长江下游湖泊水生植物现状及与水环境因子的关系*
2015-05-10孔祥虹肖兰兰苏豪杰张霄林李中强
孔祥虹,肖兰兰,苏豪杰,吴 耀,张霄林*,李中强**
(1: 湖北大学资源环境学院,武汉 430062)
(2: 郑州幼儿师范高等专科学校,郑州 450002)
(3: 中国科学院水生生物研究所东湖湖泊生态系统试验站,武汉 430072)
长江下游湖泊水生植物现状及与水环境因子的关系*
孔祥虹1,肖兰兰2,苏豪杰3,吴 耀3,张霄林3*,李中强1**
(1: 湖北大学资源环境学院,武汉 430062)
(2: 郑州幼儿师范高等专科学校,郑州 450002)
(3: 中国科学院水生生物研究所东湖湖泊生态系统试验站,武汉 430072)
研究水生植物分布与环境因子的关系可为富营养化湖泊的生态修复提供重要科学依据. 通过对长江下游10个不同营养水平湖泊的水生植物群落组成和环境状况进行野外调查,研究了长江下游湖泊主要水生植物分布状况及水环境因子对水生植物分布的影响. 调查发现长江下游10个代表性湖泊主要水生植物共计6科7属11种,主要生活型为沉水植物. 水生植物群落组成与环境因子的冗余分析结果显示,总氮、pH值和水深是显著影响这些不同营养水平湖泊水生植物分布的主导因子.
水生植物;环境因子;营养水平;主要因子;冗余分析;长江下游
水体富营养化、过度养殖以及人类生产活动引起的水体污染等导致的水质恶化和水生生态系统破坏是当前淡水湖泊环境面临的主要问题[1]. 水生植物不仅是淡水湖泊的初级生产者和水体净化者,也是湖泊生态平衡的调控者,对水生生态系统的结构和功能具有重要影响[2-3]. 其变化可以直接或间接地反映湖泊环境的状况及发展趋势[4]. 许多研究表明温度、透明度、水深、营养盐浓度、底泥等环境因子显著影响水生植物的生长分布[4-14]. 近年来,由于大量排放生活污水、工业废水以及不合理的大规模湖泊围垦,导致湖泊生态系统结构和功能不断退化,水生生物的群落受到严重影响[15]. 研究发现近50年长江中下游湖泊生物多样性显著下降[16]. 过去关于环境因子对水生植物生长分布的研究主要为室内控制实验、室外围隔实验以及单个湖泊的野外水生植物[5-6,12-14,17-19],大多考虑一个或少数几个环境因子对水生植物生长和分布的影响,较少考虑到自然水体环境因子的综合作用.
对不同营养水平的湖泊水生植物分布状况进行比较研究可以了解不同营养类型湖泊的水生植物分布、生长状况,分析其生长、分布与水环境因子的关系,阐明影响水生植物分布的主导因子,从而为富营养湖泊水生植物生态恢复提供重要的科学依据. 本研究拟选择长江下游10个具有代表性的不同营养类型淡水湖泊,调查其水生植物分布、生长状况及水环境现状,运用RDA冗余分析阐明水环境因子对水生植物生长分布的影响,以期为进一步研究水生植物的生境需求以及长江下游湖泊水生植物的生态恢复提供基础资料.
1 研究方法
1.1 研究区域
于2013年6-7月在长江下游地区选取10个具有代表性的淡水湖泊,这些湖泊代表了除重度富营养水平外(敞水区基本无水生植物)的长江下游湖泊的不同营养现状(中营养、轻度富营养和中度富营养)[20-21]. 根据所选湖泊大小、受干扰程度及水生植物实际分布情况,采用样点调查法,10个湖泊共设置采样点29个(表1),对所选湖泊进行水生植物的群落组成及水环境因子调查.
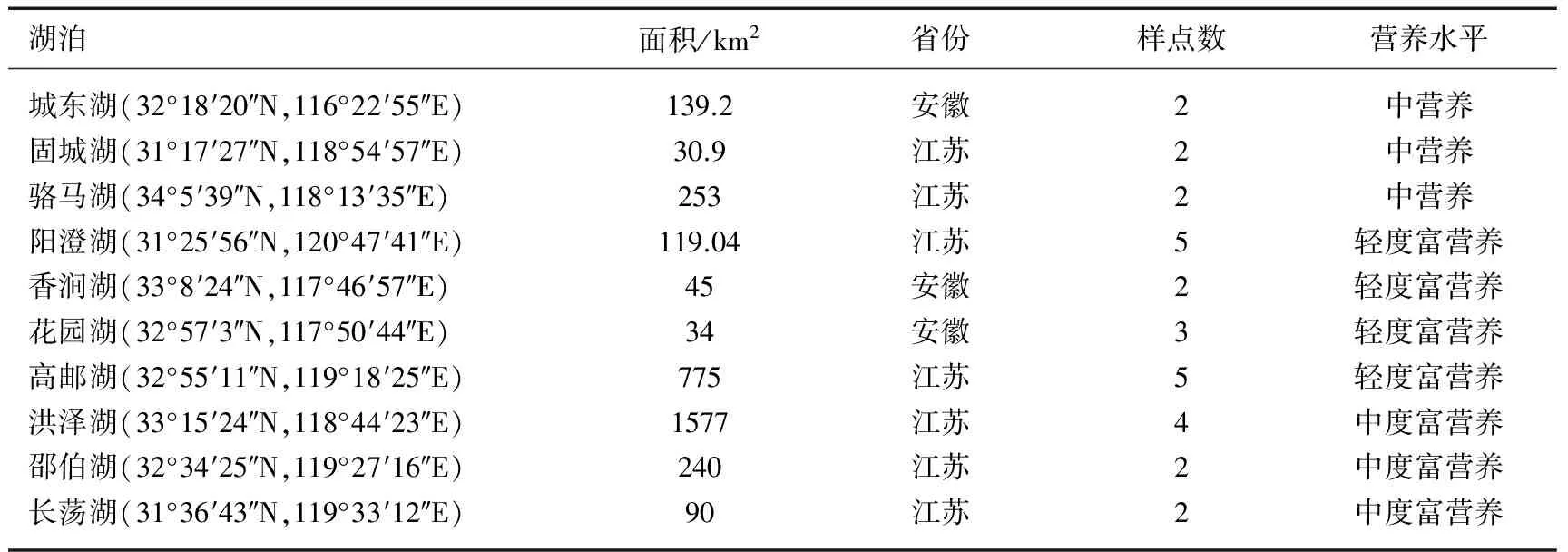
表1 调查湖泊位置、营养状况及样点数设置*
*营养水平数据引自文献[20-21].
1.2 环境因子的测定
现场采用塞氏盘测定透明度(SD),用YSI多参数水质监测仪现场测定不同水深(0.5~2.0 m)的水温(T)、pH值、电导率(EC)、氧化还原电位(ORP)、溶解氧(DO)、水深(Depth)等参数. 用采水器采集2 L混合水样带回实验室,测定总磷(TP)、总氮(TN)和叶绿素a(Chl.a)含量,水化指标的测定参照《湖泊富营养化调查规范》[16].
1.3 水生植物的采集及处理
根据湖泊面积大小,在湖泊水生植物生长良好的水域均匀布置采样点2~5个. 现场记录水生植物种类,目测法估测植物盖度、多度. 采用重要值方法确定优势种:重要值=(相对盖度+相对多度+相对频度)/3,频度=物种出现的样方数/整个样方数×100. 每个位点生物量用50 cm×50 cm的采草器随机采集3次,将所有植物连根夹起并及时冲洗干净、分类,风干15min后称重,以湿重代表生物量,根据所测物种湿重平均值和各物种盖度,计算单位面积生物量.
1.4 湖泊营养程度评价方法
本文采用综合营养指数法作为评价湖泊富营养化的方法[22-23]. 综合营养状态指数计算公式为:
TLI(Chl.a)=10(2.5+1.086ln Chl.a)
(1)
TLI(TP)=10(9.436+1.624ln TP)
(2)
TLI(TN)=10(5.453+1.694ln TN)
(3)
TLI(SD)=10(5.118-1.94ln SD)
(4)
TLI(∑)=W(Chl.a)·TLI(Chl.a)+W(TP)·TLI(TP)+W(TN)·TLI(TN)+W(SD)·TLI(SD)
(5)
式中,W(Chl.a)、W(TP)、W(TN)和W(SD)分别为0.3260、0.2300、0.2192和0.2246. 采用0~100的一系列连续数字对湖泊营养状态进行分级:TLI(∑) ≤30为贫营养;TLI(∑)∈30~50为中营养;TLI(∑)∈50~60为轻度富营养;TLI(∑)∈60~70为中度富营养;TLI(∑)∈70~100为高度富营养.
1.5 数据分析
利用One way ANOVA 对不同营养类型湖泊环境因子变量均值进行分析(P<0.05),在进行方差分析前进行齐性检验,将不符合正态分布的水温、SD、TN、TP、Chl.a通过Rank cases的转换满足参数的统计要求. 采用相关分析研究环境因子之间的相关性,并建立环境因子相关性矩阵. 运用RDA(CANOCO 4.5)分析水生植物与环境因子之间的关系,采用蒙特卡洛检测方法检测环境因子对植物影响的显著性.
2 结果
2.1 不同营养类型湖泊水环境现状、显著性差异及与环境因子之间的相互关系
综合营养状态指数法计算表明10个淡水湖泊营养水平整体呈上升趋势(图1). 城东湖、固城湖和骆马湖属于中营养湖泊,富营养指数分别为49.33、44.10和49.21,其中骆马湖和城东湖的富营养水平相比于以前有所加剧[20-21],更接近轻度富营养水平,而固城湖富营养指数相较以前的调查数据[21]有所下降,湖泊水质得到了改善. 阳澄湖、高邮湖、香涧湖和花园湖是轻度富营养湖泊,富营养指数分别为52.14、58.97、59.91和53.74,除了阳澄湖的富营养指数降低外[21],其余3个淡水湖泊的富营养指数较以前都有所上升[17-18,20-21]. 洪泽湖、邵伯湖和长荡湖是中度富营养湖泊,富营养指数分别为60.98、61.65和66.51,所有湖泊的营养水平与以前相比都有不同程度的上升[21],其中洪泽湖的富营养指数已经达到中度富营养湖泊标准.
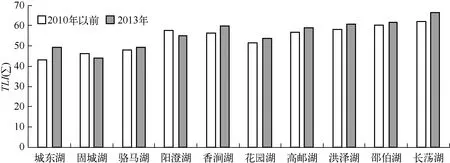
图1 不同时期湖泊富营养指数变化Fig.1 Changes of eutrophication index of lakes in different years
SPSS统计分析表明,不同营养类型湖泊的环境因子之间存在较强异质性(表2). 本次调查的湖泊水体均呈碱性,pH平均值位于8.40~9.07之间,中度富营养湖泊pH值最高,且与中营养和轻度富营养湖泊间存在显著差异(P<0.05).3种营养类型湖泊的DO均较高,其平均值范围为11.13~14.07 mg/L,且不同营养水平湖泊间差异不显著(P>0.05). 各湖泊营养盐浓度总体上表现出由中营养湖泊到中度富营养湖泊递增的趋势,TN、TP浓度平均范围分别为0.97~2.57 mg/L和0.03~0.13 mg/L,多重比较分析表明轻度富营养湖泊TN浓度与中营养(P<0.05)和中度富营养湖泊(P<0.001)存在显著性差异,中营养湖泊的TP浓度与轻度富营养(P<0.05)和中度富营养湖泊(P<0.001)存在显著性差异. 不同营养类型湖泊Chl.a含量和SD呈现相反的变化规律,中营养到中度富营养Chl.a含量显著增加,SD显著降低,中度富营养湖泊Chl.a含量是中营养湖泊的4.28倍,而SD只有中营养湖泊的1/4.
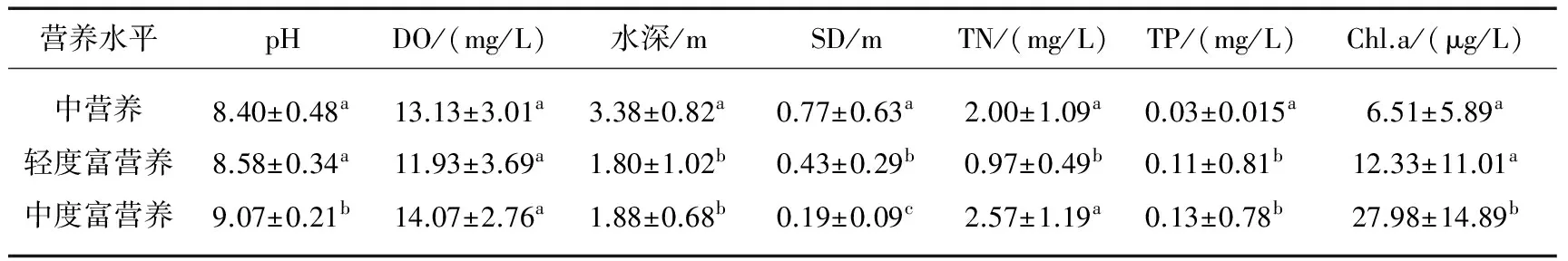
表2 不同营养水平湖泊环境因子指标*
*纵向不同字母代表该值在不同类型湖泊间存在显著差异(P<0.05).
水环境因子相关分析研究结果表明,各环境因子之间存在显著相关(表3). pH值与DO呈极显著正相关(P<0.01),与水温呈极显著负相关(P<0.01);SD与水深呈显著正相关(P<0.05),与TP和Chl.a含量呈极显著负相关(P<0.01);Chl.a含量与TN、TP浓度之间呈现极显著正相关(P<0.05).
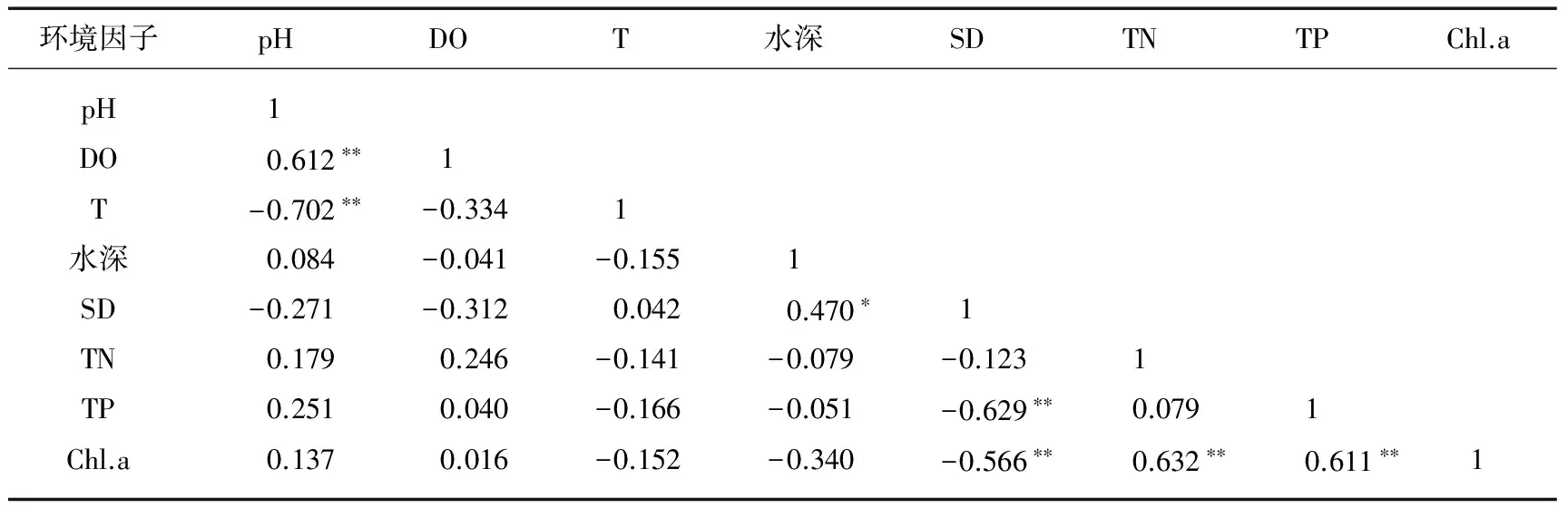
表3 水环境因子的相关性矩阵
*表示显著差异,P<0.05; **表示极显著差异,P<0.01.
2.2 不同营养类型湖泊水生植物种类组成及数量
本次调查表明,该区域不同营养类型湖泊主要水生植物有6科7属11种,生活型以沉水植物为主,包括菱(Trapabispinosa)、荇菜(Nymphoidespeltatum)、马来眼子菜(Potamogetonmalaianus)、金鱼藻(Ceratophyllumdemersum)、轮叶黑藻(Hydrillaverticillata)、穗花狐尾藻(Myriophyllumspicatum)、苦草(Vallisnerianatans)、菹草(P.crispus)、篦齿眼子菜(P.pectinatus)、微齿眼子菜(P.maackianus)和水鳖(Hydrocharisdubia).
调查发现长江下游湖泊中的主要优势种为金鱼藻、菱、荇菜、轮叶黑藻、穗花狐尾藻和马来眼子菜. 研究表明不同营养类型湖泊水生植物的优势种不同,浮叶植物菱和荇菜在各营养水平湖泊中均能成为优势或者亚优势种. 中营养湖泊的优势种有浮叶植物菱、荇菜以及沉水植物马来眼子菜,其中荇菜的重要值最高,其次是菱和马来眼子菜. 轻度富营养湖泊的优势种主要为金鱼藻、轮叶黑藻、穗花狐尾藻等,其中以菱的重要值最高,其次是金鱼藻、穗花狐尾藻、轮叶黑藻. 中度富营养湖泊的优势种为菱、荇菜、穗花狐尾藻和马来眼子菜,重要值最高的是荇菜,其次是马来眼子菜、穗花狐尾藻和菱(表4). 不同营养类型湖泊间优势种的单位面积平均生物量差异巨大,范围介于6.24~2465.78 g之间,而且不同营养水平湖泊间同一物种生物量差异显著. 一般而言,轻度富营养湖泊中水生植物单位面积平均生物量最高,其次为中度富营养湖泊,中营养水平湖泊单位面积水生植物的平均生物量最低. 从不同物种生物量看,浮叶植物菱的单位面积平均生物量最大,其次为荇菜和马来眼子菜,轮叶黑藻最少.
2.3 水生植物群落物种与环境因子的关系
水生植物群落物种与环境因子的RDA分析结果显示,4个排序轴对物种和环境因子关系的累积方差为98.8%,其中第1、2轴共解释物种与环境因子之间累计方差为83.3%,说明排序结果能够很好地解释群落组成与环境因子之间的关系(表5).
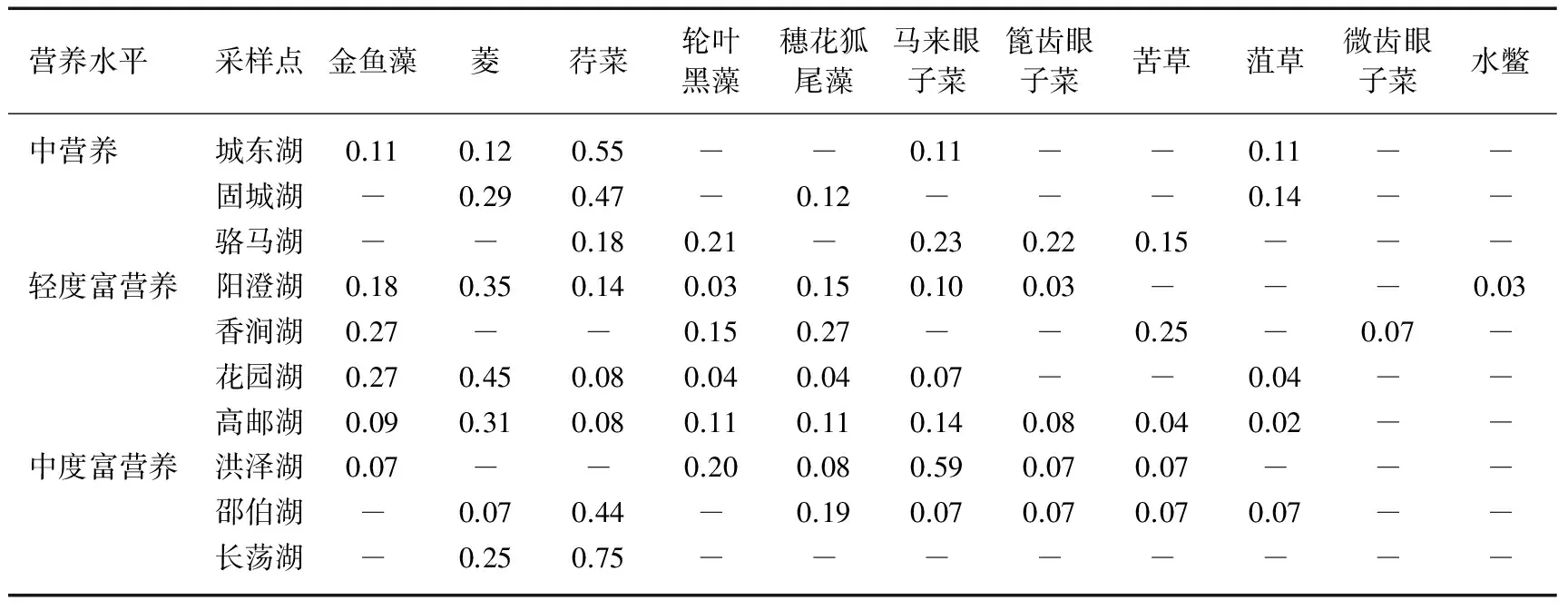
表4 调查湖泊中水生植物种类组成及重要值
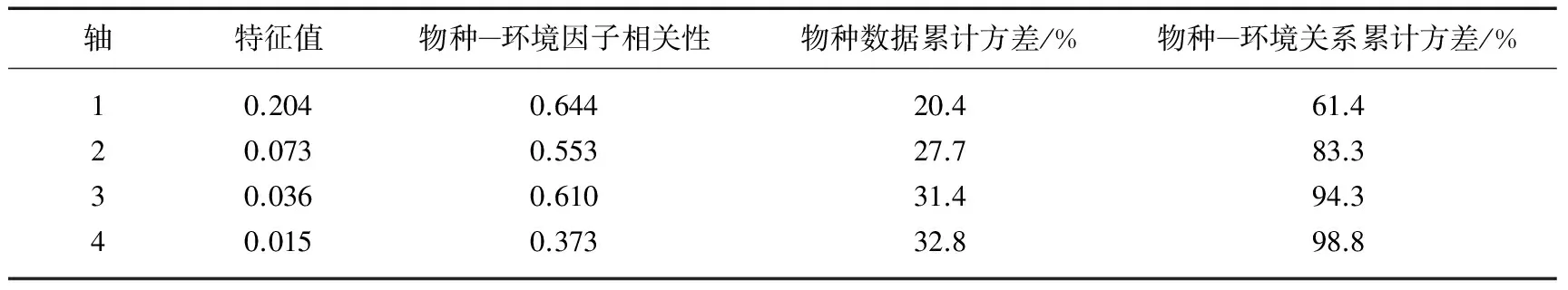
表5 物种与水环境因子的相关系数
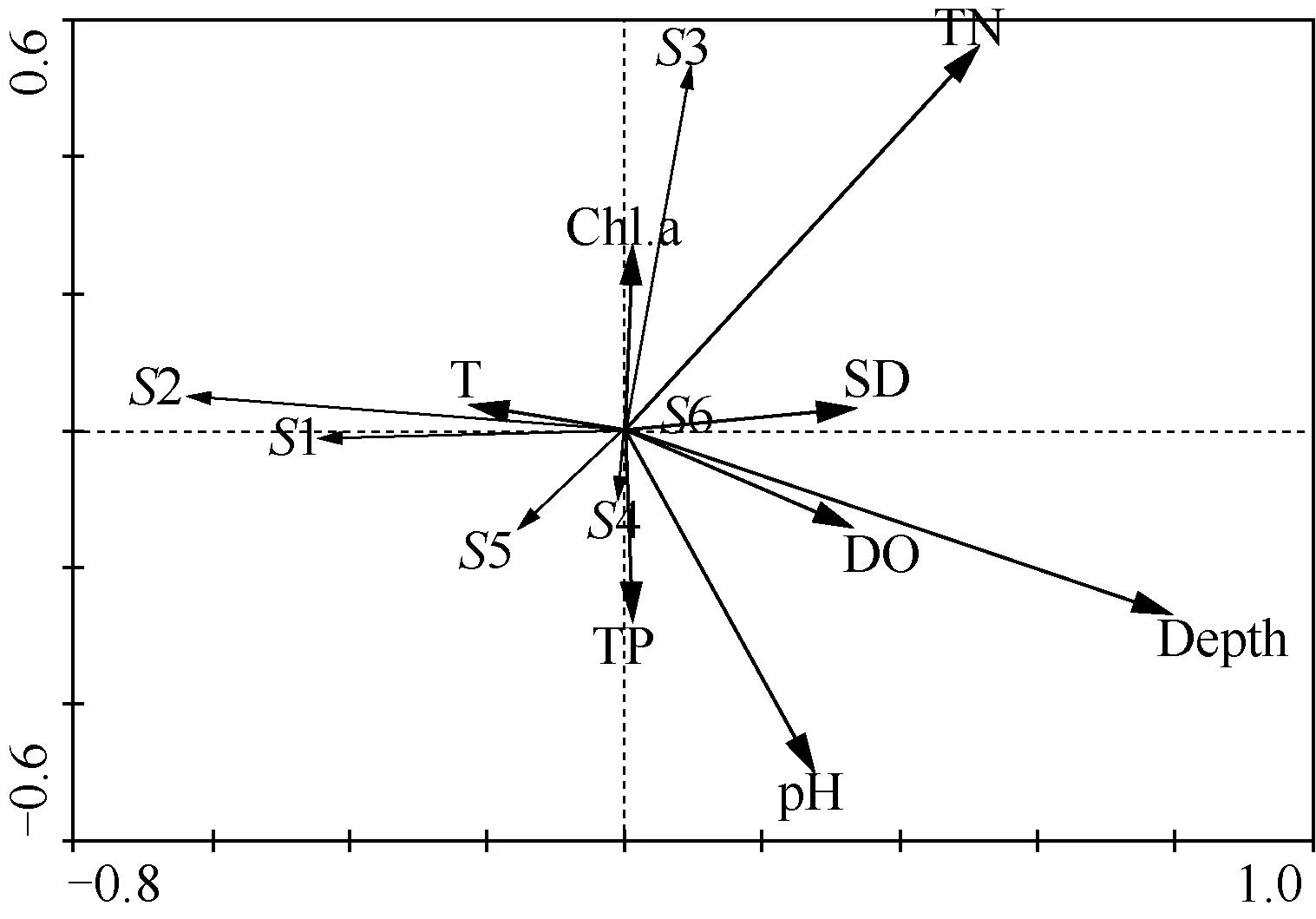
图2 基于RDA分析的物种与环境变量双序图(S1:金鱼藻,S2:菱,S3:荇菜,S4:轮叶黑藻,S5:穗花狐尾藻,S6:马来眼子菜)Fig.2 RDA ordination diagram of species and environmental variables
RDA分析表明:金鱼藻和菱这两种水生植物与水温呈显著正相关,与水深、pH值、TN、DO、SD呈显著负相关;荇菜与Chl.a和TN呈显著正相关,与pH值和TP呈显著负相关;穗花狐尾藻与水温、pH值呈显著正相关,与TN呈极显著负相关;轮叶黑藻与pH值呈极显著正相关,与Chl.a呈极显著负相关;沉水植物马来眼子菜与SD、DO、水深、TN呈显著正相关,与水温呈显著负相关. 从总体影响程度来看,TN、水深、pH值是影响长江下游水生植物生长分布的主要因子(图2).
3 讨论
本次调查发现长江下游湖泊主要水生植物共计11种,其中优势种包括金鱼藻、荇菜、菱等,且主要分布在轻度富营养湖泊中,相对而言中营养湖泊和中度富营养湖泊的水生植物多样性和生物量较低. 调查发现6种优势水生植物在不同营养水平湖泊中生物量分布不均,特别是浮叶植物菱和沉水植物马来眼子菜分别在轻度富营养湖泊和中度富营养湖泊中的生物量显著高于中营养湖泊. 导致这一现象的原因可能有以下两点:(1) 自然因素:水体生境条件对水生植物生长和分布具有重要影响,已有研究表明水温、水深、SD、营养盐、pH值等环境因子都对水生植物的生长产生显著影响[15]. (2) 人为因素:城东湖、固城湖和骆马湖等都是长江下游重要的水产养殖湖泊,渔业发达,渔船航行威胁到沉水植物的生长[20-21].
水深对水生植物的生长和分布具有重要影响[24-26],RDA分析中水深解释了物种与环境关系的18.9%,是影响水生植物的首要因子,蒙特卡洛检测呈极显著相关(P<0.01). 相关研究表明, 水深/SD是影响大型水生植物生长的关键因素[27],当水深不超过2 m时,植物的盖度和生物量明显受到水深的限制[28],而水深/SD为5.26时,大部分水生植物的生长会受到威胁[9]. 本次调查的湖泊平均深度为2.35 m,水深/SD平均值为5.47,限制了水生植物在不同营养水平湖泊中的分布,中度富营养湖泊(水深/SD为9.89)水生植物的生物量要明显低于轻度富营养湖泊(水深/SD为4.19).此外,这次调查发现菱的分布与水深呈显著负相关,在3种不同营养水平湖泊中,菱在各采样点内平均生物量和平均水深分别为:中营养湖泊(102.78 g、3.38 m)<中度富营养湖泊(647.91 g、1.88 m)<轻度富营养湖泊(2465.78 g、1.80 m),随着水体深度的降低,菱的生物量不断上升.
调查发现TN是影响长江下游湖泊水生植物分布的又一重要因子,蒙特卡洛检测呈显著相关(P<0.05). 所调查的长江下游10个淡水湖泊的TN浓度平均范围为0.97~2.57 mg/L,中营养湖泊的TN浓度平均值(2.00 mg/L)要高于轻度富营养湖泊(0.97 mg/L),这可能是因为部分湖泊周围含氮废水的大量排放[21],导致水体的TN浓度高于一般中营养湖泊. 已有研究表明,在一定范围内,较高的TN浓度有利于沉水植物马来眼子菜的生长[26],该研究证实了这一研究成果,RDA分析显示在所调查的10个不同营养程度湖泊中马来眼子菜与TN浓度呈显著正相关(图2),马来眼子菜的平均生物量在不同营养类型湖泊间的大小表现为中度富营养(1601.96 g)>中营养(465.17 g)>轻度富营养(254.95 g). 然而,该研究表明TP并不是影响长江下游浅水湖泊水生植物生长分布的主要因子,这可能因为沉水植物不仅可以从水体中获取磷,湖水底泥也是根生水生植物的重要营养源[27].
pH值显著影响水生植物的生长和分布[18-19],在RDA分析中解释了物种和环境关系的9.8%. 以往研究发现,当pH值为7~9时,富营养状态相较于中营养状态更能促进轮叶黑藻的生长[30],本次调查结果与其一致,即在pH值为7.92~8.92的湖水中,随着营养水平和pH值的增大,轮叶黑藻分布的盖度和生物量递增.
RDA分析结果表明,Chl.a、TP、DO和水温对水生植物分布的影响较小. 但已有研究和本次调查均表明Chl.a与TN、TP之间存在极显著关系,可以直接影响水体SD的大小[31-32]. 这说明水环境因子之间是相互影响、相互制约的,一个因子可以通过影响其他环境因子进而影响水生植物的生长和分布. 因此,在研究水生植物生长分布的影响因子时,应当考虑到水环境因子之间的综合作用及内在联系,全面分析水生植物与环境因子之间的关系.
[1] 张 萌,倪乐意,曹 特等. 太湖上游水环境对植物分布格局的影响机制. 环境科学与技术, 2009, 33(3):171-177.
[2] 雷泽湘,徐德兰,顾继光等. 太湖大型水生植物分布特征及其对湖泊营养盐的影响. 农业环境科学学报, 2008, 27(2): 698-704.
[3] 彭映辉,简永兴,李仁东等. 白莲湖与西凉湖水生植物多样性的比较研究. 经济林研究, 2003, 21(1):18-20.
[4] 武士蓉,徐梦佳,赵彦伟等. 白洋淀湿地水质与水生物相关性研究. 环境科学学报, 2013, 33(11):3160-3165.
[5] 施 娴,陈开宁,黄 蔚. 不同温度培养下莕菜(Nymphoidespeltata)生长与光合作用特征. 湖泊科学, 2013, 25(4): 545-550.
[6] 马 凯,蔡庆华,谢志才等. 沉水植物分布格局对湖泊水环境N、P因子影响. 水生生物学报, 2003, 27(3):232-236.
[7] 苏胜齐. 环境对菹草生长和繁殖的影响及菹草对富营养化水体净化能力的研究[学位论文]. 重庆: 西南农业大学, 2001: 35-50.
[8] 任久长,周 红,孙亦彤. 滇池光照强度的垂直分布与沉水植物的光补偿深度. 北京大学学报: 自然科学版, 1997, 33(2): 211-215.
[9] 陈中义,雷泽湘,周 进等. 梁子湖6种沉水植物种群数量和生物量周年动态. 水生生物学报, 2000, 24(6):582-588.
[10] 金相灿,颜昌宙,许秋瑾. 太湖北岸湖滨带观测场水生植物群落特征及其影响因素分析. 湖泊科学, 2007, 19(2): 151-157.
[11] Vermaat JE, Vethagen FCA, Lindenburg D. Contrasting responses in two populations ofZosteranoltitHomem to experimental photoperiod manipulation at two salinities.AquaticBotany, 2000, 67: 179-189.
[12] 任艳芹,陈开宁. 巢湖沉水植物现状(2010年)及其与环境因子的关系. 湖泊科学, 2011, 23(3): 409-416.
[13] 张兰芳. 水质条件对沉水植物(伊乐藻、菹草)生长的影响[学位论文]. 南京: 河海大学, 2006: 41-45.
[14] 任 南,严国安,马剑敏等. 环境因子对东湖几种沉水植物生理的影响研究. 武汉大学学报: 自然科学版, 1996, 42(2): 213-218.
[15] 秦伯强. 长江中下游浅水湖泊富营养化发生机制与控制途径初探. 湖泊科学, 2002, 14(3): 193-202.
[16] Fang JY, Wang ZH, Zhang SQetal. Biodiversity changes in the lakes of Central Yangtze.FrontierinEcologyandtheEnvironment, 2006, 4(7): 369-377.
[17] 吴爱平,吴世凯,倪乐意. 长江中游浅水湖泊水生植物氮磷含量与水柱营养的关系. 水生生物学报, 2005, 29(4): 406-412.
[18] 何 俊,谷孝鸿,刘国锋. 东太湖水生植物及其与环境的相互作用. 湖泊科学, 2008, 20(6): 790-795.
[19] 任 南,严国安,马剑敏. 环境因子对东湖几种沉水植物生理的影响研究. 武汉大学学报, 1996,42(2):213-218.
[20] 刘清学,霍守亮,昝逢宇等. 安徽省湖泊富营养化现状调查与评价. 安徽农业科学, 2011, 39(8):4626-4629.
[21] 范成新,羊向东,史龙新等. 江苏省湖泊富营养化特征、成因及解决途径. 长江流域资源与环境, 2005, 14(2):218-223.
[22] 王明翠,刘雪芹, 张建辉. 湖泊富营养化评价方法及分级标准. 中国环境监测, 2002, 18(5): 47-49.
[23] 金相灿. 中国湖泊环境. 北京: 海洋出版社, 1995: 44-47.
[24] Paillisson JM, Marion L. Can small water level fluctuation affect the biomass ofNymphaeaablein large lakes?AquaticBotany, 2006, 84: 259-266.
[25] Scheffer M. Ecology of shallow lakes. London: Chapmana and Hall, 1998: 40-51.
[26] 苏睿丽,李 伟. 沉水植物光合作用的特点与研究进展. 植物学通报, 2005, 22(增刊):128-138.
[27] Wang HZ, Wang HJ, Liang XM. Empirical modelling of submersed macrophytes in Yangtze lakes.EcologicalModelling, 2005, 188: 483-491.
[28] Gafhy S, Gasith A. Spatially and temporally sporadic appearance of macrophytes in the littoral zone of Lake Kinneret, Israel: taking advantage of a window of opportunity.AquaticBotany, 1999, 62: 249-267.
[29] 叶 春,邹国燕,付子轼等. 总氮浓度对3种沉水植物生长的影响. 环境科学学报, 2007, 27(5): 739-745.
[30] 张 浏,陈 灿,高 倩等. 两种营养状态下pH对轮叶黑藻(Hydrillaverticillata)生长和抗氧化酶活性的影响. 生态环境, 2007, 16(3): 748-752.
[31] 董云仙,洪雪花,谭志卫等. 高原深水湖泊程海氮磷形态分布特征及其与叶绿素 a 的相关性. 生态环境科学, 2012, 21(2): 333-337.
[32] 王 华,逢 勇,刘申宝等. 沉水植物生长影响因子研究进展. 生态学报, 2008, 28(8): 3958-3968.
Status of aquatic plants and its relationship with water environment factors in the lakes along the lower reaches of the Yangtze River
KONG Xianghong1, XIAO Lanlan2, SU Haojie3, WU Yao3, ZHANG Xiaolin3& LI Zhongqiang1
(1:SchoolofResourceandEnvironmentalScience,HubeiUniversity,Wuhan430062,P.R.China)
(2:ZhengzhouInfantNormalSchool,Zhengzhou450002,P.R.China)
(3:DonghuExperimentalStationofLakeEcosystems,InstituteofHydrobiology,ChineseAcademyofSciences,Wuhan430072,P.R.China)
The relationship between aquatic plants and environmental factors provides a significant scientific basis for lake restoration. In this study, we investigated community composition of aquatic plants and the relationship between macrophytes and environmental factors, based on the survey about 10 lakes with different trophic levels in the lower reaches of the Yangtze River, from June to July in 2013. Results showed that there were 11 macrophytes species belonging to 6 families, 7 genera. Redundancy analysis indicated that total nitrogen, pH and depth are the key factors for the distribution of aquatic plants in lakes with different trophic levels in the lower reaches of the Yangtze River.
Aquatic plants; environmental factors; trophic level; main factor; redundancy analysis;lower reaches of the Yangtze River
*国家自然科学基金项目(31270261)、国家科技基础性工作专项(2013FY112300)和国家水体污染控制与治理科技重大专项(2012ZX07101-002)联合资助.2014-07-28收稿;2014-12-18收修改稿. 孔祥虹(1990~),女,硕士研究生;E-mail: 1015447407@qq.com.
J.LakeSci.(湖泊科学), 2015, 27(3): 385-391
http: //www.jlakes.org.E-mail: jlakes@niglas.ac.cn
©2015 byJournalofLakeSciences
**通信作者;E-mail:zhangxl@ihb.ac.cn; lizhq@hubu.edu.cn.
