还需要对实际环境中浓度多变、组分复杂的抗生素混合物的生物效应开展预测与评估工作。
参考文献
[1] 邹小明, 肖春玲, 肖 晓. 水域中抗生素生态毒理效应研究进展[J]. 井冈山大学学报(自然科学版), 2012, 33(1): 47-52.
[2] 周启星, 罗 义, 王美娥. 抗生素的环境残留、 生态毒性及抗性基因污染[J]. 生态毒理学报, 2007, 2(3): 243-251.
[3] Davies J, Spiegelman G B, Yim G. The world of subinhibitory antibiotic concentrations[J]. Current Opinion in Microbiology, 2006, 9(5): 445-453.
[4] 张玉彬, 金泰虞, 金锡鹏. Hormesis简史及其中文译名[J]. 环境与职业医学, 2005, 22(6): 538-539.
[5] Calabrese E J, Baldwin L A. Hormesis as a biological hypothesis[J]. Environmental Health Perspectives, 1998, 106(Suppl 1): 357-362.
[6] 王丽娟. 含J-型浓度-效应谱特征混合物的毒性相互作用研究[D]. 上海: 同济大学, 2011.
[7] Southam C M, Ehrlich J. Effects of extract of western red-cedar heartwood on certain wood-decaying fungi in culture[J]. Phytopathology, 1943, 33: 517-524.
[8] 龚春梅, 庄志雄. 浅议毒物兴奋效应[J]. 中山大学研究生学刊, 2007, 28(4): 24-34.
[9] Stebbing A R D. Hormesis-the stimulation of growth by low levels of inhibitors[J]. Science of the Total Environment, 1982, 22(3): 213-234.
[10] Calabrese E J, Baldwin L A. Toxicology rethinks its central belief-Hormesis demands a reappraisal of the way risks are assessed[J]. Nature, 2003, 421(6924): 691-692.
[11] 郭兰萍, 张小波, 杨 光, 等. Hormesis及其在药用植物生产中的应用[J]. 中国中药杂志, 2011, 36(5): 525-529.
[12] Calabrese E J. Evidence that hormesis represents an “overcompensation” response to a disruption in homeostasis[J]. Ecotoxicology and Environmental Safety, 1999, 42(2): 135-137.
[13] Chadwick W, Maudsley S. Hormesis-The devil is in the dose: complexity of receptor systems and responses[M]. New York: Humana Press, 2010: 95-108.
[14] 范萌知, 张一倩, 刘幸俊. Hormesis现象及其同工G-蛋白耦合受体机制[J]. 毒理学杂志, 2014, 28(2): 159-163.
[15] Son T G, Cutler R G, Mattson M P, et al. Hormesis-Transcriptional mediators of cellular hormesis[M]. New York: Humana Press, 2010: 69-93.
[16] Conolly R B, Lutz W K. Nonmonotonic dose-response relationships: mechanistic basis, kinetic modeling, and implications for risk assessment[J]. Toxicological Sciences, 2004, 77(1): 151-157.
[17] 让欧艳, 让蔚清. 低剂量兴奋效应及其医学应用[J]. 实用预防医学, 2009, 16(1): 300-303.
[18] Calabrese E J. Hormesis is central to toxicology, pharmacology and risk assessment[J]. Human & Experimental Toxicology, 2010, 29(4): 249-261.
[19] 顾晓军, 田素芬. 毒物兴奋效应概念及其生物学意义[J]. 毒理学杂志, 2007, 21(5): 425-428.
[20] Calabrese E J. Expanding the reference dose concept to incorporate and optimize beneficial effects while preventing toxic responses from nonessential toxicants[J]. Regulatory Toxicology and Pharmacology, 1996, 24(1): S68-S75.
[21] 戴宇飞, 郑玉新. 毒理学中心法则的重新审视-毒物兴奋性剂量-反应关系及其对毒理学发展的影响[J]. 国外医学卫生学分册, 2003, 30(4): 246-249.
[22] 厉曙光. 食品安全与风险交流[J]. 环境与职业医学, 2014, 31(10): 762-763.
[23] Calabrese E J, Mattson M P. Hormesis-The hormetic pharmacy: the future of natural products and man-made drugs in disease prevention and treatment[M]. New York: Humana Press, 2010: 176-198.
[24] Randall W A, Price C W, Welch H. Demonstration of hormesis (increase in fatality rate)by penicillin[J]. American Journal of Public Health and the Nations Health, 1947, 37(4): 421-425.
[25] Welch H, Price C W, Randall W A. Increase in fatality rate of E. typhosa for white mice by streptomycin[J]. Journal of the American Pharmaceutical Association, 1946, 35(5): 155-158.
[26] 余月书, 沈国清, 陆贻通. 吡虫啉对桃蚜生殖力的Hormesis效应及对天敌控害效能的影响[J]. 科技通报, 2010, 26(6): 879-883.
[27] 赵 明, 沈国清, 陆贻通, 等. 低剂量吡虫啉对异色瓢虫的Hormesis效应及其抗氧化酶活性的影响[J]. 扬州大学学报, 2012, 33(4): 77-80.
[28] Morse J G, Zareh N. Pesticide-induced hormoligosis of citrus thrips (Thysanoptera: Thripidae) fecundity[J]. Journal of Economic Entomology, 1991, 84(4): 1 169-1 174.
[29] Morse J G. Agricultural implications of pesticide-induced hormesis of insects and mites[J]. Human & Experimental Toxicology, 1998, 17: 266-269.
[30] Thayer K A, Melnick R, Burns K, et al. Fundamental flaws of hormesis for public health decisions[J]. Environmental Health Perspectives, 2005, 113(10): 1 271-1 276.
[31] 刘秀英, 胡怡秀. 毒理学默认量效关系模型的重新审视[J]. 国外医学: 卫生学分册, 2008, 35(5): 257-263.
[32] Ge H L, Liu S S, Zhu X W, et al. Predicting hormetic effects of ionic liquid mixtures on luciferase activity using the concentration addition model[J]. Environmental Science & Technology, 2011, 45(4): 1 623-1 629.
[33] Flaherty C M, Dodson S I. Effects of pharmaceuticals on Daphnia survival, growth, and reproduction[J]. Chemosphere, 2005, 61(2): 200-207.
[34] 丁 君, 孙雪峰, 张 勃, 等. 以海胆为模式生物的抗生素对海水养殖生物抗氧化酶及膜脂质过氧化影响研究[C]. 环境污染与大众健康学术会议(CEPPH), 2010: 310-315.
[35] Gürbay A, Gonthier B, Barret L, et al. Cytotoxic effect of ciprofloxacin in primary culture of rat astrocytes and protection by vitamin E[J]. Toxicology, 2007, 229(1): 54-61.
[36] Hincal F, Gürbay A, Favier A. Biphasic response of ciprofloxacin in human fibroblast cell cultures[J]. Nonlinearity in Biology, Toxicology, and Medicine, 2003, 1(4): 481-492.
[37] Migliore L, Cozzolino S, Fiori M. Phytotoxicity to and uptake of flumequine used in intensive aquaculture on the aquatic weed, Lythrum salicaria L[J]. Chemosphere, 2000, 40(7): 741-750.
[38] Migliore L, Godeas F, De Filippis S P, et al. Hormetic effect(s) of tetracyclines as environmental contaminant on Zea mays[J]. Environmental Pollution, 2010, 158(1): 129-134.
[39] Migliore L, Cozzolino S, Fiori M. Phytotoxicity to and uptake of enrofloxacin in crop plants[J]. Chemosphere, 2003, 52(7): 1 233-1 244.
[40] Liu L, Liu Y H, Liu C X, et al. Potential effect and accumulation of veterinary antibiotics in Phragmites australis under hydroponic conditions[J]. Ecological Engineering, 2013, 53: 138-143.
[41] 王 朋, 温 蓓, 张淑贞. 诺氟沙星对芽期玉米的毒性和氧化损伤研究[J]. 生态毒理学报, 2010, 5(6): 849-856.
[42] Pomati F, Netting A G, Calamari D, et al. Effects of erythromycin, tetracycline and ibuprofen on the growth of Synechocystis sp. and Lemna minor[J]. Aquatic Toxicology, 2004, 67(4): 387-396.
[43] 聂湘平, 鹿金雁, 李 潇, 等. 诺氟沙星(Norfloxacin)对蛋白核小球藻(Chlorella pyrenoidosa)生长及抗氧化酶活性的影响[J]. 生态毒理学报, 2007, 2(3): 327-332.
[44] 高 礼, 石丽娟, 袁 涛. 典型抗生素对羊角月牙藻的生长抑制及其联合毒性[J]. 环境与健康杂志, 2013, 30(6): 475-478.
[45] 黄 健, 宫相忠, 唐学玺, 等. 链霉素对海洋微藻的毒物刺激效应[J]. 青岛海洋大学学报(自然科学版), 2000, 30(4): 642-644.
[46] 王东红, 彭 安, 王子健. 有毒物质低剂量刺激作用的研究进展[J]. 安全与环境学报, 2004, 4(1): 18-21.
[47] Morre D J. Chemical hormesis in cell growth: A molecular target at the cell surface[J]. Journal of Applied Toxicology, 2000, 20: 157-163.
[48] Backhaus T, Porsbring T, Arrhenius A, et al. Single-substance and mixture toxicity of five pharmaceuticals and personal care products to marine periphyton communities[J]. Environmental Toxicology and Chemistry, 2011, 30(9): 2 030-2 040.
[49] Deng Z Q, Lin Z F, Zou X M, et al. Model of hormesis and its toxicity mechanism based on quorum sensing: a case study on the toxicity of sulfonamides to Photobacterium phosphoreum[J]. Environmental Science & Technology, 2012, 46(14): 7 746-7 754.
[50] Zou X, Lin Z, Deng Z, et al. Novel approach to predicting hormetic effects of antibiotic mixtures on Vibrio fischeri[J]. Chemosphere, 2013, 90(7): 2 070-2 076.
[51] 王 婧.费氏弧菌和青海弧菌在抗生素检测技术中的应用研究[D]. 杨凌: 西北农林科技大学, 2011.
[52] 林 楠, 张 晶, 刘树深. 污染物对青海弧菌Q67的时间依赖微板毒性分析[J]. 中国科技论文在线精品论文, 2013, 6(24): 2 321-2 328.
[53] Migliore L, Rotini A, Thaller M C. Low doses of tetracycline trigger the E. coli growth: A case of hormetic response[J]. Dose-Response, 2013, 11(4): 550-557.
[54] Yang Q, Zhang J, Zhu K, et al. Influence of oxytetracycline on the structure and activity of microbial community in wheat rhizosphere soil[J]. Journal of Environmental Sciences, 2009, 21(7): 954-959.
[55] Linares J F, Gustafsson I, Baquero F, et al. Antibiotics as intermicrobial signaling agents instead of weapons[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(51): 19 484-19 489.
[56] Calabrese E J, Hoffmann G R, Stanek E J, et al. Hormesis in high-throughput screening of antibacterial compounds in E. coli[J]. Human & Experimental Toxicology, 2010, 29(8): 667-677.
[57] Calabrese E J. Hormesis: from marginalization to mainstream- A case for hormesis as the default dose-response model in risk assessment[J]. Toxicology and Applied Pharmacology, 2004, 197(2): 125-136.
[58] Mushak P. Hormesis and its place in nonmonotonic dose-response relationships: Some scientific reality checks[J]. Environmental Health Perspectives, 2007, 115(4): 500-506.
[59] Migliore L, Rotini A, Cerioli N L, et al. Phytotoxic antibiotic sulfadimethoxine elicits a complex hormetic response in the weed Lythrum salicaria L[J]. Dose-Response, 2010, 8(4): 414-427.
[60] 贾江雁, 李明利. 抗生素在环境中的迁移转化及生物效应研究进展[J]. 四川环境, 2011, 30(1): 121-125.
[61] 杨晓华, 庄志雄. 毒物刺激作用的研究进展[J]. 中华预防医学杂志, 2006, 40(1): 60-61.
